
Heterologous Expression of a Thermostable Chitinase from Myxococcus xanthus and Its Application for High Yield Production of Glucosamine from Shrimp Shell

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Colloidal Chitin Preparation, Gene Amplification and Plasmid Construction
2.3. Expression and Purification of Heterologous MxChi
2.4. Enzyme Activity Assay and Analytical Methods
2.5. Chitinase Biochemical Characterization
2.6. Enzymatic Synthesis of GlcNAc from Colloidal Chitin
2.7. Enzymatic Synthesis of GlcN from Colloidal Chitin and Shrimp Shell
2.8. Orthogonal Design for GlcN Production from Shrimp Shell
2.9. Phylogenetic Analysis Homology Modeling
2.10. Statistical Analysis
3. Results and Discussion
3.1. Cloning and Homology Analysis of the MxChi Gene
3.2. Recombinant Protein Expression and Purification
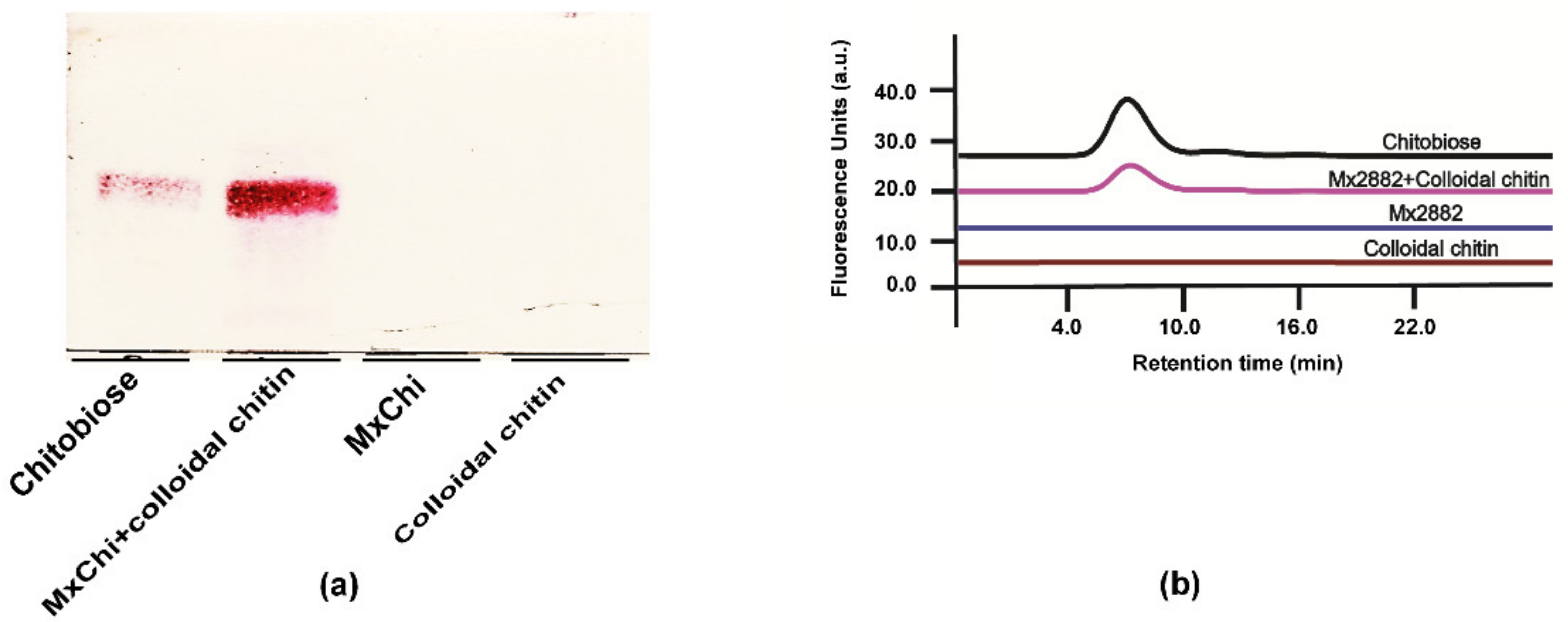
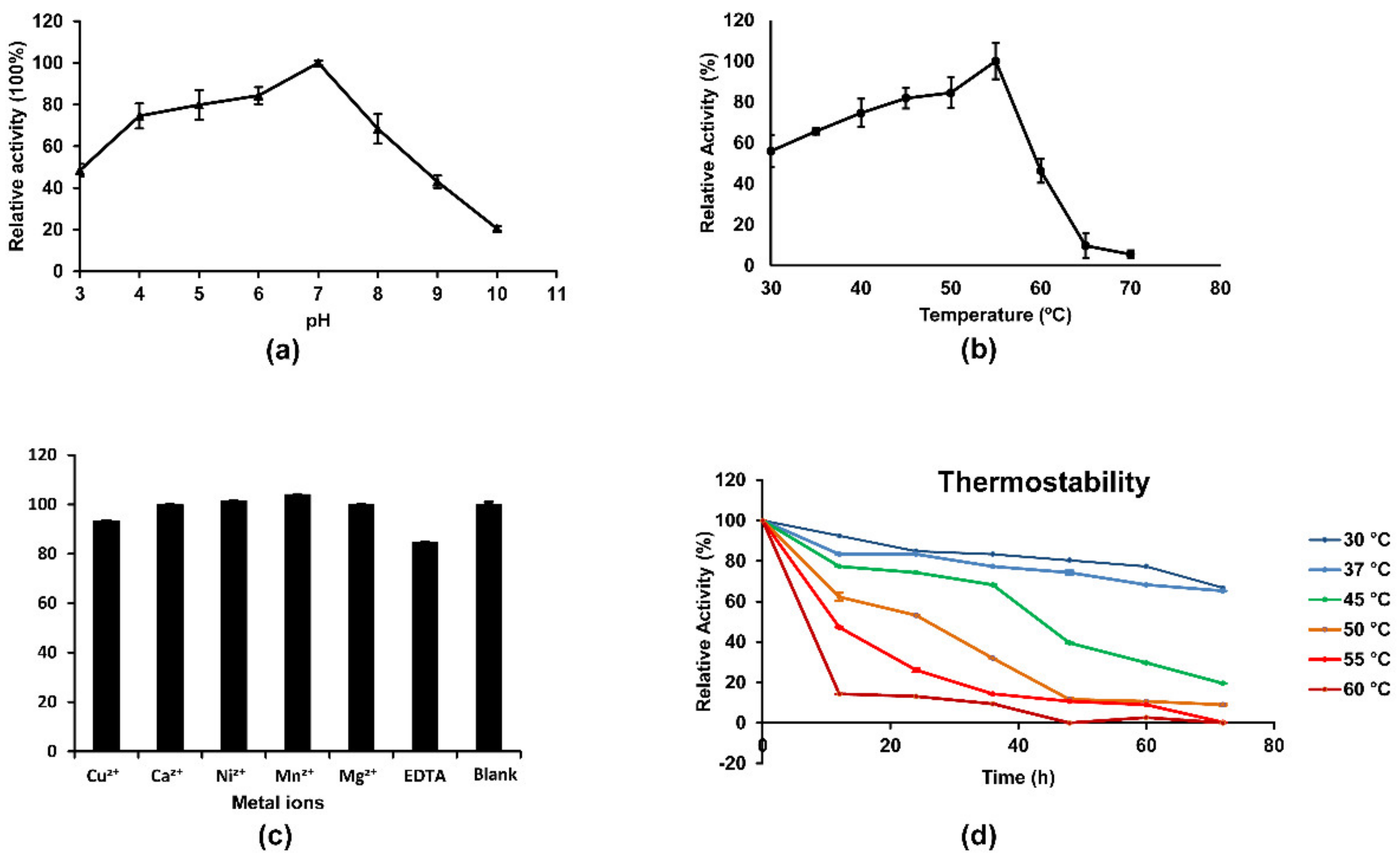
3.3. Enzymatic Properties of MxChi
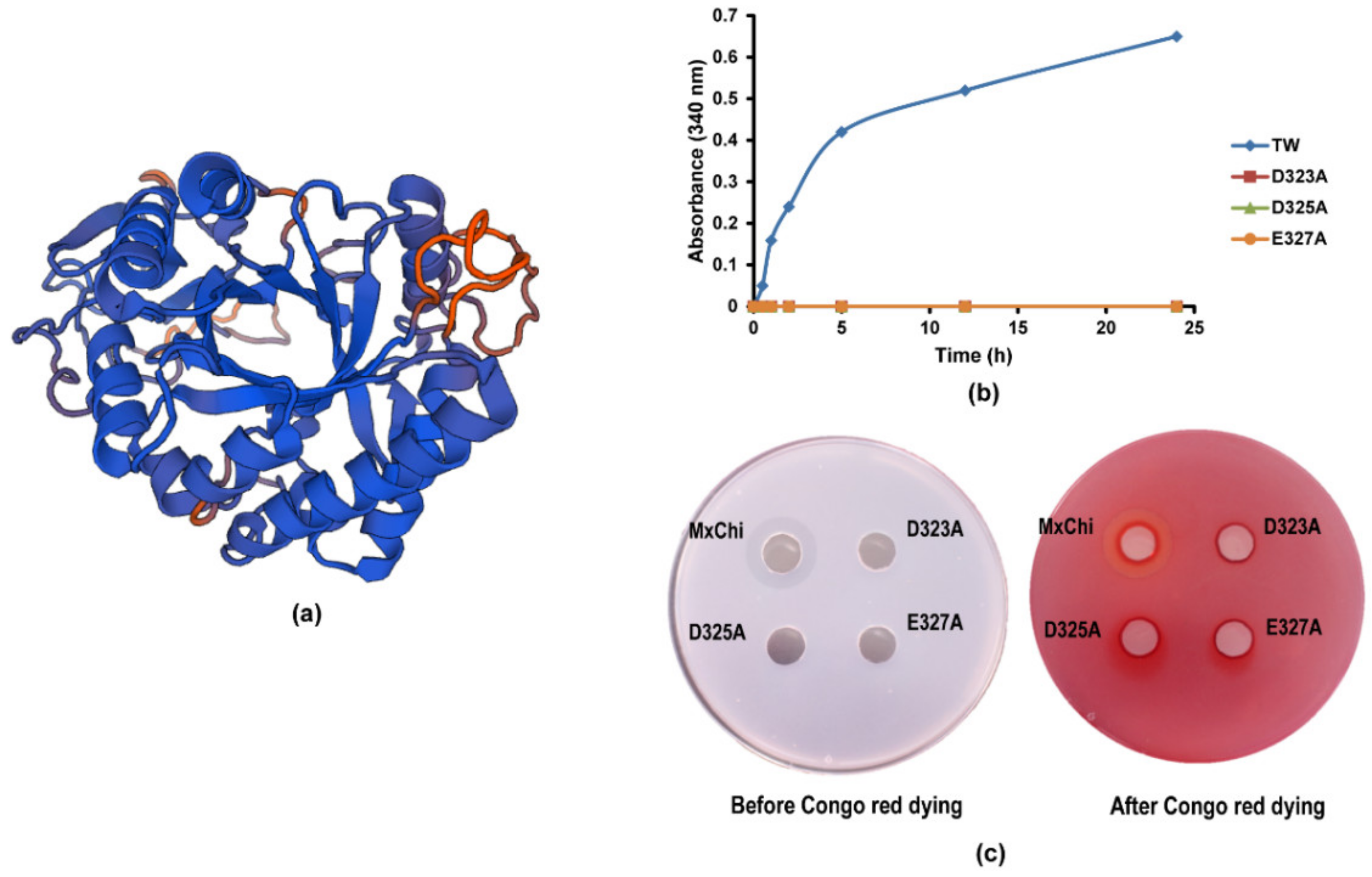
3.4. Mutational Analysis
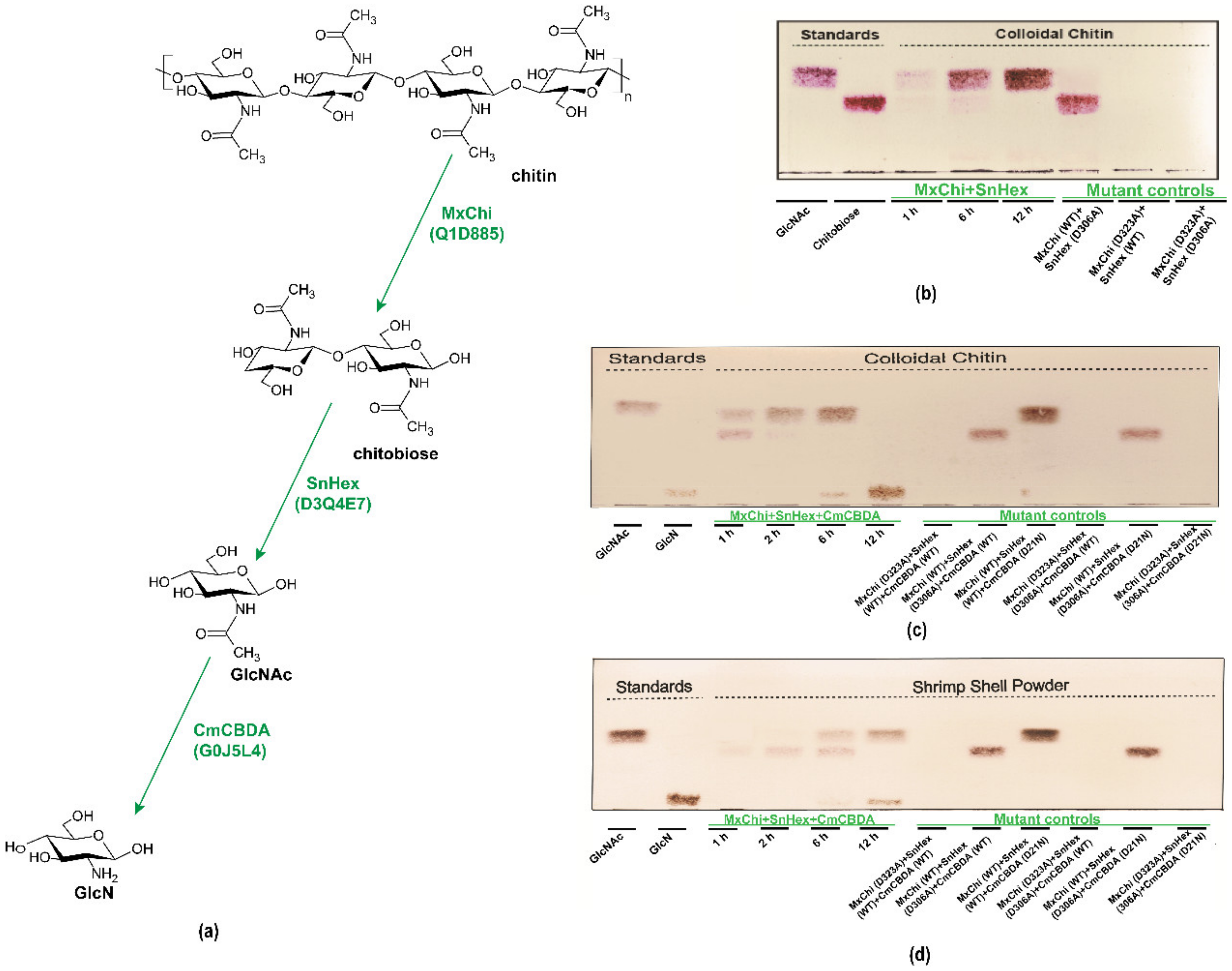
3.5. Biosynthesis of GlcN from Colloidal Chitin and Shrimp Shell
3.6. Single Factor Test Results
3.7. Optimization of GlcN Biosynthesis by Orthogonal Design
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yan, N.; Chen, X. Sustainability: Don’t waste seafood waste. Nature 2015, 524, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, C.; González Auza, L.; Koberidze, D.; Rasche, S.; Fischer, R.; Bortesi, L. Conversion of Chitin to Defined Chitosan Oligomers: Current Status and Future Prospects. Mar. Drugs 2019, 17, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanty, B.; Mohanty, U.; Pattanaik, S.S.; Panda, A.; Jena, A.K. Future prospects and trends for effective utilization of fish processing wastes in India. Innovat. Farm 2018, 3, 1–5. [Google Scholar]
- Veliz, E.A.; Martinez-Hidalgo, P.; Hirsch, A.M. Chitinase-producing bacteria and their role in biocontrol. AIMS Microbiol. 2017, 3, 689–705. [Google Scholar] [CrossRef] [PubMed]
- Frankowski, J.; Lorito, M.; Scala, F.; Schmid, R.; Berg, G.; Bahl, H. Purification and properties of two chitinolytic enzymes of Serratia plymuthica HRO-C48. Arch. Microbiol. 2001, 176, 421–426. [Google Scholar] [CrossRef]
- Huang, L.; Chen, J.; Cao, P.; Pan, H.; Ding, C.; Xiao, T.; Zhang, P.; Guo, J.; Su, Z. Anti-obese effect of glucosamine and chitosan oligosaccharide in high-fat diet-induced obese rats. Mar. Drugs 2015, 13, 2732–2756. [Google Scholar] [CrossRef] [PubMed]
- Papadaki, A.; Diamantopoulou, P.; Papanikolaou, S.; Philippoussis, A. Evaluation of Biomass and Chitin Production of Morchella Mushrooms Grown on Starch-Based Substrates. Foods 2019, 8, 239. [Google Scholar] [CrossRef] [Green Version]
- Son, Y.J.; Choi, S.Y.; Hwang, I.K.; Nho, C.W.; Kim, S.H. Could Defatted Mealworm (Tenebrio molitor) and Mealworm Oil Be Used as Food Ingredients? Foods 2020, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Yadav, M.; Goswami, P.; Paritosh, K.; Kumar, M.; Pareek, N.; Vivekanand, V. Seafood waste: A source for preparation of commercially employable chitin/chitosan materials. Bioresour. Bioprocess. 2019, 6, 8. [Google Scholar] [CrossRef]
- Kucharz, E.J.; Kovalenko, V.; Szántó, S.; Bruyère, O.; Cooper, C.; Reginster, J.-Y. A review of glucosamine for knee osteoarthritis: Why patented crystalline glucosamine sulfate should be differentiated from other glucosamines to maximize clinical outcomes. Curr. Med. Res. Opin. 2016, 32, 997–1004. [Google Scholar] [CrossRef] [Green Version]
- Kwoh, C.K.; Roemer, F.W.; Hannon, M.J.; Moore, C.E.; Jakicic, J.M.; Guermazi, A.; Green, S.M.; Evans, R.W.; Boudreau, R. Effect of Oral Glucosamine on Joint Structure in Individuals with Chronic Knee Pain: A Randomized, Placebo-Controlled Clinical Trial. Arthritis Rheumatol. 2014, 66, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Tokuyasu, K.; Mitsutomi, M.; Yamaguchi, I.; Hayashi, K.; Mori, Y. Recognition of Chitooligosaccharides and Their N-Acetyl Groups by Putative Subsites of Chitin Deacetylase from a Deuteromycete, Colletotrichum lindemuthianum. Biochemistry 2000, 39, 8837–8843. [Google Scholar] [CrossRef]
- Hajji, S.; Younes, I.; Ghorbel-Bellaaj, O.; Hajji, R.; Rinaudo, M.; Nasri, M.; Jellouli, K. Structural differences between chitin and chitosan extracted from three different marine sources. Int. J. Biol. Macromol. 2014, 65, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Dhillon, G.S. Recent trends in biological extraction of chitin from marine shell wastes: A review. Crit. Rev. Biotechnol. 2015, 35, 44–61. [Google Scholar] [CrossRef]
- Nidheesh, T.; Gaurav Kumar, P.; Suresh, P.V. Enzymatic degradation of chitosan and production of d-glucosamine by solid substrate fermentation of exo-β-d-glucosaminidase (exochitosanase) by Penicillium decumbens CFRNT15. Int. Biodeter. Biodegr. 2015, 97, 97–106. [Google Scholar] [CrossRef]
- Niu, X.; Zhou, J.S.; Wang, Y.X.; Liu, C.C.; Liu, Z.H.; Yuan, S. Heterologous Expression and Characterization of a Novel Chitinase (ChiEn1) from Coprinopsis cinerea and its Synergism in the Degradation of Chitin. J. Agric. Food Chem. 2017, 65, 6943–6956. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, D.; Nagpure, A.; Gupta, R.K. Bacterial chitinases: Properties and potential. Crit. Rev. Biotechnol. 2007, 27, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Kasprzewska, A. Plant chitinases--regulation and function. Cell Mol. Biol. Lett. 2003, 8, 809–824. [Google Scholar] [PubMed]
- Van Dyken, S.J.; Locksley, R.M. Chitins and chitinase activity in airway diseases. J. Allergy Clin. Immunol. 2018, 142, 364–369. [Google Scholar] [CrossRef]
- Pan, M.Y.; Li, J.H.; Lv, X.Q.; Du, G.C.; Liu, L. Molecular engineering of chitinase from Bacillus sp. DAU101 for enzymatic production of chitooligosaccharides. Enzyme Microb. Technol. 2019, 124, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Loc, N.H.; Huy, N.D.; Quang, H.T.; Lan, T.T.; Thu Ha, T.T. Characterisation and antifungal activity of extracellular chitinase from a biocontrol fungus, Trichoderma asperellum PQ34. Mycology 2020, 11, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Dworkin, M. Recent advances in the social and developmental biology of the myxobacteria. Microbiol. Rev. 1996, 60, 70–102. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.K.; Ye, X.F.; Liu, M.X.; Xia, C.Y.; Zhang, L.; Luo, X.; Wang, T.; Chen, Y.; Zhao, Y.Q.; Qiao, Y. A novel outer membrane β-1, 6-glucanase is deployed in the predation of fungi by myxobacteria. ISME J. 2019, 13, 2223–2235. [Google Scholar] [CrossRef]
- Zhang, W.J.; Ma, J.W.; Yan, Q.J.; Jiang, Z.Q.; Yang, S.Q. Biochemical characterization of a novel acidic chitinase with antifungal activity from Paenibacillus xylanexedens Z2–4. Int. J. Biol. Macromol. 2021, 182, 1528–1536. [Google Scholar] [CrossRef] [PubMed]
- Mahuku, G.S. A simple extraction method suitable for PCR-based analysis of plant, fungal, and bacterial DNA. Plant Mol. Biol. Rep. 2004, 22, 71–81. [Google Scholar] [CrossRef]
- Wattam, A.R.; Abraham, D.; Dalay, O.; Disz, T.L.; Driscoll, T.; Gabbard, J.L.; Gillespie, J.J.; Gough, R.; Hix, D.; Kenyon, R. PATRIC, the bacterial bioinformatics database and analysis resource. Nucleic Acids Res. 2014, 42, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Lv, Y.M.; Laborda, P.; Huang, K.; Cai, Z.P.; Wang, M.; Lu, A.M.; Doherty, C.; Liu, L.; Flitsch, S.L.; Voglmeir, J. Highly efficient and selective biocatalytic production of glucosamine from chitin. Green Chem. 2017, 19, 527–535. [Google Scholar] [CrossRef]
- Cai, Z.P.; Hagan, A.K.; Wang, M.M.; Flitsch, S.L.; Liu, L.; Voglmeir, J. 2-Pyridylfuran: A new fluorescent tag for the analysis of carbohydrates. Anal. Chem. 2014, 86, 5179–5186. [Google Scholar] [CrossRef]
- Raval, N.P.; Shah, P.U.; Ladha, D.G.; Wadhwani, P.M.; Shah, N.K. Comparative study of chitin and chitosan beads for the adsorption of hazardous anionic azo dye Congo Red from wastewater. Desalin. Water Treat. 2015, 57, 9247–9262. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, F.; Wang, T.; Lyu, Y.M.; Alteen, M.G.; Cai, Z.P.; Cui, Z.L.; Liu, L.; Voglmeir, J. Characterization of Stackebrandtia nassauensis GH 20 Beta-Hexosaminidase, a Versatile Biocatalyst for Chitobiose Degradation. Int. J. Mol. Sci. 2019, 20, 1243. [Google Scholar] [CrossRef] [Green Version]
- Eswar, N.; Webb, B.; Marti-Renom, M.A.; Madhusudhan, M.; Eramian, D.; Shen, M.-Y.; Pieper, U.; Sali, A. Comparative protein structure modeling using Modeller. Curr. Protoc. Bioinform. 2006, 15, 1–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.-F.; Guindon, S.; Lefort, V.; Lescot, M. Phylogeny. fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Van Aalten, D.; Synstad, B.; Brurberg, M.; Hough, E.; Riise, B.; Eijsink, V.; Wierenga, R. Structure of a two-domain chitotriosidase from Serratia marcescens at 1.9-Å resolution. Proc. Natl. Acad. Sci. USA 2000, 97, 5842–5847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malecki, P.H.; Bejger, M.; Rypniewski, W.; Vorgias, C.E. The Crystal Structure of a Streptomyces thermoviolaceus Thermophilic Chitinase Known for Its Refolding Efficiency. Int. J. Mol. Sci. 2020, 21, 2892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rani, T.S.; Madhuprakash, J.; Podile, A.R. Chitinase-E from Chitiniphilus shinanonensis generates chitobiose from chitin flakes. Int. J. Biol. Macromol. 2020, 163, 1037–1043. [Google Scholar] [CrossRef]
- Xie, X.H.; Fu, X.; Yan, X.Y.; Peng, W.F.; Kang, L.X. A Broad-Specificity Chitinase from Penicillium oxalicum k10 Exhibits Antifungal Activity and Biodegradation Properties of Chitin. Mar. Drugs 2021, 19, 356. [Google Scholar] [CrossRef]
- Wu, X.; Wang, J.; Shi, Y.Q.; Chen, S.; Yan, Q.J.; Jiang, Z.Q.; Jing, H. N-Acetyl-chitobiose ameliorates metabolism dysfunction through Erk/p38 MAPK and histone H3 phosphorylation in type 2 diabetes mice. J. Funct. Foods 2017, 28, 96–105. [Google Scholar] [CrossRef]
- Winkler, A.J.; Dominguez-Nuñez, J.A.; Aranaz, I.; Poza-Carrión, C.; Ramonell, K.; Somerville, S.; Berrocal-Lobo, M. Short-chain chitin oligomers: Promoters of plant growth. Mar. Drugs 2017, 15, 40. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.Q.; Fu, X.; Yan, Q.J.; Guo, Y.; Liu, Z.Q.; Jiang, Z.Q. Cloning, expression, purification and application of a novel chitinase from a thermophilic marine bacterium Paenibacillus barengoltzii. Food Chem. 2016, 192, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.L.; He, Y.M.; Wei, G.G.; Zhou, J.; Dong, W.L.; Chen, K.Q.; Ouyang, P.K. Molecular characterization of a novel chitinase CmChi1 from Chitinolyticbacter meiyuanensis SYBC-H1 and its use in N-acetyl-d-glucosamine production. Biotechnol. Biofuels 2018, 11, 179. [Google Scholar] [CrossRef]
- Stoykov, Y.M.; Pavlov, A.I.; Krastanov, A.I. Chitinase biotechnology: Production, purification, and application. Eng. Life Sci. 2015, 15, 30–38. [Google Scholar] [CrossRef]
- Hammami, I.; Siala, R.; Jridi, M.; Ktari, N.; Nasri, M.; Triki, M. Partial purification and characterization of chi IO 8, a novel antifungal chitinase produced by Bacillus cereus IO 8. J. Appl. Microbiol. 2013, 115, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Suma, K.; Podile, A.R. Chitinase A from Stenotrophomonas maltophilia shows transglycosylation and antifungal activities. Bioresour. Technol. 2013, 133, 213–220. [Google Scholar] [CrossRef]
- Xu, P.; Ni, Z.F.; Zong, M.H.; Ou, X.Y.; Yang, J.G.; Lou, W.Y. Improving the thermostability and activity of Paenibacillus pasadenensis chitinase through semi-rational design. Int. J. Biol. Macromol. 2020, 150, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.G.; Koh, H.Y.; Han, S.J.; Park, H.; Na, D.C.; Kim, I.-C.; Lee, H.K.; Yim, J.H. Expression of recombinant endochitinase from the Antarctic bacterium, Sanguibacter antarcticus KOPRI 21702 in Pichia pastoris by codon optimization. Protein Expr. Purif. 2010, 71, 108–114. [Google Scholar] [CrossRef]
- Jankiewicz, U.; Baranowski, B.; Swiontek Brzezinska, M.; Frak, M. Purification, characterization and cloning of a chitinase from Stenotrophomonas rhizophila G22. 3 Biotech 2020, 10, 16. [Google Scholar] [CrossRef]
- Abassi, S.; Emtiazi, G.; Hosseini-Abari, A.; Kim, B.G. Chitooligosaccharides and Thermostable Chitinase Against Vulvovaginal Candidiasis and Saprophyte Fungi: LC Mass Studies of Shrimp Shell Fermentation by Bacillus altitudinis. Curr. Microbiol. 2020, 77, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.; Zeng, S.Q.; Qin, X.; Sun, X.W.; Zhang, S.; Zhao, X.Y.; Yu, Z.N.; Li, L. Molecular docking and site-directed mutagenesis of a Bacillus thuringiensis chitinase to improve chitinolytic, synergistic lepidopteran-larvicidal and nematicidal activities. Int. J. Biol. Sci. 2015, 11, 304. [Google Scholar] [CrossRef] [Green Version]
- Hou, F.R.; Wang, D.L.; Ma, X.B.; Fan, L.H.; Ding, T.; Ye, X.Q.; Liu, D.H. Enhanced adsorption of Congo red using chitin suspension after sonoenzymolysis. Ultrason. Sonochem. 2021, 70, 105327. [Google Scholar] [CrossRef] [PubMed]
- Vaaje-Kolstad, G.; Houston, D.R.; Rao, F.V.; Peter, M.G.; Synstad, B.; Van Aalten, D.M.; Eijsink, V.G. Structure of the D142N mutant of the family 18 chitinase ChiB from Serratia marcescens and its complex with allosamidin. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2004, 1696, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Jiang, W.X.; Zhang, Y.S.; Cao, H.Y.; Zhang, Y.; Chen, X.L.; Li, C.Y.; Wang, P.; Zhang, Y.Z.; Song, X.Y. Structural insight into chitin degradation and thermostability of a novel endochitinase from the glycoside hydrolase family 18. Front. Microbiol. 2019, 10, 2457. [Google Scholar] [CrossRef]
- Kumar, M.; Dangayach, P.; Pareek, N. Enhanced glucosamine production through synergistic action of Aspergillus terreus chitozymes. J. Clean. Prod. 2020, 262, 121363. [Google Scholar] [CrossRef]
- Bao, J.J.; Liu, N.; Zhu, L.Y.; Xu, Q.; Huang, H.; Jiang, L. Programming a Biofilm-Mediated Multienzyme-Assembly-Cascade System for the Biocatalytic Production of Glucosamine from Chitin. J. Agric. Food Chem. 2018, 66, 8061–8068. [Google Scholar] [CrossRef]
- Wang, J.; Yu, X.H.; Li, F.W.; Wang, D.J.; Bu, W.L.; Ding, X.X.; Shang, Y.L. Optimization of semi-continuous fermentation conditions and preservation of Flammulina velutipes liquid strains. J. Food Process. Preserv. 2019, 43, e14010. [Google Scholar] [CrossRef]
- Zhou, C.S.; Hu, J.L.; Ma, H.L.; Yagoub, A.E.A.; Yu, X.J.; Owusu, J.; Ma, H.Y.; Qin, X. Antioxidant peptides from corn gluten meal: Orthogonal design evaluation. Food Chem. 2015, 187, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Li, H.L.; Chen, X.D.; Xiong, L.; Luo, M.T.; Chen, X.F.; Wang, C.; Huang, C.; Chen, X.D. Stepwise enzymatic hydrolysis of alkaline oxidation treated sugarcane bagasse for the co-production of functional xylo-oligosaccharides and fermentable sugars. Bioresour. Technol. 2019, 275, 345–351. [Google Scholar] [CrossRef]
- Song, H.B.; Zhao, G.H.; Zhang, M.; Bi, R.M.; Meng, X.H.; Song, J.L.; Wang, B.; Liu, J.; Liu, L.; Lyu, Y.M.; et al. Optimization of the UDP-Xyl biocatalytic synthesis from Crassostrea gigas by orthogonal design method. Protein Expr. Purif. 2021, 190, 106002. [Google Scholar] [CrossRef]
- Mojarrad, J.S.; Nemati, M.; Valizadeh, H.; Ansarin, M.; Bourbour, S. Preparation of glucosamine from exoskeleton of shrimp and predicting production yield by response surface methodology. J. Agric. Food Chem. 2007, 55, 2246–2250. [Google Scholar] [CrossRef]
- Benavente, M.; Arias, S.; Moreno, L.; Martínez, J. Production of glucosamine hydrochloride from crustacean shell. J. Pharm. Pharmacol. 2015, 3, 20–26. [Google Scholar]
- Faraco, G.; Brea, D.; Garcia-Bonilla, L.; Wang, G.; Racchumi, G.; Chang, H.; Buendia, I.; Santisteban, M.M.; Segarra, S.G.; Koizumi, K.; et al. Dietary salt promotes neurovascular and cognitive dysfunction through a gut-initiated TH17 response. Nat. Neurosci. 2018, 21, 240–249. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level | Factors | ||
|---|---|---|---|
| pH(A) | Temperature(B) | Enzyme Amount Ratio (C) (MxChi: SnHex: CmCBDA, μg) | |
| 1 | 7 | 32 | 150: 450: 2000 |
| 2 | 8 | 37 | 150: 450: 2250 |
| 3 | 9 | 42 | 150: 450: 2500 |
| Exp. Number | Factors | Product (mg) | ||
|---|---|---|---|---|
| pH(A) | Temperature(B) (℃) | Enzyme Amount Ratio (C) MxChi: SnHex: CmCBDA, μg) | ||
| 1 | 7 | 32 | 150: 450: 2500 | 0.96 |
| 2 | 7 | 37 | 150: 450: 2250 | 2.02 |
| 3 | 7 | 42 | 150: 450: 2000 | 1.57 |
| 4 | 8 | 32 | 150: 450: 2250 | 0.88 |
| 5 | 8 | 37 | 150: 450: 2000 | 2.48 |
| 6 | 8 | 42 | 150: 450: 2500 | 1.99 |
| 7 | 9 | 32 | 150: 450: 2000 | 0.45 |
| 8 | 9 | 37 | 150: 450: 2500 | 1.67 |
| 9 | 9 | 42 | 150: 450: 2250 | 1.21 |
| K1 | 4.55 | 2.29 | 4.50 | |
| K2 | 5.35 | 6.17 | 4.11 | |
| K3 | 3.33 | 4.77 | 4.62 | |
| 1.51 | 0.76 | 1.50 | ||
| 1.78 | 2.06 | 1.37 | ||
| 1.11 | 1.59 | 1.54 | ||
| R | 0.76 | 1.30 | 0.17 | |
| Best level | A2B2C3 | |||
| Source of Variation | Sum of Square | Variance | Mean Square | F-Value | p-Value | Significant |
|---|---|---|---|---|---|---|
| Factor A | 0.41 | 1 | 0.41 | 32.03 | 0.0008 | ** |
| Factor B | 1.17 | 1 | 1.17 | 91.54 | <0.0001 | *** |
| Factor C | 0.18 | 1 | 0.18 | 14.31 | 0.0069 | ** |
| Error | 0.038 | 4 | 9.57 × 10−3 | |||
| Sum | 1.798 | 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, Y.; Zheng, F.; Qiu, C.; Wang, M.; Wang, D.; Zhang, X.; Voglmeir, J.; Liu, L.; Yu, X. Heterologous Expression of a Thermostable Chitinase from Myxococcus xanthus and Its Application for High Yield Production of Glucosamine from Shrimp Shell. Foods 2021, 10, 2808. https://doi.org/10.3390/foods10112808
Lyu Y, Zheng F, Qiu C, Wang M, Wang D, Zhang X, Voglmeir J, Liu L, Yu X. Heterologous Expression of a Thermostable Chitinase from Myxococcus xanthus and Its Application for High Yield Production of Glucosamine from Shrimp Shell. Foods. 2021; 10(11):2808. https://doi.org/10.3390/foods10112808
Chicago/Turabian StyleLyu, Yongmei, Feng Zheng, Chuanxing Qiu, Meng Wang, Dujun Wang, Xiaoyang Zhang, Josef Voglmeir, Li Liu, and Xiaohong Yu. 2021. "Heterologous Expression of a Thermostable Chitinase from Myxococcus xanthus and Its Application for High Yield Production of Glucosamine from Shrimp Shell" Foods 10, no. 11: 2808. https://doi.org/10.3390/foods10112808
APA StyleLyu, Y., Zheng, F., Qiu, C., Wang, M., Wang, D., Zhang, X., Voglmeir, J., Liu, L., & Yu, X. (2021). Heterologous Expression of a Thermostable Chitinase from Myxococcus xanthus and Its Application for High Yield Production of Glucosamine from Shrimp Shell. Foods, 10(11), 2808. https://doi.org/10.3390/foods10112808