
Combined Metabolome and Transcriptome Profiling Reveal Optimal Harvest Strategy Model Based on Different Production Purposes in Olive



Abstract
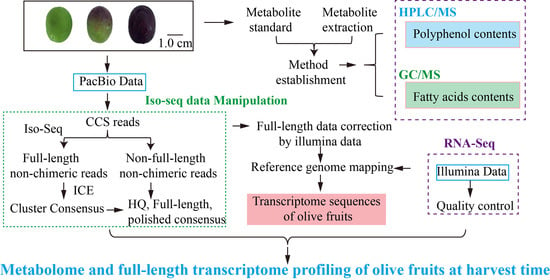
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. GC/MS- and LC/MS-Based Targeted and Untargeted Metabolomic Analysis
2.3. RNA Extraction and Iso-Seq and RNA-Seq Sequencing
3. Results
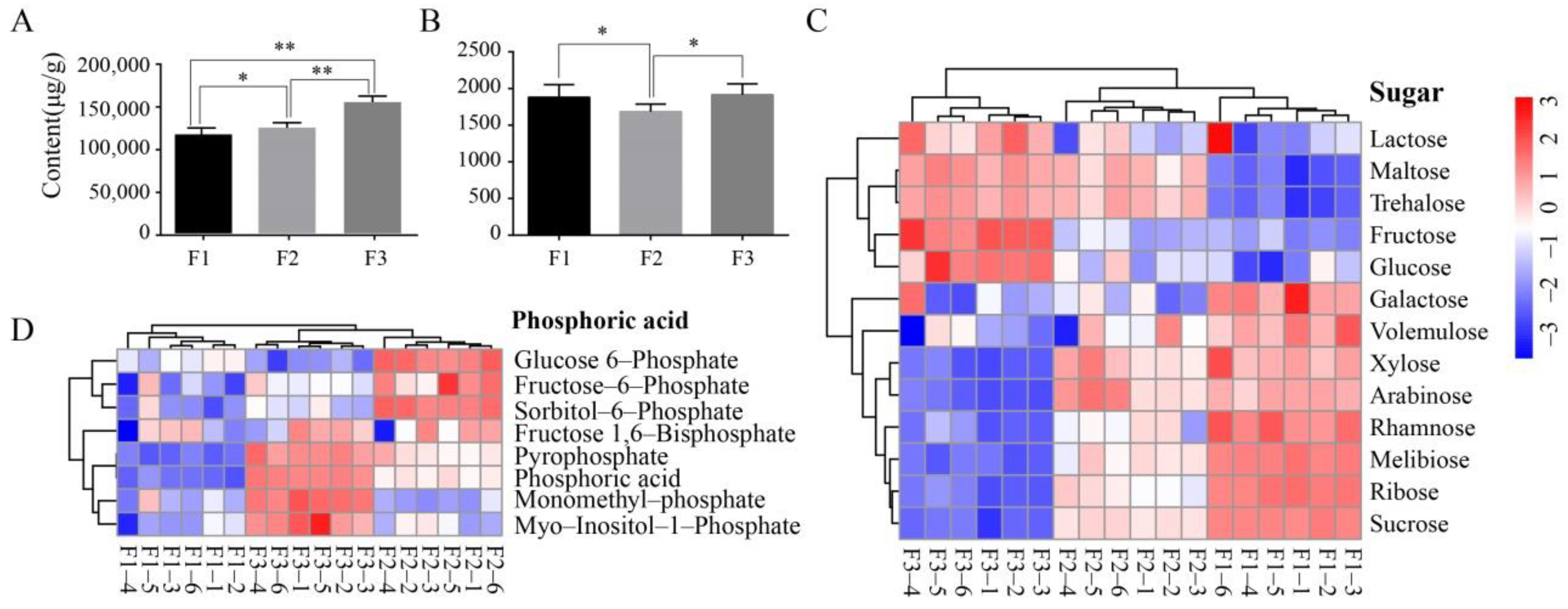
3.1. Comprehensive Metabolomic Analysis of Olive Fruits at Different Maturity Stages
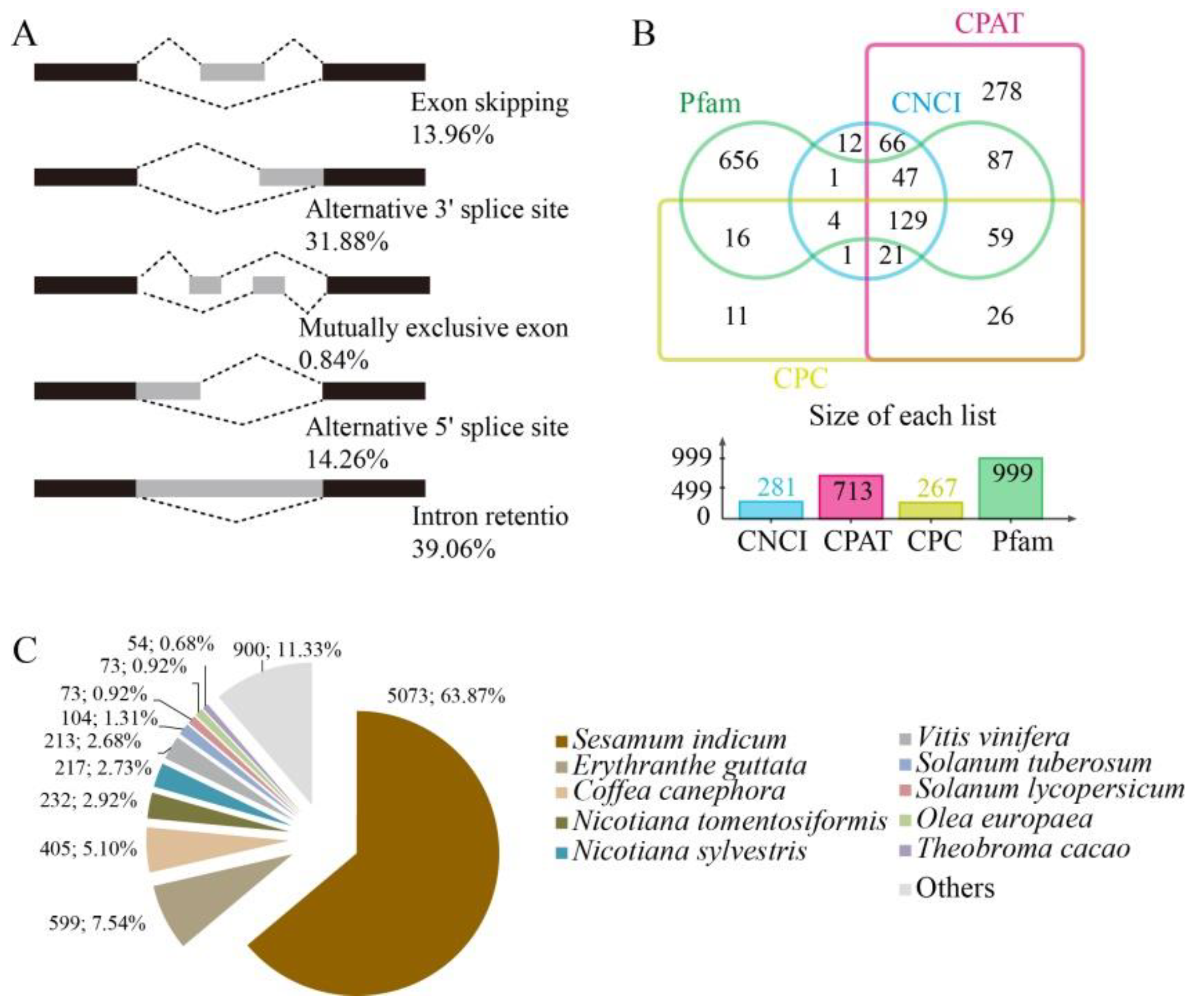
3.2. Transcriptomic Profiling with PacBio Full-Length and Illumina RNA-Seq
3.3. Combined Transcriptomic and Metabolomics Analysis of Hydroxytyrosol and Oleuropein Biosynthesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Navarro, M.; Morales, F.J. Effect of hydroxytyrosol and olive leaf extract on 1,2-dicarbonyl compounds, hydroxymethylfurfural and advanced glycation endproducts in a biscuit model. Food Chem. 2017, 217, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Luchetti, F. Importance and future of olive oil in the world market-an introduction to olive oil. Eur. J. Lipid Sci. Technol. 2010, 104, 559–563. [Google Scholar] [CrossRef]
- Mougiou, N.; Trikka, F.; Trantas, E.; Ververidis, F.; Makris, A.; Argiriou, A.; Vlachonasios, K.E. Expression of hydroxytyrosol and oleuropein biosynthetic genes are correlated with metabolite accumulation during fruit development in olive, Olea europaea, cv. Koroneiki. Plant Physiol. Biochem. 2018, 128, 41–49. [Google Scholar] [CrossRef]
- Ouni, Y.; Flamini, G.; Zarrouk, M. The Chemical properties and volatile compounds of virgin olive oil from oueslati variety: Influence of maturity stages in olives. J. Am. Oil Chem. Soc. 2016, 93, 1–9. [Google Scholar] [CrossRef]
- Elena, G.; Vincent, B.; Pierna, J.A.F.; José, A.G. Determination of the olive maturity index of intact fruits using image analysis. Int. J. Food Sci. Technol. 2015, 52, 1462. [Google Scholar]
- Vriet, C.; Russinova, E.; Reuzeau, C. From squalene to brassinolide: The steroid metabolic and signaling pathways across the plant kingdom. Mol. Plant 2013, 6, 1738–1757. [Google Scholar] [CrossRef]
- Dag, A.; Kerem, Z.; Yogev, N.; Zipori, I.; Lavee, S.; Ben-David, E. Influence of time of harvest and maturity index on olive oil yield and quality. Sci. Hortic. 2011, 127, 358–366. [Google Scholar] [CrossRef]
- Lazzez, A.; Perri, E.; Caravita, M.A.; Khlif, M.; Cossentini, M. Influence of olive maturity stage and geographical origin on some minor components in virgin olive oil of the chemlali variety. J. Agric. Food Chem. 2008, 56, 982–988. [Google Scholar] [CrossRef]
- Bouaziz, M.; Chamkha, M.S. Comparative study on phenolic content and antioxidant activity during maturation of the olive cultivar Chemlali from Tunisia. J. Agric. Food Chem. 2004, 52, 5476–5481. [Google Scholar] [CrossRef]
- Cruz, F.; Julca, I.; Gomez-Garrido, J.; Loska, D.; Marcet-Houben, M.; Cano, E.; Galan, B.; Frias, L.; Ribeca, P.; Derdak, S.; et al. Genome sequence of the olive tree, Olea europaea. Gigascience 2016, 5, 29. [Google Scholar] [CrossRef]
- Giampetruzzi, A.; Morelli, M.; Saponari, M.; Loconsole, G.; Chiumenti, M.; Boscia, D.; Savino, V.N.; Martelli, G.P.; Saldarelli, P. Transcriptome profiling of two olive cultivars in response to infection by the CoDiRO strain of Xylella fastidiosa subsp. pauca. BMC Genom. 2016, 17, 1–18. [Google Scholar] [CrossRef]
- Muñoz-Mérida, A.; González-Plaza, J.J.; Cañada, A.; Blanco, A.M.; García-López, M.C.; Rodríguez, J.M.; Pedrola, L.; Sicardo, M.D.; Hernández, M.L.; Rosa, R.D.; et al. De novo assembly and functional annotation of the olive (Olea europaea) Transcriptome. DNA Res. 2013, 20, 93–108. [Google Scholar] [CrossRef]
- Guodong, R.; Xiaoxia, L.; Weiwei, Z.; Wenjun, W.; Jianguo, Z. Metabolomics reveals variation and correlation among different tissues of olive (Olea europaea L.). Biol. Open 2017, 6, 1317. [Google Scholar] [CrossRef]
- Rao, G.; Zhang, J.; Liu, X.; Luo, Y. Identification of putative genes for polyphenol biosynthesis in olive fruits and leaves using full-length transcriptome sequencing. Food Chem. 2019, 300, 125246. [Google Scholar]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, 36. [Google Scholar] [CrossRef]
- Omar, S.H. Oleuropein in olive and its pharmacological effects. Sci. Pharm. 2010, 78, 133. [Google Scholar] [CrossRef]
- Ullrich, J.; Calvin, M. Mechanism of methyl phosphate formation by “killing” of spinach chloroplast fragments with methanol in the presence of phosphate. Biochim. Biophys. Acta 1962, 57, 190–191. [Google Scholar] [CrossRef]
- Foissac, S.; Sammeth, M. Astalavista: Dynamic and flexible analysis of alternative splicing events in custom gene datasets. Nucleic Acids Res. 2007, 35, W297–W299. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Zhang, Y.; Ye, Z.Q.; Liu, X.Q.; Zhao, S.Q.; Wei, L.; Gao, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 2007, 35, W345. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Luo, H.; Bu, D.; Zhao, G.; Yu, K.; Zhang, C.; Liu, Y.; Chen, R.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Park, H.J.; Dasari, S.; Wang, S.; Kocher, J.P.; Li, W. CPAT: Coding-potential assessment tool using an alignment-free logistic regression model. Nucleic Acids Res. 2013, 41, e74. [Google Scholar] [CrossRef]
- Gordon, M.H.; Paiva-Martins, F.; Almeida, M. Antioxidant activity of hydroxytyrosol acetate compared with that of other olive oil polyphenols. J. Agric. Food Chem. 2001, 49, 2480. [Google Scholar] [CrossRef]
- Miettinen, K.; Dong, L.; Navrot, N.; Schneider, T.; Burlat, V.; Pollier, J.; Woittiez, L.; Krol, S.V.D.; Lugan, R.; Ilc, T. Corrigendum: The seco-iridoid pathway from Catharanthus roseus. Nat. Commun. 2014, 5, 3606–3616. [Google Scholar] [CrossRef]
- Damtoft, S.; Franzyk, H.; Jensen, S.R. Biosynthesis of iridoids in Syringa and Fraxinus: Secoiridoid precursors. Phytochemistry 1995, 40, 773–784. [Google Scholar] [CrossRef]
- Francisca, G.R.; María Paz, R.; María, C.; María José, M.; María Isabel, M.M. Metabolites involved in oleuropein accumulation and degradation in fruits of Olea europaea L.: Hojiblanca and Arbequina varieties. J. Agric. Food Chem. 2010, 58, 12924. [Google Scholar]
- Bendini, A.; Cerretani, L.; Carrasco-Pancorbo, A.; Gómez-Caravaca, A.; Segura-Carretero, A.; Fernández-Gutiérrez, A.; Lercker, G. Phenolic molecules in virgin olive oils: A survey of their sensory properties, health effects, antioxidant activity and analytical methods. An overview of the last decade alessandra. Molecules 2007, 12, 1679–1719. [Google Scholar] [CrossRef]
- Andreadou, I.; Iliodromitis, E.K.; Mikros, E.; Constantinou, M.; Agalias, A.; Magiatis, P.; Skaltsounis, A.L.; Kamber, E.; Tsantili-Kakoulidou, A.; Kremastinos, D.T. The olive constituent oleuropein exhibits anti-ischemic, antioxidative, and hypolipidemic effects in anesthetized rabbits. J. Nutr. 2006, 136, 2213–2219. [Google Scholar] [CrossRef] [PubMed]
- Romero, C.; Medina, E.; Mateo, M.A.; Brenes, M. Quantification of bioactive compounds in Picual and Arbequina olive leaves and fruit. J. Sci. Food Agric. 2017, 97, 1725–1732. [Google Scholar] [CrossRef] [PubMed]
- Cecchi, L.; Migliorini, M.; Cherubini, C.; Giusti, M.; Zanoni, B.; Innocenti, M.; Mulinacci, N. Phenolic profiles, oil amount and sugar content during olive ripening of three typical Tuscan cultivars to detect the best harvesting time for oil production. Food Res. Int. 2013, 54, 1876–1884. [Google Scholar] [CrossRef]
- Trapani, S.; Migliorini, M.; Cherubini, C.; Cecchi, L.; Canuti, V.; Fia, G.; Zanoni, B. Direct quantitative indices for ripening of olive oil fruits to predict harvest time. Eur. J. Lipid Sci. Technol. 2016, 118, 1202–1212. [Google Scholar] [CrossRef]
- Spanova, M.; Daum, G. Squalene-biochemistry, molecular biology, process biotechnology, and applications. Eur. J. Lipid Sci. Technol. 2011, 113, 1299–1320. [Google Scholar] [CrossRef]
- Fernández-Cuesta, A.; León, L.; Velasco, L.; Rosa, R.D.L. Changes in squalene and sterols associated with olive maturation. Food Res. Int. 2013, 54, 1885–1889. [Google Scholar] [CrossRef]
- Huang, Z.R.; Lin, Y.K.; Fang, J.Y. Biological and pharmacological activities of squalene and related compounds: Potential uses in cosmetic dermatology. Molecules 2009, 14, 540–554. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | F1 | F2 | F3 |
|---|---|---|---|
| C16:0 | 19,675.13 ± 1350.06 | 22,751.13 ± 801.68 | 28,291.71 ± 1905.85 |
| C16:1 | 663.19 ± 67.49 | 1064.50 ± 67.88 | 1722.41 ± 97.58 |
| C18:0 | 2587.96 ± 354.19 | 2661.12 ± 140.25 | 3453.95 ± 163.80 |
| C18:1 | 80,340.02 ± 9798.77 | 84,364.30 ± 4234.48 | 102,126.63 ± 4138.54 |
| C18:2 | 6612.13 ± 917.52 | 11,506.93 ± 487.85 | 16,425.71 ± 755.58 |
| C18:3 | 755.37 ± 99.69 | 676.34 ± 55.74 | 833.11 ± 73.71 |
| C20:0 | 354.63 ± 34.61 | 329.54 ± 20.78 | 391.51 ± 41.65 |
| C20:1 | 355.15 ± 82.82 | 357.00 ± 58.59 | 412.05 ± 53.37 |
| Vanillic acid | 13.17 ± 1.22 | n.d. | n.d. |
| Caffeic acid | n.d. | n.d. | 0.21 ± 0.03 |
| p-Coumaric acid | 0.60 ± 0.08 | n.d. | 1.45 ± 0.16 |
| Ferulic acid | 0.27 ± 0.03 | 0.28 ± 0.05 | 0.61 ± 0.08 |
| Maslinic acid | 1189.25 ± 146.23 | 913.34 ± 86.91 | 1075.54 ± 122.00 |
| Hydroxytyrosol | 21.28 ± 1.54 | 20.83 ± 1.78 | 27.25 ± 2.22 |
| Oleuropein | 15.18 ± 1.26 | 22.32 ± 1.63 | 6.76 ± 0.41 |
| Luteolin | 74.64 ± 6.24 | 31.93 ± 2.22 | 102.21 ± 7.73 |
| Apigenin | 6.90 ± 0.70 | 2.05 ± 0.32 | 4.76 ± 0.29 |
| Rutin | 285.29 ± 21.91 | 157.00 ± 17.40 | 248.58 ± 20.18 |
| Tyrosol | 11.12 ± 0.77 | 18.28 ± 1.15 | 23.63 ± 2.05 |
| Squalene | 262.49 ± 6.40 | 518.22 ± 5.12 | 425.14 ± 7.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rao, G.; Zhang, J.; Liu, X.; Li, X.; Wang, C. Combined Metabolome and Transcriptome Profiling Reveal Optimal Harvest Strategy Model Based on Different Production Purposes in Olive. Foods 2021, 10, 360. https://doi.org/10.3390/foods10020360
Rao G, Zhang J, Liu X, Li X, Wang C. Combined Metabolome and Transcriptome Profiling Reveal Optimal Harvest Strategy Model Based on Different Production Purposes in Olive. Foods. 2021; 10(2):360. https://doi.org/10.3390/foods10020360
Chicago/Turabian StyleRao, Guodong, Jianguo Zhang, Xiaoxia Liu, Xue Li, and Chenhe Wang. 2021. "Combined Metabolome and Transcriptome Profiling Reveal Optimal Harvest Strategy Model Based on Different Production Purposes in Olive" Foods 10, no. 2: 360. https://doi.org/10.3390/foods10020360
APA StyleRao, G., Zhang, J., Liu, X., Li, X., & Wang, C. (2021). Combined Metabolome and Transcriptome Profiling Reveal Optimal Harvest Strategy Model Based on Different Production Purposes in Olive. Foods, 10(2), 360. https://doi.org/10.3390/foods10020360