
How Safe to Eat Are Raw Bivalves? Host Pathogenic and Public Health Concern Microbes within Mussels, Oysters, and Clams in Greek Markets

 ,
,  ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. E. coli and Salmonella spp. Detection
2.3. Microbiological Culture of Tissues and Molecular Identification of Cultures
2.4. Molecular Examination for the Presence of Marteilia, Bonamia, and Vibrio spp.
2.5. Sequencing and Phylogenetic Analysis
2.6. Molecular Investigation of Foodborne Viruses
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tacon, A.G.J.; Metian, M. Fish Matters: Importance of Aquatic Foods in Human Nutrition and Global Food Supply. Rev. Fish. Sci. 2013, 21, 22–38. [Google Scholar] [CrossRef]
- Aakre, I.; Bøkevoll, A.; Chaira, J.; Bouthir, F.Z.; Frantzen, S.; Kausland, A.; Kjellevold, M. Variation in nutrient composition of seafood from North West Africa: Implications for food and nutrition security. Foods 2020, 9, 1516. [Google Scholar] [CrossRef] [PubMed]
- Orban, E.; Di Lena, G.; Nevigato, T.; Casini, I.; Caproni, R.; Santaroni, G.; Giulini, G. Nutritional and commercial quality of the striped venus clam, Chamelea gallina, from the Adriatic sea. Food Chem. 2007, 101, 1063–1070. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J. Omega-3 fatty acids and cardiovascular disease: New recommendations from the American Heart Association. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 151–152. [Google Scholar] [CrossRef]
- Sidhu, K.S. Health benefits and potential risks related to consumption of fish or fish oil. Regul. Toxicol. Pharmacol. 2003, 38, 336–344. [Google Scholar] [CrossRef]
- Biandolino, F.; Di Leo, A.; Parlapiano, I.; Papa, L.; Giandomenico, S.; Spada, L.; Prato, E. Nutritional quality of edible marine bivalves from the southern coast of Italy, Mediterranean Sea. Pol. J. Food Nutr. Sci. 2019, 69, 71–81. [Google Scholar] [CrossRef]
- Theodorou, J.A.; Viaene, J.; Sorgeloos, P.; Tzovenis, I. Production and marketing trends of the cultured mediterranean mussel Μytilus galloprovincialis lamarck 1819, in Greece. J. Shellfish Res. 2011, 30, 859–874. [Google Scholar] [CrossRef]
- Froehlich, H.E.; Gentry, R.R.; Halpern, B.S. Global change in marine aquaculture production potential under climate change. Nat. Ecol. Evol. 2018, 2, 1745–1750. [Google Scholar] [CrossRef]
- Anestis, A.; Lazou, A.; Pörtner, H.O.; Michaelidis, B. Behavioral, metabolic, and molecular stress responses of marine bivalve Mytilus galloprovincialis during long-term acclimation at increasing ambient temperature. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, 911–921. [Google Scholar] [CrossRef]
- Amorim, V.E.; Gonçalves, O.; Capela, R.; Fernández-Boo, S.; Oliveira, M.; Dolbeth, M.; Arenas, F.; Cardoso, P.G. Immunological and oxidative stress responses of the bivalve Scrobicularia plana to distinct patterns of heatwaves. Fish Shellfish Immunol. 2020, 106, 1067–1077. [Google Scholar] [CrossRef]
- Matozzo, V.; Marin, M.G. Bivalve immune responses and climate changes: Is there a relationship? Invertebr. Surviv. J. 2011, 8, 70–77. [Google Scholar]
- Matozzo, V.; Chinellato, A.; Munari, M.; Finos, L.; Bressan, M.; Marin, M.G. First evidence of immunomodulation in bivalves under seawater acidification and increased temperature. PLoS ONE 2012, 7, e33820. [Google Scholar] [CrossRef]
- Lattos, A.; Giantsis, I.A.; Karagiannis, D.; Michaelidis, B. First detection of the invasive Haplosporidian and Mycobacteria parasites hosting the endangered bivalve Pinna nobilis in Thermaikos Gulf, North Greece. Mar. Environ. Res. 2020, 155, 104889. [Google Scholar] [CrossRef]
- Lattos, A.; Giantsis, I.A.; Karagiannis, D.; Theodorou, J.A.; Michaelidis, B. Gut Symbiotic Microbial Communities in the IUCN Critically Endangered Pinna nobilis Suffering from Mass Mortalities, Revealed by 16S rRNA Amplicon NGS. Pathogens 2020, 9, 1002. [Google Scholar] [CrossRef] [PubMed]
- Lattos, A.; Bitchava, K.; Giantsis, I.A.; Theodorou, J.A.; Batargias, C.; Michaelidis, B. The implication of vibrio bacteria in the winter mortalities of the critically endangered Pinna nobilis. Microorganisms 2021, 9, 922. [Google Scholar] [CrossRef]
- Lattos, A.; Feidantsis, K.; Georgoulis, I.; Giantsis, I.A.; Karagiannis, D.; Theodorou, J.A.; Staikou, A.; Michaelidis, B. Pathophysiological Responses of Pinna nobilis Individuals Enlightens the Etiology of Mass Mortality Situation in the Mediterranean Populations. Cells 2021, 10, 2838. [Google Scholar] [CrossRef]
- Virvilis, C.; Angelidis, P. Presence of the parasite Marteilia sp. in the flat oyster (Ostrea edulis L) in Greece. Aquaculture 2006, 259, 1–5. [Google Scholar] [CrossRef]
- Karagiannis, D.; Angelidis, P. Infection of cultured mussels Mytilus galloprovincialis by the protozoan Marteilia sp. in the Thermaikos Gulf (N Greece). Bull. Eur. Assoc. Fish Pathol. 2007, 27, 131–141. [Google Scholar]
- Karagiannis, D.; Michaelidis, B.; Theodoridis, A.; Angelidis, P.; Feidantsis, K.; Staikou, A. Field studies on the effects of Marteilia sp. on growth of mussel Mytilus galloprovincialis in Thermaikos Gulf. Mar. Environ. Res. 2018, 142, 116–123. [Google Scholar] [CrossRef]
- Park, K.; Mok, J.S.; Ryu, A.R.; Kwon, J.Y.; Ham, I.T.; Shim, K.B. Occurrence and virulence of Vibrio parahaemolyticus isolated from seawater and bivalve shellfish of the Gyeongnam coast, Korea, in 2004–2016. Mar. Pollut. Bull. 2018, 137, 382–387. [Google Scholar] [CrossRef]
- Ryu, A.R.; Mok, J.S.; Lee, D.E.; Kwon, J.Y.; Park, K. Occurrence, virulence, and antimicrobial resistance of Vibrio parahaemolyticus isolated from bivalve shellfish farms along the southern coast of Korea. Environ. Sci. Pollut. Res. 2019, 26, 21034–21043. [Google Scholar] [CrossRef]
- Ghribi, F.; Boussoufa, D.; Aouini, F.; Bejaoui, S.; Chetoui, I.; Bouaziz, M.; El Cafsi, M. Nutritional quality traits of raw and cooked Ark shell (Bivalvia: Arcidae): Balancing the benefits and risks of seafood consumption. J. Food Sci. Technol. 2021, 58, 3346–3356. [Google Scholar] [CrossRef] [PubMed]
- Newell, D.G.; Koopmans, M.; Verhoef, L.; Duizer, E.; Aidara-Kane, A.; Sprong, H.; Opsteegh, M.; Langelaar, M.; Threfall, J.; Scheutz, F.; et al. Food-borne diseases—The challenges of 20years ago still persist while new ones continue to emerge. Int. J. Food Microbiol. 2010, 139, S3–S15. [Google Scholar] [CrossRef] [PubMed]
- Potasman, I.; Paz, A.; Odeh, M. Infectious outbreaks associated with bivalve shellfish consumption: A worldwide perspective. Clin. Infect. Dis. 2002, 35, 921–928. [Google Scholar] [CrossRef]
- Leight, A.K.; Crump, B.C.; Hood, R.R. Assessment of fecal indicator bacteria and potential pathogen co-occurrence at a shellfish growing area. Front. Microbiol. 2018, 9, 384. [Google Scholar] [CrossRef] [PubMed]
- Balière, C.; Rincé, A.; Thevenot, D.; Gourmelon, M. Successful detection of pathogenic Shiga-toxin-producing Escherichia coli in shellfish, environmental waters and sediment using the ISO/TS-13136 method. Lett. Appl. Microbiol. 2015, 60, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Touchon, M.; Hoede, C.; Tenaillon, O.; Barbe, V.; Baeriswyl, S.; Bidet, P.; Bingen, E.; Bonacorsi, S.; Bouchier, C.; Bouvet, O.; et al. Organised genome dynamics in the Escherichia coli species results in highly diverse adaptive paths. PLoS Genet. 2009, 5, e1000344. [Google Scholar] [CrossRef]
- Kanayama, A.; Yahata, Y.; Arima, Y.; Takahashi, T.; Saitoh, T.; Kanou, K.; Kawabata, K.; Sunagawa, T.; Matsui, T.; Oishi, K. Enterohemorrhagic Escherichia coli outbreaks related to childcare facilities in Japan, 2010–2013. BMC Infect. Dis. 2015, 15, 539. [Google Scholar] [CrossRef]
- Malham, S.K.; Rajko-Nenow, P.; Howlett, E.; Tuson, K.E.; Perkins, T.L.; Pallett, D.W.; Wang, H.; Jago, C.F.; Jones, D.L.; McDonald, J.E. The interaction of human microbial pathogens, particulate material and nutrients in estuarine environments and their impacts on recreational and shellfish waters. Environ. Sci. Process. Impacts 2014, 16, 2145–2155. [Google Scholar] [CrossRef]
- ECDC; ECFDPC. Salmonellosis—Annual Epidemiological Report for 2017. Eur. Cent. Dis. Prev. Control 2019, 8, 1–8. [Google Scholar]
- Catalao Dionisio, L.P.; Joao, M.; Soares Ferreiro, V.; Leonor Fidalgo, M.; García Rosado, M.E.; Borrego, J.J. Occurrence of Salmonella spp in estuarine and coastal waters of Portugal. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2000, 78, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Ayers, T.; Mahon, B.E.; Swerdlow, D.L. Epidemiology of seafood-associated infections in the United States. Clin. Microbiol. Rev. 2010, 23, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Collin, B.; Rehnstam-Holm, A.S. Occurrence and potential pathogenesis of Vibrio cholerae, Vibrio parahaemolyticus and Vibrio vulnificus on the South Coast of Sweden. FEMS Microbiol. Ecol. 2011, 78, 306–313. [Google Scholar] [CrossRef]
- Park, K.; Mok, J.S.; Kwon, J.Y.; Ryu, A.R.; Kim, S.H.; Lee, H.J. Food-borne outbreaks, distributions, virulence, and antibiotic resistance profiles of Vibrio parahaemolyticus in Korea from 2003 to 2016: A review. Fish. Aquat. Sci. 2018, 21, 3. [Google Scholar] [CrossRef]
- Letchumanan, V.; Chan, K.G.; Lee, L.H. Vibrio parahaemolyticus: A review on the pathogenesis, prevalence, and advance molecular identification techniques. Front. Microbiol. 2014, 5, 705. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.H.; Shin, Y.J.; Jang, S.C.; Yu, H.S.; Kim, S.K.; An, S.; Park, K.; So, J.S. Characterization of Vibrio parahaemolyticus isolated from oysters in Korea: Resistance to various antibiotics and prevalence of virulence genes. Mar. Pollut. Bull. 2017, 118, 261–266. [Google Scholar] [CrossRef]
- Faruque, S.M.; Nair, G.B.; Mekalanos, J.J. Genetics of stress adaptation and virulence in toxigenic Vibrio cholerae. DNA Cell Biol. 2004, 23, 723–741. [Google Scholar] [CrossRef]
- Meena, B.; Anburajan, L.; Sathish, T.; Das, A.K.; Vinithkumar, N.V.; Kirubagaran, R.; Dharani, G. Studies on diversity of Vibrio sp. and the prevalence of hapA, tcpI, st, rtxA&C, acfB, hlyA, ctxA, ompU and toxR genes in environmental strains of Vibrio cholerae from Port Blair bays of South Andaman, India. Mar. Pollut. Bull. 2019, 144, 105–116. [Google Scholar] [CrossRef]
- Paranjpye, R.N.; Johnson, A.B.; Baxter, A.E.; Strom, M.S. Role of type IV pilins in persistence of Vibrio vulnificus in Crassostrea virginica oysters. Appl. Environ. Microbiol. 2007, 73, 5041–5044. [Google Scholar] [CrossRef] [PubMed]
- Arias, C.R.; Macián, M.C.; Aznar, R.; Garay, E.; Pujalte, M.J. Low incidence of Vibrio vulnificus among Vibrio isolates from sea water and shellfish of the western Mediterranean coast. J. Appl. Microbiol. 1999, 86, 125–134. [Google Scholar] [CrossRef]
- Yano, Y.; Yokoyama, M.; Satomi, M.; Oikawa, H.; Chen, S.S. Occurrence of Vibrio vulnificus in fish and shellfish available from markets in China. J. Food Prot. 2004, 67, 1617–1623. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, E.; Falzano, L.; Fiorentini, C.; Pianetti, A.; Baffone, W.; Fabbri, A.; Matarrese, P.; Casiere, A.; Katouli, M.; Kühn, I.; et al. Occurrence, diversity, and pathogenicity of halophilic Vibrio spp. and non-O1 Vibrio cholerae from estuarine waters along the Italian Adriatic coast. Appl. Environ. Microbiol. 1999, 65, 2748–2753. [Google Scholar] [CrossRef] [PubMed]
- Baffone, W.; Pianetti, A.; Bruscolini, F.; Barbieri, E.; Citterio, B. Occurrence and expression of virulence-related properties of Vibrio species isolated from widely consumed seafood products. Int. J. Food Microbiol. 2000, 54, 9–18. [Google Scholar] [CrossRef]
- Mustapha Ennaji, M.; Mustapha, S.; Moulay Mustapha, E.; Nozha, C. Vibrio alginolyticus: An Emerging Pathogen of Foodborne Diseases. Int. J. Sci. Technol. 2013, 2, 302–309. [Google Scholar]
- Selvin, J.; Lipton, A.P. Vibrio alginolyticus associated with white spot disease of Penaeus monodon. Dis. Aquat. Organ. 2003, 57, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Zhai, S.; Li, X.; Tian, J.; Li, Q.; Shan, H.; Liu, S. Identification of Vibrio alginolyticus as a causative pathogen associated with mass summer mortality of the Pacific Oyster (Crassostrea gigas) in China. Aquaculture 2021, 535, 736363. [Google Scholar] [CrossRef]
- Masini, L.; De Grandis, G.; Principi, F.; Mengarelli, C.; Ottaviani, D. Research and characterization of pathogenic vibrios from bathing water along the Conero Riviera (Central Italy). Water Res. 2007, 41, 4031–4040. [Google Scholar] [CrossRef]
- Lafisca, A.; Pereira, C.S.; Giaccone, V.; Rodrigues, D.D.P. Enzymatic characterization of Vibrio alginolyticus strains isolated from bivalves harvested at Venice Lagoon (Italy) and Guanabara Bay (Brazil). Rev. Inst. Med. Trop. Sao Paulo 2008, 50, 199–202. [Google Scholar] [CrossRef]
- EFSA Panel on Animal Health and Welfare (AHAW). Scientific Opinion on Review of the European Union Summary Report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2009 and 2010 specifically for the data related to bovine tuberculosis, Echinococcus, Q fever, brucellosis and non-food borne diseases. EFSA J. 2012, 10, 2765. [Google Scholar] [CrossRef]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States-Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Tam, C.C.; Rodrigues, L.C.; Viviani, L.; Dodds, J.P.; Evans, M.R.; Hunter, P.R.; Gray, J.J.; Letley, L.H.; Rait, G.; Tompkins, D.S.; et al. Longitudinal study of infectious intestinal disease in the UK (IID2 study): Incidence in the community and presenting to general practice. Gut 2012, 61, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Al-Thani, A.; Baris, M.; Al-Lawati, N.; Al-Dhahry, S. Characterising the aetiology of severe acute gastroenteritis among patients visiting a hospital in Qatar using real-time polymerase chain reaction. BMC Infect. Dis. 2013, 13, 329. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, M. Progress in understanding norovirus epidemiology. Curr. Opin. Infect. Dis. 2008, 21, 544–552. [Google Scholar] [CrossRef]
- Patel, M.M.; Widdowson, M.A.; Glass, R.I.; Akazawa, K.; Vinjé, J.; Parashar, U.D. Systematic literature review of role of noroviruses in sporadic gastroenteritis. Emerg. Infect. Dis. 2008, 14, 1224–1231. [Google Scholar] [CrossRef]
- Vinjé, J. Advances in laboratory methods for detection and typing of norovirus. J. Clin. Microbiol. 2015, 53, 373–381. [Google Scholar] [CrossRef]
- Chhabra, P.; de Graaf, M.; Parra, G.I.; Chan, M.C.W.; Green, K.; Martella, V.; Wang, Q.; White, P.A.; Katayama, K.; Vennema, H.; et al. Updated classification of norovirus genogroups and genotypes. J. Gen. Virol. 2019, 100, 1393–1406. [Google Scholar] [CrossRef]
- Chan, M.C.; Roy, S.; Bonifacio, J.; Zhang, L.; Chhabra, P.; Chan, J.C.M.; Celma, C.; Igoy, M.A.; Lau, S.; Mohammad, K.N.; et al. Detection of Norovirus Variant. Emerg. Infect. Dis. 2021, 27, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.G.; Kuroiwa, T.; Kaneshi, K.; Ueda, Y.; Nakaya, S.; Nishimura, S.; Yamamoto, A.; Sugita, K.; Nishimura, T.; Yagyu, F.; et al. Changing distribution of norovirus genotypes and genetic analysis of recombinant GIIb among infants and children with diarrhea in Japan. J. Med. Virol. 2006, 78, 971–978. [Google Scholar] [CrossRef]
- Lees, D. International standardisation of a method for detection of human pathogenic viruses in molluscan shellfish. Food Environ. Virol. 2010, 2, 146–155. [Google Scholar] [CrossRef]
- Le Guyader, F.S.; Le Saux, J.C.; Ambert-Balay, K.; Krol, J.; Serais, O.; Parnaudeau, S.; Giraudon, H.; Delmas, G.; Pommepuy, M.; Pothier, P.; et al. Aichi virus, norovirus, astrovirus, enterovirus, and rotavirus involved in clinical cases from a French oyster-related gastroenteritis outbreak. J. Clin. Microbiol. 2008, 46, 4011–4017. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, M.; Duizer, E. Foodborne viruses: An emerging problem. Int. J. Food Microbiol. 2004, 90, 23–41. [Google Scholar] [CrossRef]
- Prato, R.; Lopalco, P.L.; Chironna, M.; Barbuti, G.; Germinario, C.; Quarto, M. Norovirus gastroenteritis general outbreak associated with raw shellfish consumption in South Italy. BMC Infect. Dis. 2004, 4, 37. [Google Scholar] [CrossRef]
- Guillois-Becel, Y.; Couturier, E.; Le Saux, J.C.; Roque-Afonso, A.M.; Le Guyader, F.S.; Le Goas, A.; Pernes, J.; Le Bechec, S.; Briand, A.; Robert, C.; et al. An oyster-associated hepatitis A outbreak in France in 2007. Eurosurveillance 2009, 14, 19144. [Google Scholar] [CrossRef]
- Rippey, S.R. Infectious diseases associated with molluscan shellfish consumption. Clin. Microbiol. Rev. 1994, 7, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Rohayem, J. Norovirus seasonality and the potential impact of climate change. Clin. Microbiol. Infect. 2009, 15, 524–527. [Google Scholar] [CrossRef]
- International Organization of Standardization (ISO). ISO 6579-1: Microbiology of the Food Chain-Horizontal Method for the Detection and Enumeration and Serotyping of Salmonella spp.—Part 1: Detection of Salmonella spp.; ISO—International Organization of Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- International Organization of Standardization (ISO). ISO 16649-3: Microbiology of the Food Chain-Horizontal Method for the Enumeration of Betaglucuronidase-Positive Escherichia Coli—Part 3: Detection and Most Probable Number Technique Using 5-bromo-4-chloro-3-indolyl-ß-D-glucuronide; ISO—International Organization of Standardization: Geneva, Switzerland, 2015. [Google Scholar]
- International Organization of Standardization (ISO). ISO 7218: Microbiology of Food and Animal Feeding Stuffs—General Requirements and Guidance for Microbiological Examinations; ISO—International Organization for Standardization: Geneva, Switzerland, 2007. [Google Scholar]
- International Organization of Standardization (ISO). ISO 21872-1: Microbiology of Food and Animal Feeding Stuffs-Horizontal Method for the Detection of Potentially Enteropathogenic Vibrio spp.—Part 1: Detection of Vibrio Parahaemolyticus and Vibrio Cholerae; ISO—International Organization of Standardization: Geneva, Switzerland, 2007. [Google Scholar]
- International Organization of Standardization (ISO). ISO 21872-2: Microbiology of Food and Animal Feeding Stuffs-Horizontal Method for the Detection of Potentially Enteropathogenic Vibrio spp.—Part 2: Detection of Species Other Than Vibrio Parahaemolyticus and Vibrio Cholerae; ISO—International Organization of Standardization: Geneva, Switzerland, 2007. [Google Scholar]
- Le Roux, F.; Audemard, C.; Barnaud, A.; Berthe, F. DNA probes as potential tools for the detection of Marteilia refringens. Mar. Biotechnol. 1999, 1, 588–597. [Google Scholar] [CrossRef]
- Carnegie, R.B.; Burreson, E.M.; Mike Hine, P.; Stokes, N.A.; Audemard, C.; Bishop, M.J.; Peterson, C.H. Bonamia perspora sp. (Haplosporidia), a parasite of the oyster Ostreola equestris, is the first Bonamia species known to produce spores. J. Eukaryot. Microbiol. 2006, 53, 232–245. [Google Scholar] [CrossRef]
- Xu, Y.G.; Sun, L.M.; Wang, Y.S.; Chen, P.P.; Liu, Z.M.; Li, Y.J.; Tang, L.J. Simultaneous detection of Vibrio cholerae, Vibrio alginolyticus, Vibrio parahaemolyticus and Vibrio vulnificus in seafood using dual priming oligonucleotide (DPO) system-based multiplex PCR assay. Food Control 2017, 71, 64–70. [Google Scholar] [CrossRef]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular evolutionary genetics analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, T.; Kojima, S.; Shinohara, M.; Uchida, K.; Fukushi, S.; Hoshino, F.B.; Takeda, N.; Katayama, K. Broadly reactive and highly sensitive assay for Norwalk-like viruses based on real-time quantitative reverse transcription-PCR. J. Clin. Microbiol. 2003, 41, 1548–1557. [Google Scholar] [CrossRef] [PubMed]
- Brehm, T.T.; Berneking, L.; Rohde, H.; Chistner, M.; Schlickewei, C.; Sena Martins, M.; Schmiedel, S. Wound infection with Vibrio harveyi following a traumatic leg amputation after a motorboat propeller injury in Mallorca, Spain: A case report and review of literature. BMC Infect. Dis. 2020, 20, 104. [Google Scholar] [CrossRef] [PubMed]
- Vezzulli, L.; Baker-Austin, C.; Kirschner, A.; Pruzzo, C.; Martinez-Urtaza, J. Global emergence of environmental non-O1/O139 Vibrio cholerae infections linked with climate change: A neglected research field? Environ. Microbiol. 2020, 22, 4342–4355. [Google Scholar] [CrossRef]
- Su, Y.C.; Liu, C. Vibrio parahaemolyticus: A concern of seafood safety. Food Microbiol. 2007, 24, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Nordio, D.; Khtikian, N.; Andrews, S.; Bertotto, D.; Leask, K.; Green, T. Adaption potential of Crassostrea gigas to ocean acidification and disease caused by Vibrio harveyi. ICES J. Mar. Sci. 2021, 78, 360–367. [Google Scholar] [CrossRef]
- Pasharawipas, T.; Thaikua, S.; Sriurairatana, S.; Ruangpan, L.; Direkbusarakum, S.; Manopvisetcharean, J.; Flegel, T.W. Partial characterization of a novel bacteriophage of Vibrio harveyi isolated from shrimp culture ponds in Thailand. Virus Res. 2005, 114, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Alavandi, S.V.; Manoranjita, V.; Vijayan, K.K.; Kalaimani, N.; Santiago, T.C. Phenotypic and molecular typing of Vibrio harveyi isolates and their pathogenicity to tiger shrimp larvae. Lett. Appl. Microbiol. 2006, 43, 566–570. [Google Scholar] [CrossRef]
- Defoirdt, T.; Sorgeloos, P. Monitoring of Vibrio harveyi quorum sensing activity in real time during infection of brine shrimp larvae. ISME J. 2012, 6, 2314–2319. [Google Scholar] [CrossRef]
- Zhou, J.; Fang, W.; Yang, X.; Zhou, S.; Hu, L.; Li, X.; Qi, X.; Su, H.; Xie, L. A nonluminescent and highly virulent Vibrio harveyi strain is associated with “bacterial white tail disease” of Litopenaeus vannamei shrimp. PLoS ONE 2012, 7, 19–22. [Google Scholar] [CrossRef]
- Wei, Z.; Xin, L.; Zhang, W.; Bai, C.; Wang, C.; Li, C. Isolation and characterization of Vibrio harveyi as a major pathogen associated with mass mortalities of ark clam, Scapharca broughtonii, in summer. Aquaculture 2019, 511, 734248. [Google Scholar] [CrossRef]
- Travers, M.A.; Le Goïc, N.; Huchette, S.; Koken, M.; Paillard, C. Summer immune depression associated with increased susceptibility of the European abalone, Haliotis tuberculata to Vibrio harveyi infection. Fish Shellfish Immunol. 2008, 25, 800–808. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Xu, L.; Chen, H.; Liu, S.; Guo, Z.; Cheng, C.; Ma, H.; Feng, J. Prevalence, virulence genes, and antimicrobial resistance of Vibrio species isolated from diseased marine fish in South China. Sci. Rep. 2020, 10, 14329. [Google Scholar] [CrossRef] [PubMed]
- Firmino, J.; Furones, M.D.; Andree, K.B.; Sarasquete, C.; Ortiz-Delgado, J.B.; Asencio-Alcudia, G.; Gisbert, E. Contrasting outcomes of Vibrio harveyi pathogenicity in gilthead seabream, Sparus aurata and European seabass, Dicentrachus labrax. Aquaculture 2019, 511, 734210. [Google Scholar] [CrossRef]
- Mougin, J.; Roquigny, R.; Flahaut, C.; Bonnin-Jusserand, M.; Grard, T.; Le Bris, C. Abundance and spatial patterns over time of Vibrionaceae and Vibrio harveyi in water and biofilm from a seabass aquaculture facility. Aquaculture 2021, 542, 736862. [Google Scholar] [CrossRef]
- Del Gigia-Aguirre, L.; Sánchez-Yebra-Romera, W.; García-Muñoz, S.; Rodríguez-Maresca, M. First description of wound infection with Vibrio harveyi in Spain. New Microbes New Infect. 2017, 19, 15–16. [Google Scholar] [CrossRef] [PubMed]
- Rivas, A.J.; Lemos, M.L.; Osorio, C.R. Photobacterium damselae subsp. Damselae, a bacterium pathogenic for marine animals and humans. Front. Microbiol. 2013, 4, 283. [Google Scholar] [CrossRef]
- Lozano-león, A.; Osorio, C.R.; Martínez-urtaza, J.; Magariños, B. Occurrence of Photobacterium damselae subsp. damselae in bivalve molluscs from Northwest Spain. Bull. Eur. Ass. Fish Pathol. 2003, 23, 40–44. [Google Scholar]
- Chiu, T.H.; Kao, L.Y.; Chen, M.L. Antibiotic resistance and molecular typing of Photobacterium damselae subsp. damselae, isolated from seafood. J. Appl. Microbiol. 2013, 114, 1184–1192. [Google Scholar] [CrossRef]
- Grimes, D.J.; Brayton, P.; Colwell, R.R.; Gruber, S.H. Vibrios as Autochthonous Flora of Neritic Sharks. Syst. Appl. Microbiol. 1985, 6, 221–226. [Google Scholar] [CrossRef]
- Morris, J.G.; Wilson, R.; Hollis, D.G.; Weaver, R.E.; Miller, H.G.; Tacket, C.O.; Hickman, F.W.; Blake, P.A. Illness Caused By Vibrio damsela and Vibrio hollisae. Lancet 1982, 319, 1294–1297. [Google Scholar] [CrossRef]
- Smith, S.K.; Sutton, D.C.; Fuerst, J.A.; Reichelt, J.L. Evaluation of the genus Listonella and reassignment of Listonella damsela (love et al.) MacDonell and Colwell to the genus Photobacterium as Photobacterium damsela comb. nov. with an emended description. Int. J. Syst. Bacteriol. 1991, 41, 529–534. [Google Scholar] [CrossRef]
- Gauthier, G.; Lafay, B.; Ruimy, R.; Breittmayer, V.; Nicolas, J.L.; Gauthier, M.; Christen, R. Small-subunit rRNA sequences and whole DNA relatedness concur for the reassignment of Pasteurella piscicida (Snieszko et al.) Janssen and Surgalla to the genus Photobacterium as Photobacterium damsela subsp. piscicida comb. nov. Int. J. Syst. Bacteriol. 1995, 45, 139–144. [Google Scholar] [CrossRef]
- Essam, H.M.; Abdellrazeq, G.S.; Tayel, S.I.; Torky, H.A.; Fadel, A.H. Pathogenesis of Photobacterium damselae subspecies infections in sea bass and sea bream. Microb. Pathog. 2016, 99, 41–50. [Google Scholar] [CrossRef]
- Osorio, C.R.; Vences, A.; Matanza, X.M.; Terceti, M.S. Photobacterium damselae subsp. damselae, a generalist pathogen with unique virulence factors and high genetic diversity. J. Bacteriol. 2018, 200, e00002-18. [Google Scholar] [CrossRef] [PubMed]
- Romalde, J.L. Photobacterium damselae subsp. piscicida: An integrated view of a bacterial fish pathogen. Int. Microbiol. 2002, 5, 3–9. [Google Scholar] [CrossRef]
- Ho, L.P.; Han-You Lin, J.; Liu, H.C.; Chen, H.E.; Chen, T.Y.; Yang, H.L. Identification of antigens for the development of a subunit vaccine against Photobacterium damselae ssp. piscicida. Fish Shellfish Immunol. 2011, 30, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Feng, J.; Su, Y.; Ye, L.; Wang, J. Studies on the isolation of Photobacterium damselae subsp. piscicida from diseased golden pompano (Trachinotus ovatus Linnaeus) and antibacterial agents sensitivity. Vet. Microbiol. 2013, 162, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Matanza, X.M.; Osorio, C.R. Transcriptome changes in response to temperature in the fish pathogen Photobacterium damselae subsp. damselae: Clues to understand the emergence of disease outbreaks at increased seawater temperatures. PLoS ONE 2018, 13, e0210118. [Google Scholar] [CrossRef]
- Wang, F.I.; Chen, J.C. The immune response of tiger shrimp Penaeus monodon and its susceptibility to Photobacterium damselae subsp. damselae under temperature stress. Aquaculture 2006, 258, 34–41. [Google Scholar] [CrossRef]
- Zhang, Z.; Yu, Y.-X.; Wang, K.; Wang, Y.-G.; Jiang, Y.; Liao, M.-J.; Rong, X.-J. First report of skin ulceration caused by Photobacterium damselae subsp. damselae in net-cage cultured black rockfish (Sebastes schlegeli). Aquaculture 2019, 503, 1–7. [Google Scholar] [CrossRef]
- Yuen, K.Y.; Ma, L.; Wong, S.S.Y.; Ng, W.F. Fatal necrotizing fasciitis due to Vibrio damsela. Scand. J. Infect. Dis. 1993, 25, 659–661. [Google Scholar] [CrossRef]
- Lam, L.C.; Chiu, H.F.; Hung, S.F. Vitamin E in the treatment of tardive dyskinesia: A replication study. J. Nerv. Ment. Dis. 1994, 182, 113–114. [Google Scholar] [CrossRef]
- Barber, G.R.; Swygert, J.S. Necrotizing Fasciitis Due to Photobacterium damsela in a Man Lashed by a Stingray. N. Engl. J. Med. 2000, 342, 824. [Google Scholar] [CrossRef]
- Goodell, K.H.; Jordan, M.R.; Graham, R.; Cassidy, C.; Nasraway, S.A. Rapidly advancing necrotizing fasciitis caused by Photobacterium (Vibrio) damsela: A hyperaggressive variant. Crit. Care Med. 2004, 32, 278–281. [Google Scholar] [CrossRef]
- Alvarez, J.R.; Lamba, S.; Dyer, K.Y.; Apuzzio, J.J. An unusual case of urinary tract infection in a pregnant woman with Photobacterium damsela. Infect. Dis. Obstet. Gynecol. 2006, 2006, 080682. [Google Scholar] [CrossRef]
- Akram, A.; Stevens, R.P.; Konecny, P. Photobacterium damselae and Vibrio harveyi hand infection from marine exposure. Med. J. Aust. 2015, 203, 224–225.e1. [Google Scholar] [CrossRef] [PubMed]
- Yamane, K.; Asato, J.; Kawade, N.; Takahashi, H.; Kimura, B.; Arakawa, Y. Two Cases of Fatal Necrotizing Fasciitis Caused by Photobacterium damsela in Japan. J. Clin. Microbiol. 2004, 42, 1370–1372. [Google Scholar] [CrossRef] [PubMed]
- Lees, D. Viruses and bivalve shellfish. Int. J. Food Microbiol. 2000, 59, 81–116. [Google Scholar] [CrossRef]
- Hassard, F.; Sharp, J.H.; Taft, H.; LeVay, L.; Harris, J.P.; McDonald, J.E.; Tuson, K.; Wilson, J.; Jones, D.L.; Malham, S.K. Critical Review on the Public Health Impact of Norovirus Contamination in Shellfish and the Environment: A UK Perspective. Food Environ. Virol. 2017, 9, 123–141. [Google Scholar] [CrossRef] [PubMed]
- Woods, J.W.; Calci, K.R.; Marchant-Tambone, J.G.; Burkhardt, W. Detection and molecular characterization of norovirus from oysters implicated in outbreaks in the US. Food Microbiol. 2016, 59, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Diez-Valcarce, M.; Kokkinos, P.; Söderberg, K.; Bouwknegt, M.; Willems, K.; de Roda-Husman, A.M.; von Bonsdorff, C.H.; Bellou, M.; Hernández, M.; Maunula, L.; et al. Occurrence of Human Enteric Viruses in Commercial Mussels at Retail Level in Three European Countries. Food Environ. Virol. 2012, 4, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Loisy, F.; Atmar, R.L.; Le Saux, J.C.; Cohen, J.; Caprais, M.P.; Pommepuy, M.; Le Guyader, F.S. Use of rotavirus virus-like particles as surrogates to evaluate virus persistence in shellfish. Appl. Environ. Microbiol. 2005, 71, 6049–6053. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ho, B.S.W.; Tam, T.Y. Natural depuration of shellfish for human consumption: A note of caution. Water Res. 2000, 34, 1401–1406. [Google Scholar] [CrossRef]
- Croci, L.; Suffredini, E.; Cozzi, L.; Toti, L. Effects of depuration of molluscs experimentally contaminated with Escherichia coli, Vibrio cholerae O1 and Vibrio parahaemolyticus. J. Appl. Microbiol. 2002, 92, 460–465. [Google Scholar] [CrossRef]
- Polo, D.; Álvarez, C.; Díez, J.; Darriba, S.; Longa, Á.; Romalde, J.L. Viral elimination during commercial depuration of shellfish. Food Control. 2014, 43, 206–212. [Google Scholar] [CrossRef]
- Chironna, M.; Germinario, C.; De Medici, D.; Fiore, A.; Di Pasquale, S.; Quarto, M.; Barbuti, S. Detection of hepatitis A virus in mussels from different sources marketed in Puglia region (South Italy). Int. J. Food Microbiol. 2002, 75, 11–18. [Google Scholar] [CrossRef]
- Croci, L.; Losio, M.N.; Suffredini, E.; Pavoni, E.; Di Pasquale, S.; Fallacara, F.; Arcangeli, G. Assessment of human enteric viruses in shellfish from the northern Adriatic sea. Int. J. Food Microbiol. 2007, 114, 252–257. [Google Scholar] [CrossRef]
- Terio, V.; Martella, V.; Moschidou, P.; Di Pinto, P.; Tantillo, G.; Buonavoglia, C. Norovirus in retail shellfish. Food Microbiol. 2010, 27, 29–32. [Google Scholar] [CrossRef]
- La Bella, G.; Martella, V.; Basanisi, M.G.; Nobili, G.; Terio, V.; La Salandra, G. Food-Borne Viruses in Shellfish: Investigation on Norovirus and HAV Presence in Apulia (SE Italy). Food Environ. Virol. 2017, 9, 179–186. [Google Scholar] [CrossRef]
- Suffredini, E.; Pepe, T.; Ventrone, I.; Croci, L. Norovirus detection in shellfish using two real-time RT-PCR methods. New Microbiol. 2011, 34, 9–16. [Google Scholar]
- Henigman, U.; Biasizzo, M.; Vadnjal, S.; Toplak, I.; Gombač, M.; Steyer, A.; Prijatelj, M.P.; Ambrožič, M.; Fonda, I.; Kirbiš, A.; et al. Molecular characterisation of noroviruses detected in mussels (Mytilus galloprovincialis) from harvesting areas in Slovenia. New Microbiol. 2015, 38, 225–233. [Google Scholar] [PubMed]
- Woods, J.W.; Burkhardt, W. Occurrence of norovirus and hepatitis a virus in U.S. Oysters. Food Environ. Virol. 2010, 2, 176–182. [Google Scholar] [CrossRef]
- Torok, V.; Hodgson, K.; McLeod, C.; Tan, J.; Malhi, N.; Turnbull, A. National survey of foodborne viruses in Australian oysters at production. Food Microbiol. 2018, 69, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Polo, D.; Varela, M.F.; Romalde, J.L. Detection and quantification of hepatitis A virus and norovirus in Spanish authorized shellfish harvesting areas. Int. J. Food Microbiol. 2015, 193, 43–50. [Google Scholar] [CrossRef]
- Lowther, J.A.; Gustar, N.E.; Powell, A.L.; Hartnell, R.E.; Lees, D.N. Two-year systematic study to assess norovirus contamination in oysters from commercial harvesting areas in the United Kingdom. Appl. Environ. Microbiol. 2012, 78, 5812–5817. [Google Scholar] [CrossRef]
- Lee, R.J.; Silk, R. Sources of variation of Escherichia coli concentrations in bivalve molluscs. J. Water Health 2013, 11, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.S.; Rodrigues, M.J.; Henriques, A.R. Specific hygiene procedures and practices assessment: A cross-sectional study in fresh fishery product retailers of lisbon’s traditional food markets. Foods 2021, 10, 1805. [Google Scholar] [CrossRef]
- Cavalier-Smith, T.; Chao, E.E.Y. Phylogeny and Classification of Phylum Cercozoa (Protozoa). Protist 2003, 154, 341–358. [Google Scholar] [CrossRef] [PubMed]
- Feist, S.W.; Hine, P.M.; Bateman, K.S.; Stentiford, G.D.; Longshaw, M. Paramarteilia canceri sp. n. (Cercozoa) in the European edible crab (Cancer pagurus) with a proposal for the revision of the order Paramyxida Chatton, 1911. Folia Parasitol. 2009, 56, 73–85. [Google Scholar] [CrossRef]
- Arzul, I.; Chollet, B.; Boyer, S.; Bonnet, D.; Gaillard, J.; Baldi, Y.; Robert, M.; Joly, J.Â.P.; Garcia, C.; Bouchoucha, M. Contribution to the understanding of the cycle of the protozoan parasite Marteilia refringens. Parasitology 2014, 141, 227–240. [Google Scholar] [CrossRef]
- Carrasco, N.; Green, T.; Itoh, N. Marteilia spp. parasites in bivalves: A revision of recent studies. J. Invertebr. Pathol. 2015, 131, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Zgouridou, A.; Tripidaki, E.; Giantsis, I.A.; Theodorou, J.A.; Kalaitzidou, M.; Raitsos, D.E.; Lattos, A.; Mavropoulou, A.M.; Sofianos, S.; Karagiannis, D.; et al. The current situation and potential effects of climate change on the microbial load of marine bivalves of the Greek coastlines: An integrative review. Environ. Microbiol. 2021, in press. [Google Scholar] [CrossRef] [PubMed]



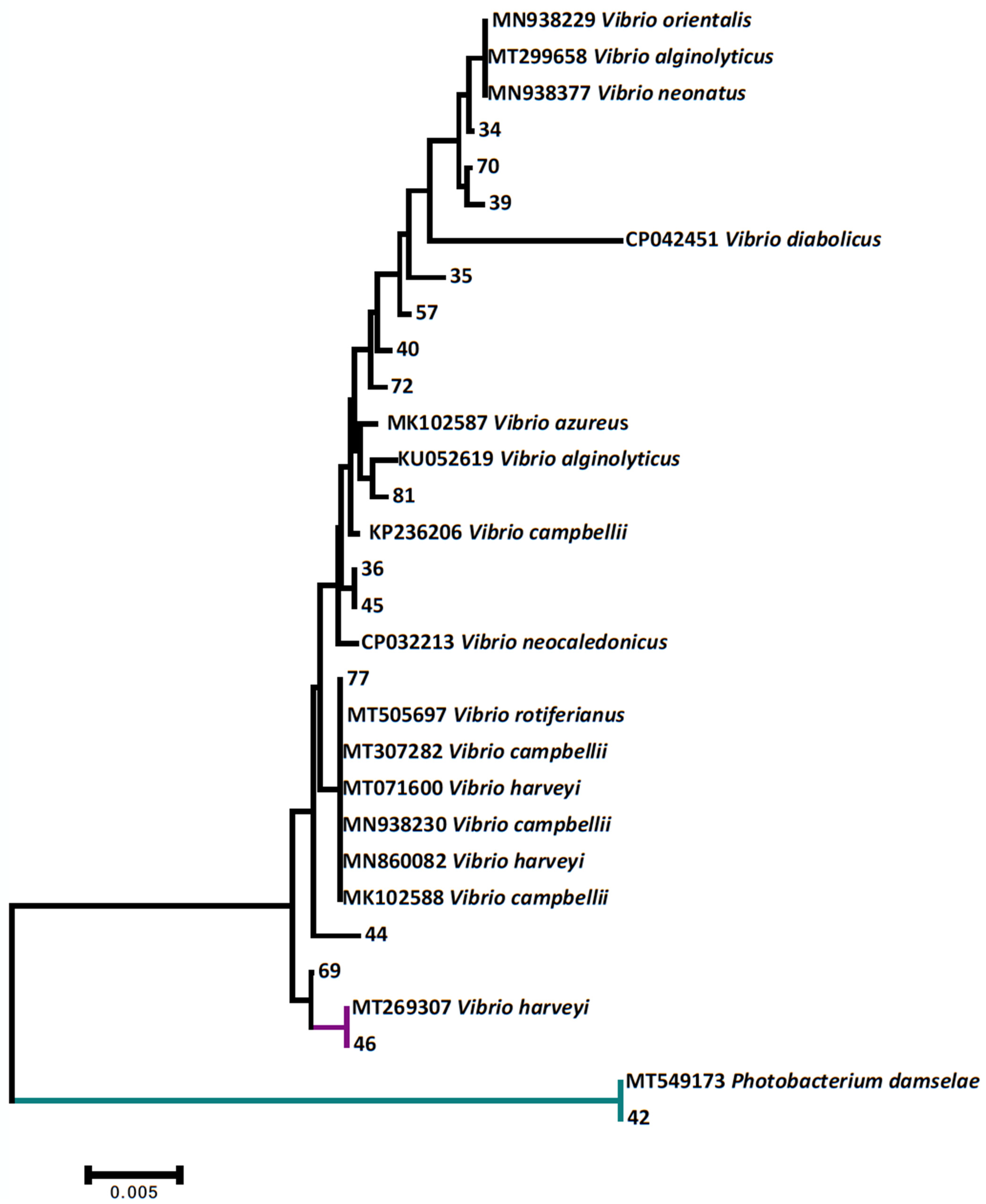{kind=link}
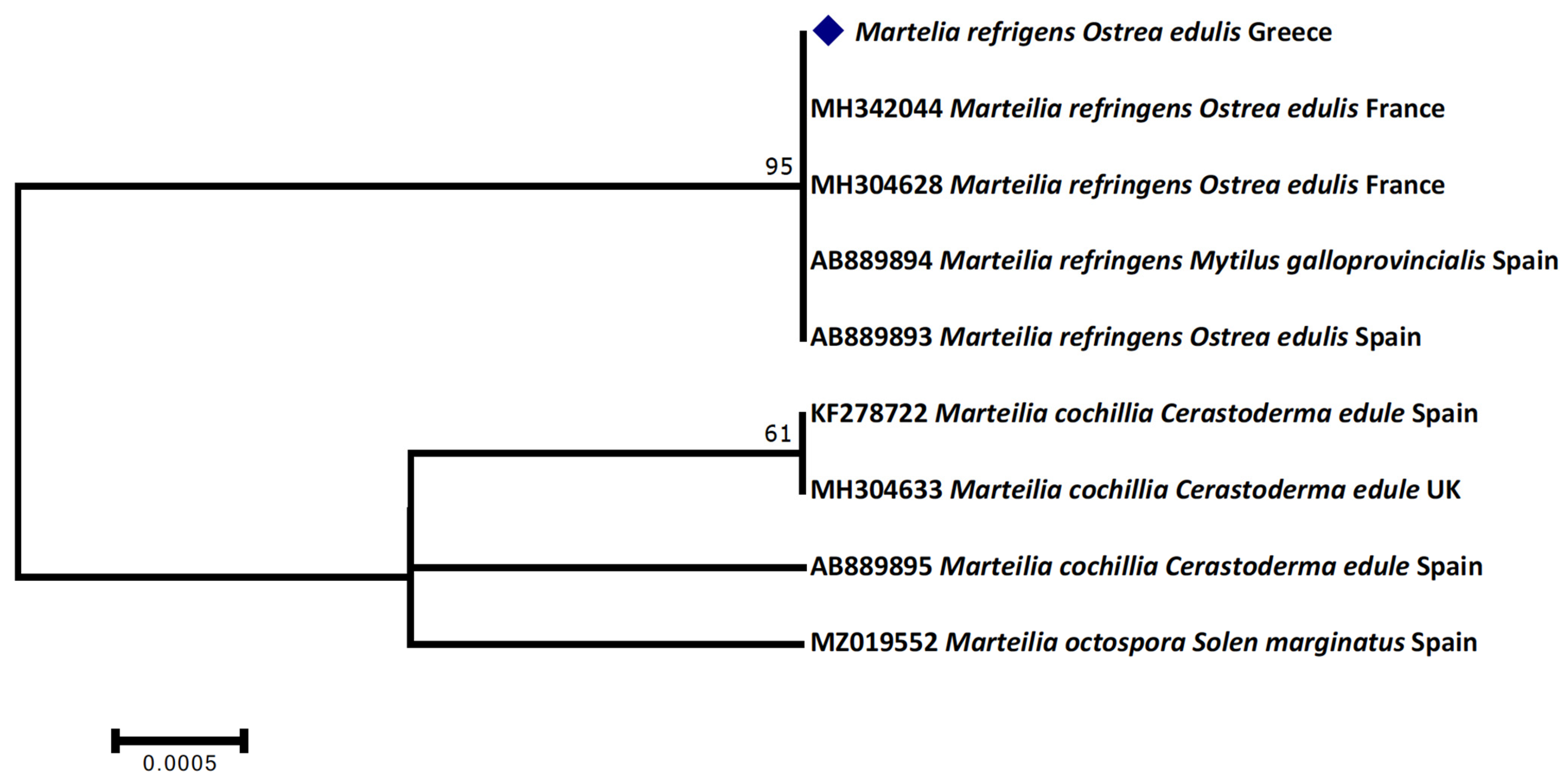{kind=link}
| ID No | Species | Market | Date | ID No | Species | Market | Date |
|---|---|---|---|---|---|---|---|
| 14 | Callista chione | Nea Michaniona | 3 March 2020 | 42 | Ostrea edulis | Kapani | 26 June 2020 |
| 16 | Callista chione | Kapani | 9 April 2020 | 56 | Ostrea edulis | Kapani | 7 August 2020 |
| 43 | Callista chione | Nea Michaniona | 22 July 2020 | 68 | Ostrea edulis | Nea Michaniona | 1 September 2020 |
| 54 | Callista chione | Kapani | 26 June 2020 | 70 | Ostrea edulis | Nea Michaniona | 22 July 2020 |
| 58 | Callista chione | Nea Michaniona | 10 February 2020 | 77 | Ostrea edulis | Nea Michaniona | 4 August 2020 |
| 59 | Callista chione | Kapani | 7 August 2020 | 88 | Ostrea edulis | Kapani | 2 November 2020 |
| 60 | Callista chione | Nea Michaniona | 1 September 2020 | 89 | Ostrea edulis | Nea Michaniona | 10 November 2020 |
| 64 | Callista chione | Kapani | 4 September 2020 | 116 | Ostrea edulis | Kapani | 12 January 2021 |
| 69 | Callista chione | Kapani | 9 July 2020 | 200 | Ostrea edulis | Nea Michaniona | 10 December 2020 |
| 74 | Callista chione | Nea Michaniona | 15 November 2020 | 10 | Ruditapes decussatus | Kapani | 28 February 2020 |
| 78 | Callista chione | Kapani | 5 October 2020 | 39 | Ruditapes decussatus | Kapani | 3 June 2020 |
| 79 | Callista chione | Nea Michaniona | 4 October 2020 | 57 | Ruditapes decussatus | Nea Michaniona | 4 August 2020 |
| 95 | Callista chione | Nea Michaniona | 10 November 2020 | 87 | Ruditapes decussatus | Kapani | 5 October 2020 |
| 97 | Callista chione | Kapani | 12 November 2020 | 98 | Ruditapes decussatus | Nea Michaniona | 10 November 2020 |
| 104 | Callista chione | Nea Michaniona | 4 August 2020 | 111 | Ruditapes decussatus | Kapani | 2 December 2020 |
| 113 | Callista chione | Kapani | 2 December 2020 | 112 | Ruditapes decussatus | Nea Michaniona | 10 December 2020 |
| 114 | Callista chione | Nea Michaniona | 10 December 2020 | 1 | Venus verrucosa | Nea Michaniona | 20 January 2020 |
| 3 | Mytilus galloprovincialis | Nea Michaniona | 20 January 2020 | 2 | Venus verrucosa | Kapani | 28 January 2020 |
| 4 | Mytilus galloprovincialis | Kapani | 28 January 2020 | 12 | Venus verrucosa | Nea Michaniona | 3 March 2020 |
| 17 | Mytilus galloprovincialis | Nea Michaniona | 16 April 2020 | 13 | Venus verrucosa | Kapani | 19 March 2020 |
| 18 | Mytilus galloprovincialis | Kapani | 21 April 2020 | 19 | Venus verrucosa | Kapani | 9 April 2020 |
| 26 | Mytilus galloprovincialis | Nea Michaniona | 26 May 2020 | 20 | Venus verrucosa | Nea Michaniona | 10 February 2020 |
| 33 | Mytilus galloprovincialis | Kapani | 22 May 2020 | 24 | Venus verrucosa | Nea Michaniona | 16 April 2020 |
| 34 | Mytilus galloprovincialis | Nea Michaniona | 26 May 2020 | 25 | Venus verrucosa | Kapani | 21 February 2020 |
| 45 | Mytilus galloprovincialis | Kapani | 3 June 2020 | 30 | Venus verrucosa | Kapani | 12 May 2020 |
| 46 | Mytilus galloprovincialis | Nea Michaniona | 23 June 2020 | 35 | Venus verrucosa | Nea Michaniona | 26 May 2020 |
| 48 | Mytilus galloprovincialis | Kapani | 26 June 2020 | 36 | Venus verrucosa | Kapani | 10 June 2020 |
| 49 | Mytilus galloprovincialis | Nea Michaniona | 16 June 2020 | 38 | Venus verrucosa | Nea Michaniona | 30 October 2020 |
| 52 | Mytilus galloprovincialis | Kapani | 15 July 2020 | 40 | Venus verrucosa | Kapani | 3 June 2020 |
| 61 | Mytilus galloprovincialis | Nea Michaniona | 1 September 2020 | 44 | Venus verrucosa | Nea Michaniona | 22 July 2020 |
| 62 | Mytilus galloprovincialis | Kapani | 4 September 2020 | 50 | Venus verrucosa | Kapani | 23 July 2020 |
| 71 | Mytilus galloprovincialis | Nea Michaniona | 2 October 2020 | 51 | Venus verrucosa | Nea Michaniona | 23 June 2020 |
| 72 | Mytilus galloprovincialis | Kapani | 2 October 2020 | 53 | Venus verrucosa | Kapani | 7 August 2020 |
| 76 | Mytilus galloprovincialis | Nea Michaniona | 30 October 2020 | 55 | Venus verrucosa | Nea Michaniona | 4 August 2020 |
| 81 | Mytilus galloprovincialis | Nea Michaniona | 22 July 2020 | 82 | Venus verrucosa | Kapani | 5 October 2020 |
| 83 | Mytilus galloprovincialis | Kapani | 7 August 2020 | 94 | Venus verrucosa | Kapani | 2 November 2020 |
| 84 | Mytilus galloprovincialis | Nea Michaniona | 4 August 2020 | 96 | Venus verrucosa | Nea Michaniona | 10 November 2020 |
| 85 | Mytilus galloprovincialis | Kapani | 15 November 2020 | 99 | Venus verrucosa | Nea Michaniona | 1 September 2020 |
| 90 | Mytilus galloprovincialis | Kapani | 16 December 2020 | 100 | Venus verrucosa | Kapani | 10 September 2020 |
| 91 | Mytilus galloprovincialis | Nea Michaniona | 10 December 2020 | 101 | Venus verrucosa | Nea Michaniona | 25 August 2020 |
| 92 | Mytilus galloprovincialis | Kapani | 2 February 2020 | 103 | Venus verrucosa | Kapani | 12 December 2020 |
| 102 | Mytilus galloprovincialis | Nea Michaniona | 10 February 2020 | 108 | Venus verrucosa | Kapani | 2 December 2020 |
| 107 | Mytilus galloprovincialis | Nea Michaniona | 9 March 2021 | 109 | Venus verrucosa | Nea Michaniona | 10 December 2020 |
| 119 | Venus verrucosa | Kapani | 12 January 2021 |
| ID No | E. coli | Salmonella spp. | Vibrio sp. | Marteilia refringens | ID No | E. coli | Salmonella spp. | Vibrio sp. | Marteilia refringens |
|---|---|---|---|---|---|---|---|---|---|
| 1 | <18 | N.D. | − | − | 64 | <18 | N.D. | + | − |
| 2 | <18 | N.D. | − | − | 68 | <18 | N.D. | − | − |
| 3 | <18 | N.D. | − | − | 69 | <18 | N.D. | + | − |
| 4 | <18 | N.D. | − | − | 70 | 20 | N.D. | + | − |
| 10 | 20 | N.D. | − | − | 71 | <18 | N.D. | + | − |
| 12 | <18 | N.D. | − | − | 72 | 20 | N.D. | + | − |
| 13 | <18 | N.D. | − | − | 74 | <18 | N.D. | + | − |
| 14 | 20 | N.D. | − | − | 76 | 20 | N.D. | + | − |
| 16 | <18 | N.D. | − | − | 77 | <18 | N.D. | + | − |
| 17 | 20 | N.D. | − | − | 78 | <18 | N.D. | + | − |
| 18 | <18 | N.D. | − | − | 79 | 45 | N.D. | + | − |
| 19 | <18 | N.D. | − | − | 81 | <18 | N.D. | + | − |
| 20 | 92 | N.D. | − | − | 82 | <18 | N.D. | + | − |
| 24 | <18 | N.D. | − | − | 83 | <18 | N.D. | − | − |
| 25 | <18 | N.D. | − | − | 84 | <18 | N.D. | − | − |
| 26 | <18 | N.D. | − | − | 85 | 40 | N.D. | − | − |
| 30 | 45 | N.D. | − | − | 87 | <18 | N.D. | + | − |
| 33 | 20 | N.D. | + | − | 88 | <18 | N.D. | + | − |
| 34 | <18 | N.D. | + | − | 89 | <18 | N.D. | + | + |
| 35 | <18 | N.D. | + | − | 90 | <18 | N.D. | + | − |
| 36 | <18 | N.D. | + | − | 91 | <18 | N.D. | − | − |
| 38 | <18 | N.D. | + | − | 92 | <18 | N.D. | − | − |
| 39 | <18 | N.D. | + | − | 94 | <18 | N.D. | − | − |
| 40 | <18 | N.D. | + | − | 95 | <18 | N.D. | + | − |
| 42 | <18 | N.D. | − | − | 96 | <18 | N.D. | − | − |
| 43 | 20 | N.D. | + | − | 97 | <18 | N.D. | − | − |
| 44 | <18 | N.D. | + | − | 98 | <18 | N.D. | + | − |
| 45 | 230 | N.D. | + | − | 99 | <18 | N.D. | − | − |
| 46 | 20 | N.D. | + | − | 100 | <18 | N.D. | − | − |
| 48 | <18 | N.D. | − | − | 101 | <18 | N.D. | + | − |
| 49 | 20 | N.D. | + | − | 102 | 20 | N.D. | − | − |
| 50 | <18 | N.D. | + | − | 103 | <18 | N.D. | − | − |
| 51 | 20 | N.D. | + | − | 104 | <18 | N.D. | − | − |
| 52 | <18 | N.D. | + | − | 107 | <18 | N.D. | − | − |
| 53 | <18 | N.D. | + | − | 108 | 130 | N.D. | − | − |
| 54 | <18 | N.D. | + | − | 109 | 45 | N.D. | − | − |
| 55 | <18 | N.D. | + | − | 111 | <18 | N.D. | − | − |
| 56 | 20 | N.D. | + | − | 112 | 20 | N.D. | − | − |
| 57 | <18 | N.D. | + | − | 113 | <18 | N.D. | − | − |
| 58 | <18 | N.D. | + | − | 114 | <18 | N.D. | − | − |
| 59 | <18 | N.D. | + | − | 116 | <18 | N.D. | − | + |
| 60 | <18 | N.D. | + | − | 119 | <18 | N.D. | − | − |
| 61 | <18 | N.D. | + | − | 200 | <18 | N.D. | − | + |
| 62 | 2400 | N.D. | + | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lattos, A.; Chaligiannis, I.; Papadopoulos, D.; Giantsis, I.A.; Petridou, E.I.; Vafeas, G.; Staikou, A.; Michaelidis, B. How Safe to Eat Are Raw Bivalves? Host Pathogenic and Public Health Concern Microbes within Mussels, Oysters, and Clams in Greek Markets. Foods 2021, 10, 2793. https://doi.org/10.3390/foods10112793
Lattos A, Chaligiannis I, Papadopoulos D, Giantsis IA, Petridou EI, Vafeas G, Staikou A, Michaelidis B. How Safe to Eat Are Raw Bivalves? Host Pathogenic and Public Health Concern Microbes within Mussels, Oysters, and Clams in Greek Markets. Foods. 2021; 10(11):2793. https://doi.org/10.3390/foods10112793
Chicago/Turabian StyleLattos, Athanasios, Ilias Chaligiannis, Dimitrios Papadopoulos, Ioannis A. Giantsis, Evanthia I. Petridou, George Vafeas, Alexandra Staikou, and Basile Michaelidis. 2021. "How Safe to Eat Are Raw Bivalves? Host Pathogenic and Public Health Concern Microbes within Mussels, Oysters, and Clams in Greek Markets" Foods 10, no. 11: 2793. https://doi.org/10.3390/foods10112793
APA StyleLattos, A., Chaligiannis, I., Papadopoulos, D., Giantsis, I. A., Petridou, E. I., Vafeas, G., Staikou, A., & Michaelidis, B. (2021). How Safe to Eat Are Raw Bivalves? Host Pathogenic and Public Health Concern Microbes within Mussels, Oysters, and Clams in Greek Markets. Foods, 10(11), 2793. https://doi.org/10.3390/foods10112793