
Mineral Concentrations in Bovine Milk from Farms with Contrasting Grazing Management

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Design of the Survey and Experiment
2.2. Analysis of Milk Samples for Basic Composition and Minerals
2.3. Statistical Analysis
3. Results
3.1. Breeding and Feeding in the Herds
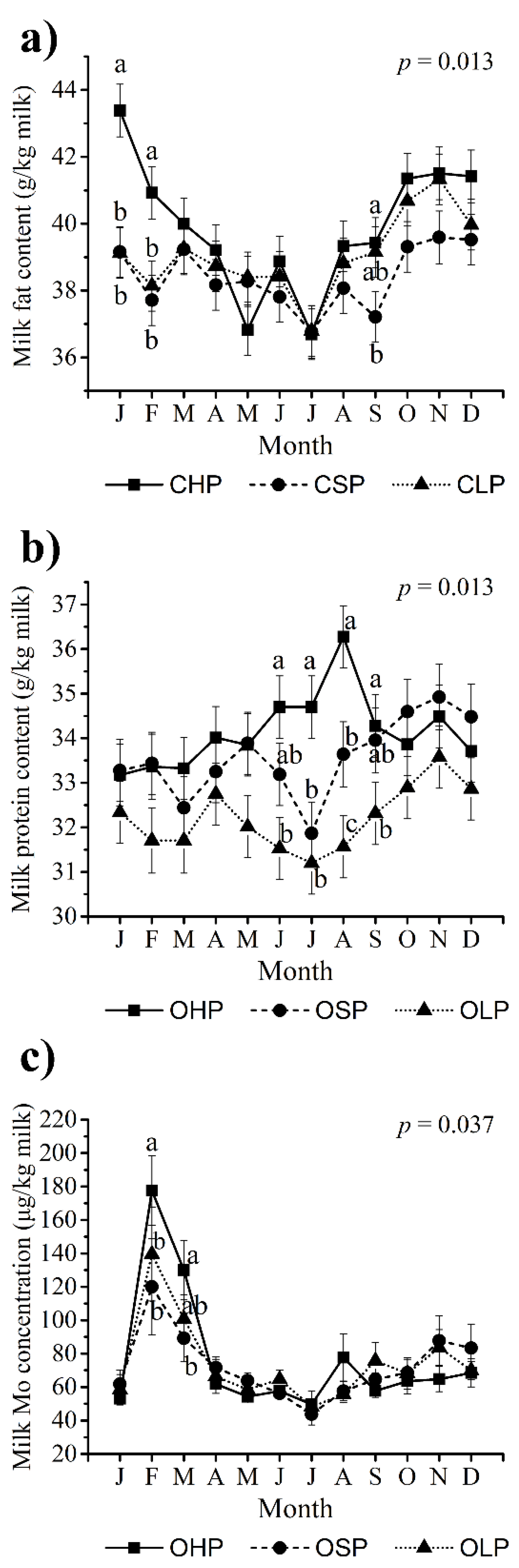
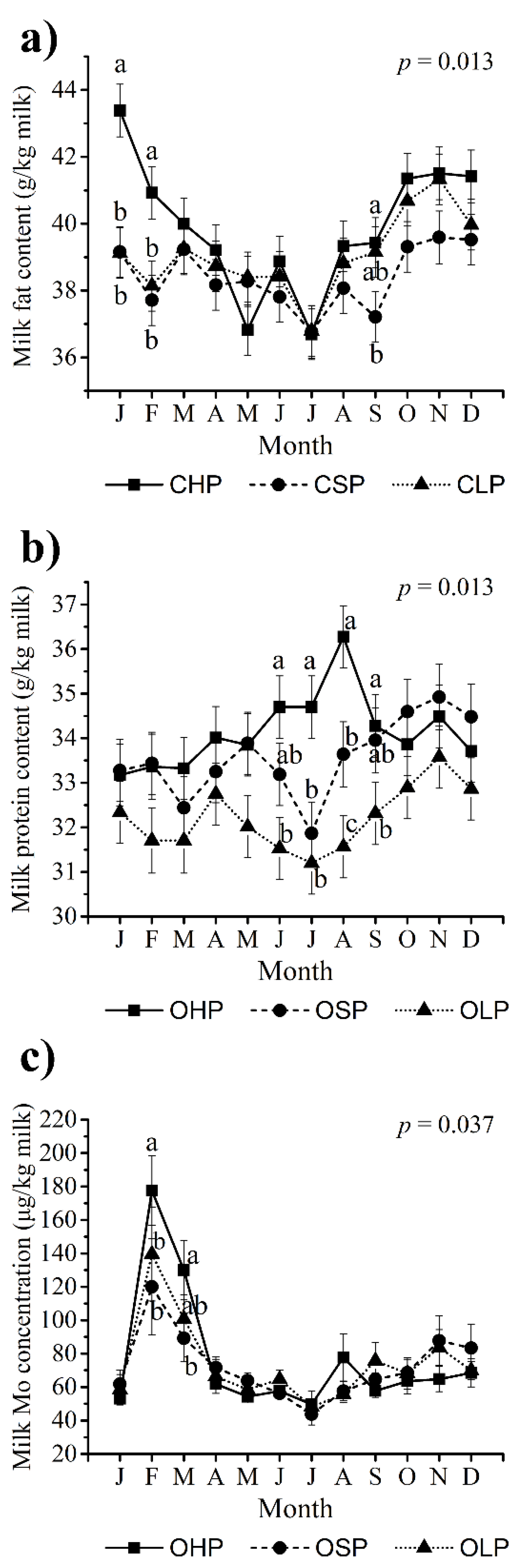
3.2. Milk Yield and Basic Composition
3.3. Milk Mineral Profile
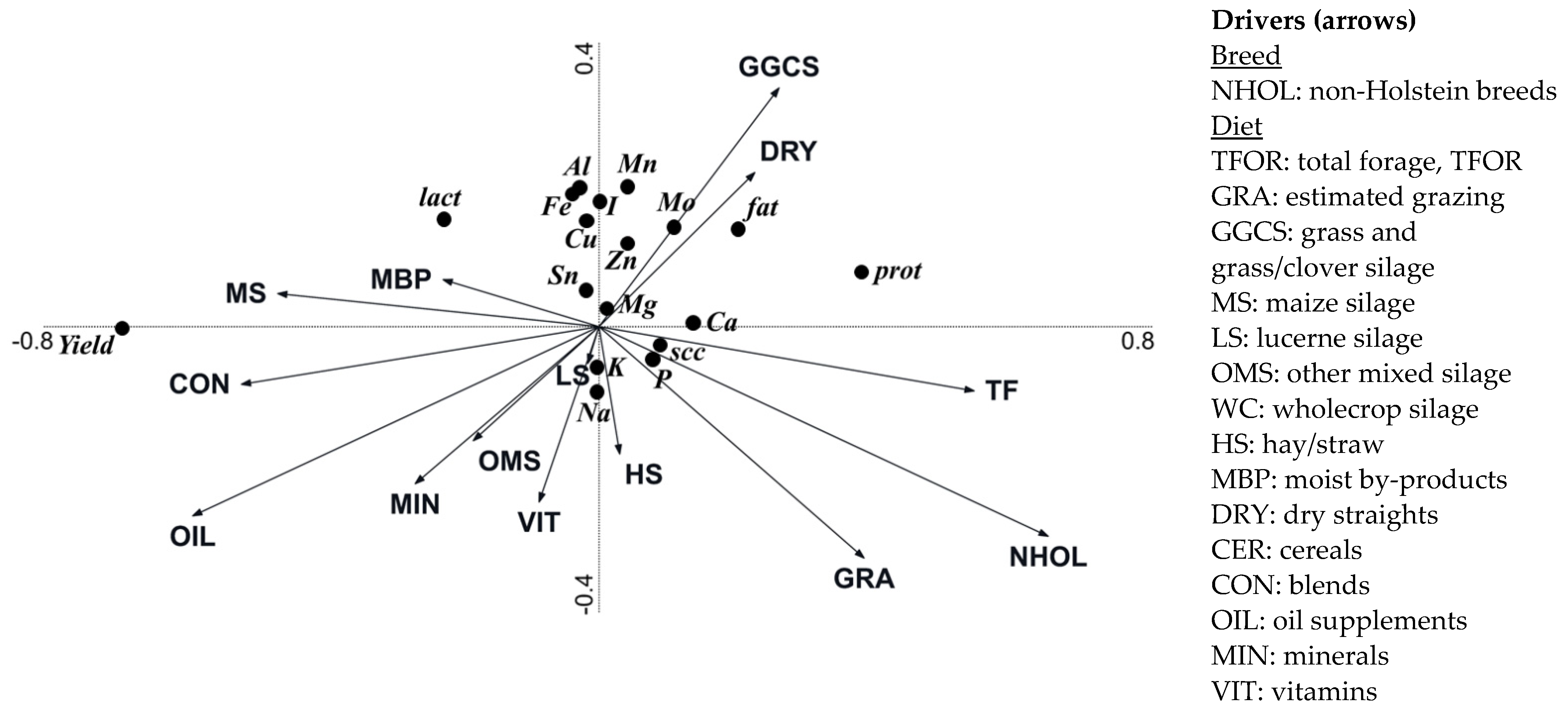
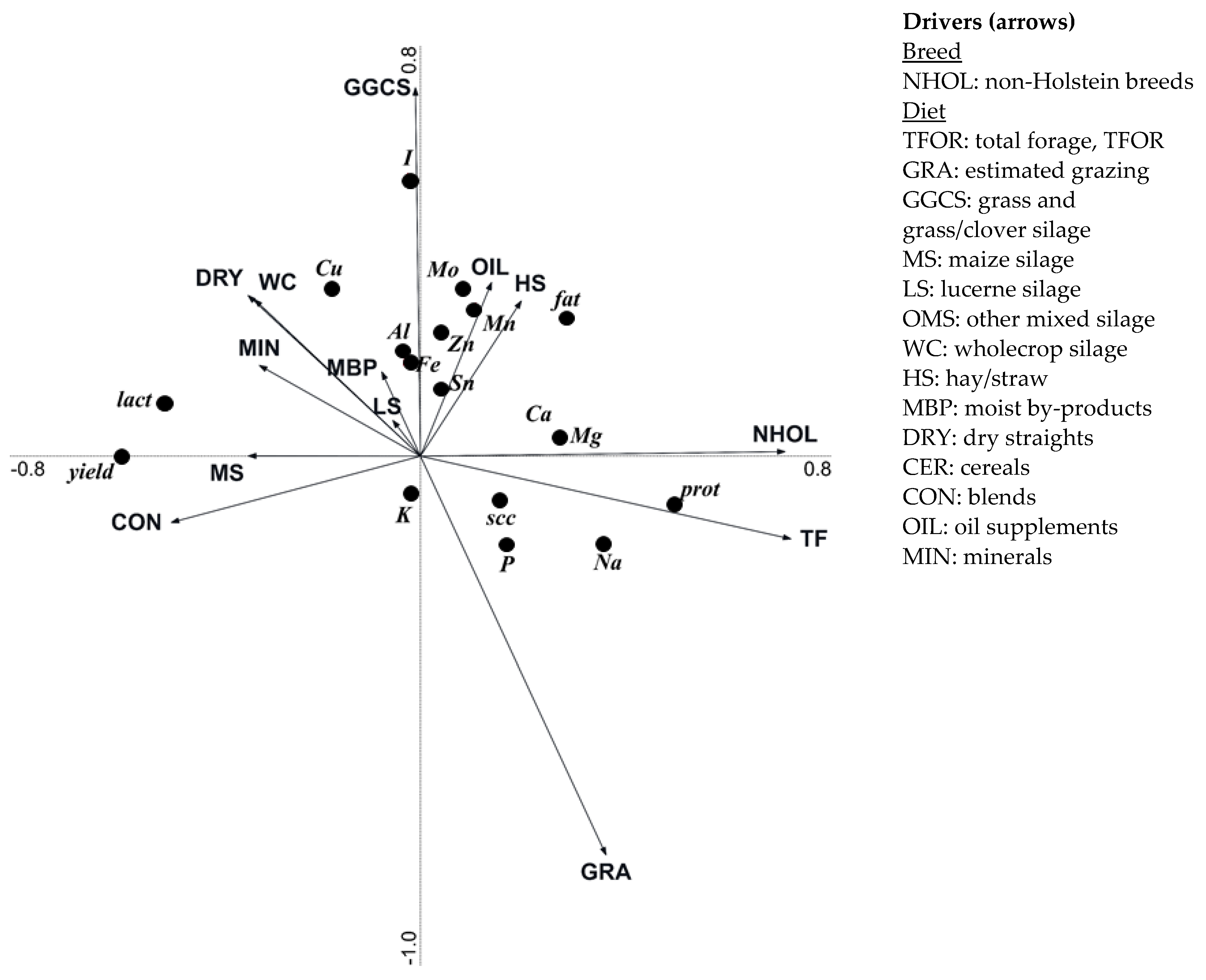
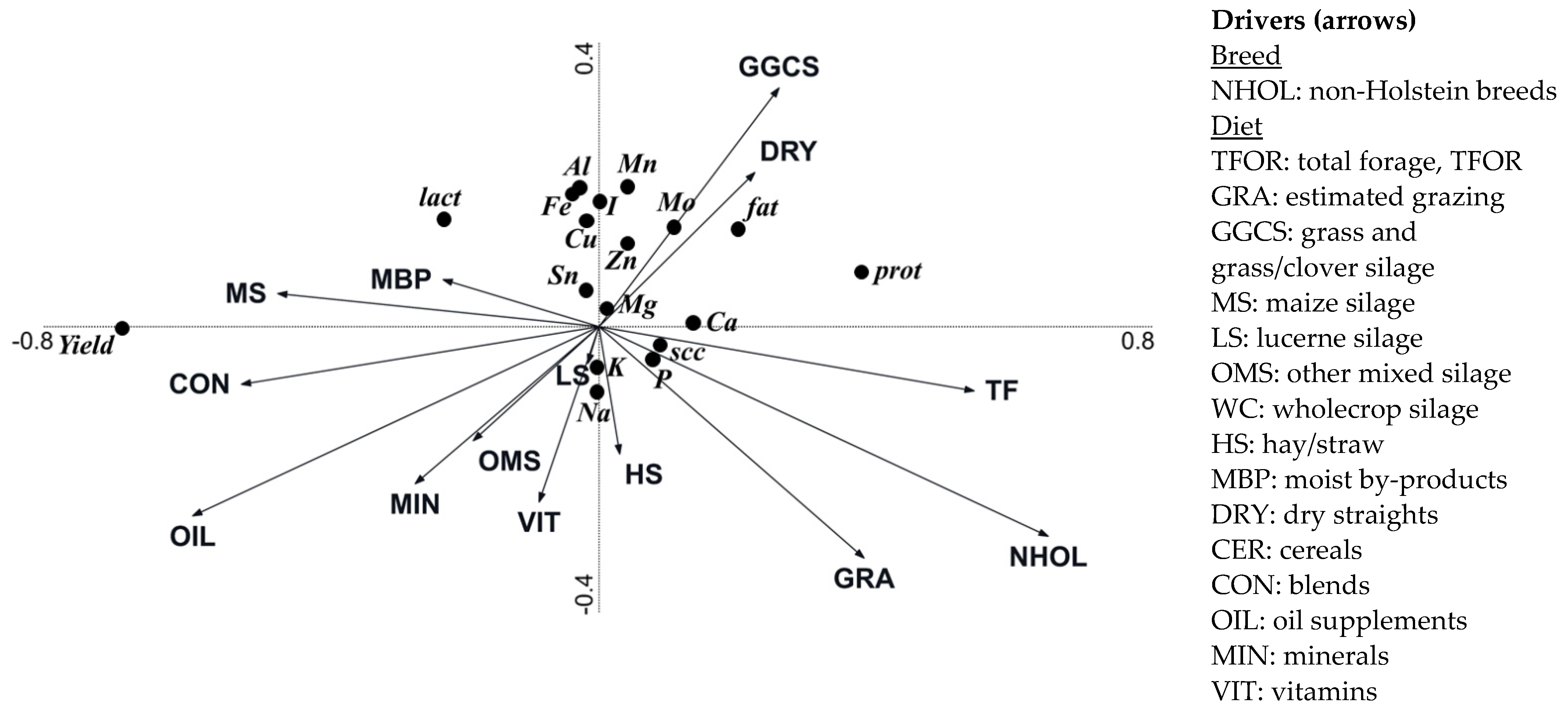
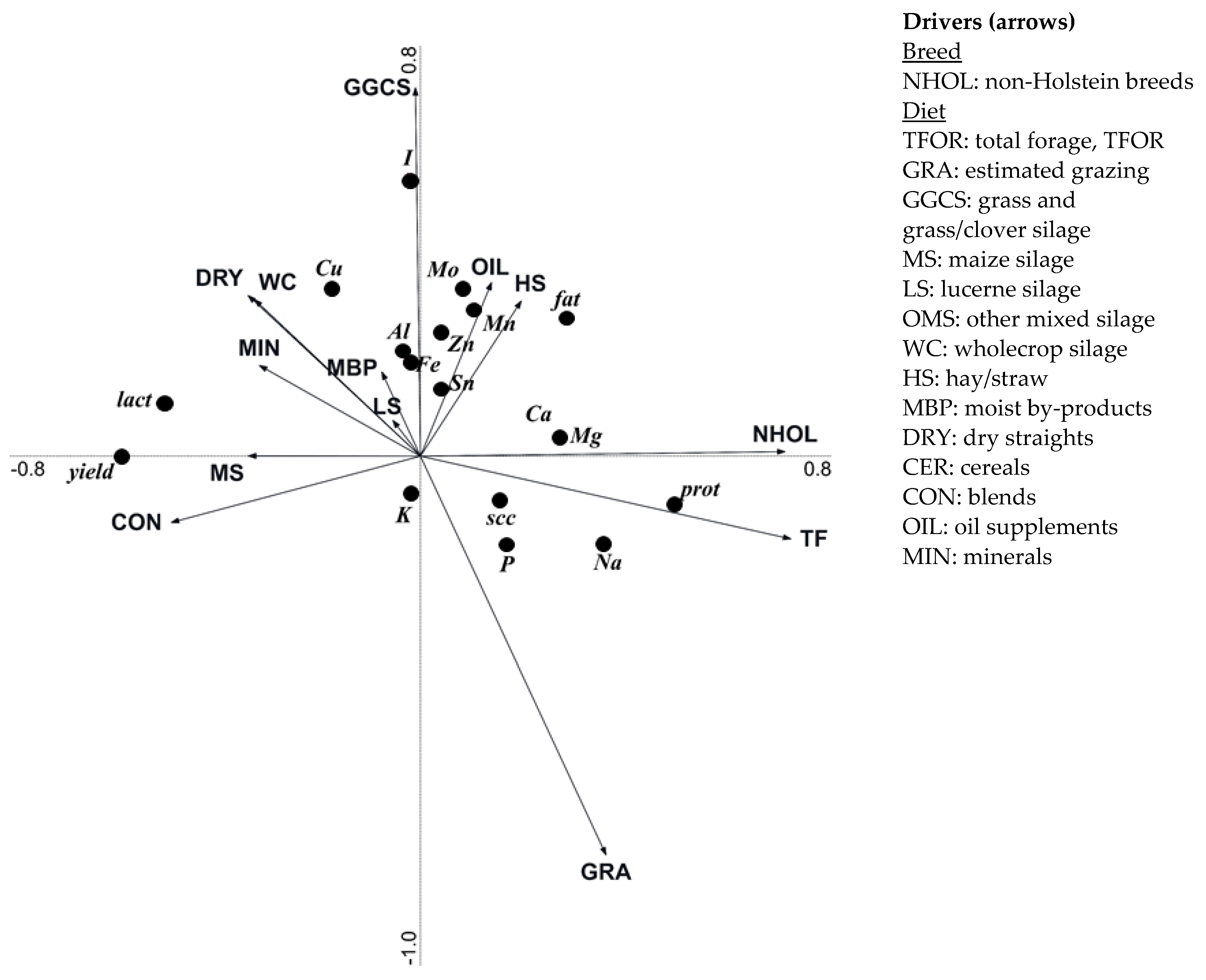
3.4. Effect of Breeding and Diet Parameters
4. Discussion
4.1. Milk Yield and Basic Composition
4.2. Milk Mineral Profile
4.3. Potential Impact of Grazing Management on the Mineral Intakes of UK Consumers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flynn, A. Minerals and trace elements in milk. Adv. Food Nutr. Res. 1992, 36, 209–252. [Google Scholar] [CrossRef]
- Haug, A.; Høstmark, A.T.; Harstad, O.M. Bovine milk in human nutrition—A review. Lipids Health Dis. 2007, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Thorning, T.K.; Raben, A.; Tholstrup, T.; Soedamah-Muthu, S.S.; Givens, I.; Astrup, A. Milk and dairy products: Good or bad for human health? An assessment of the totality of scientific evidence. Food Nutr. Res. 2016, 60, 32527. [Google Scholar] [CrossRef] [Green Version]
- Roberts, C.; Steer, T.; Maplethorpe, N.; Cox, L.; Meadows, S.; Nicholson, S.; Page, P.; Swan, G. National Diet and Nutrition Survey. Results from Years 7–8 (Combined) of the Rolling Programme (2014/15 to 2015/16): A Survey Carried out on Behalf of Public Health England and the Food Standards Agency. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/699241/NDNS_results_years_7_and_8.pdf/ (accessed on 10 August 2021).
- Amenu, B.; Deeth, H. The impact of milk composition on cheddar cheese manufacture. Aust. J. Dairy Technol. 2007, 62, 171–184. [Google Scholar]
- Stocco, G.; Summer, A.; Cipolat-Gotet, C.; Malacarne, M.; Cecchinato, A.; Amalfitano, N.; Bittante, G. The mineral profile affects the coagulation pattern and cheese-making efficiency of bovine milk. J. Dairy Sci. 2021, 104, 8439–8453. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, D.; Delaby, L.; van den Pol-van Dasselaar, A.; Shalloo, L. Increasing Grazing in Dairy Cow Milk Production Systems in Europe. Sustainability 2020, 12, 2443. [Google Scholar] [CrossRef] [Green Version]
- Cabiddu, A.; Delgadillo-Puga, C.; Decandia, M.; Molle, G. Extensive Ruminant Production Systems and Milk Quality with Emphasis on Unsaturated Fatty Acids, Volatile Compounds, Antioxidant Protection Degree and Phenol Content. Animals 2019, 9, 771. [Google Scholar] [CrossRef] [Green Version]
- Elgersma, A. Grazing increases the unsaturated fatty acid concentration of milk from grass-fed cows: A review of the contributing factors, challenges and future perspectives. Eur. J. Lipid Sci. Technol. 2015, 117, 1345–1369. [Google Scholar] [CrossRef]
- Kay, J.K.; Roche, J.R.; Kolver, E.S.; Thomson, N.A.; Baumgard, L.H. A comparison between feeding systems (pasture and TMR) and the effect of vitamin E supplementation on plasma and milk fatty acid profiles in dairy cows. J. Dairy Res. 2005, 72, 322–332. [Google Scholar] [CrossRef]
- Orjales, I.; Herrero-Latorre, C.; Miranda, M.; Rey-Crespo, F.; Rodriguez-Bermudez, R.; Lopez-Alonso, M. Evaluation of trace element status of organic dairy cattle. Animals 2018, 12, 1296–1305. [Google Scholar] [CrossRef]
- Stergiadis, S.; Leifert, C.; Seal, C.J.; Eyre, M.D.; Nielsen, J.H.; Larsen, M.K.; Slots, T.; Steinshamn, H.; Butler, G. Effect of Feeding Intensity and Milking System on Nutritionally Relevant Milk Components in Dairy Farming Systems in the North East of England. J. Agric. Food Chem. 2012, 60, 7270–7281. [Google Scholar] [CrossRef]
- Stergiadis, S.; Leifert, C.; Seal, C.J.; Eyre, M.D.; Larsen, M.K.; Slots, T.; Nielsen, J.H.; Butler, G. A 2-year study on milk quality from three pasture-based dairy systems of contrasting production intensities in Wales. J. Agric. Sci. 2015, 153, 708–731. [Google Scholar] [CrossRef] [Green Version]
- Butler, G.; Stergiadis, S.; Seal, C.; Eyre, M.; Leifert, C. Fat composition of organic and conventional retail milk in northeast England. J. Dairy Sci. 2011, 94, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Stergiadis, S.; Berlitz, C.B.; Hunt, B.; Garg, S.; Ian Givens, D.; Kliem, K.E. An update to the fatty acid profiles of bovine retail milk in the United Kingdom: Implications for nutrition in different age and gender groups. Food Chem. 2019, 276, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Kliem, K.E.; Shingfield, K.J.; Livingstone, K.M.; Givens, D.I. Seasonal variation in the fatty acid composition of milk available at retail in the United Kingdom and implications for dietary intake. Food Chem. 2013, 141, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Średnicka-Tober, D.; Barański, M.; Seal, C.J.; Sanderson, R.; Benbrook, C.; Steinshamn, H.; Gromadzka-Ostrowska, J.; Rembiałkowska, E.; Skwarło-Sońta, K.; Eyre, M.; et al. Higher PUFA and n-3 PUFA, conjugated linoleic acid, α-tocopherol and iron, but lower iodine and selenium concentrations in organic milk: A systematic literature review and meta- and redundancy analyses. Br. J. Nutr. 2016, 115, 1043–1060. [Google Scholar] [CrossRef] [Green Version]
- Nozière, P.; Graulet, B.; Lucas, A.; Martin, B.; Grolier, P.; Doreau, M. Carotenoids for ruminants: From forages to dairy products. Anim. Feed. Sci. Technol. 2006, 131, 418–450. [Google Scholar] [CrossRef]
- Butler, G.; Nielsen, J.H.; Slots, T.; Seal, C.; Eyre, M.D.; Sanderson, R.; Leifert, C. Fatty acid and fat-soluble antioxidant concentrations in milk from high- and low-input conventional and organic systems: Seasonal variation. J. Sci. Food Agric. 2008, 88, 1431–1441. [Google Scholar] [CrossRef]
- Qin, N.; Faludi, G.; Beauclercq, S.; Pitt, J.; Desnica, N.; Pétursdóttir, Á.; Newton, E.E.; Angelidis, A.; Givens, I.; Juniper, D.; et al. Macromineral and trace element concentrations and their seasonal variation in milk from organic and conventional dairy herds. Food Chem. 2021, 359, 129865. [Google Scholar] [CrossRef]
- Stevenson, M.C.; Drake, C.; Givens, D.I. Further studies on the iodine concentration of conventional, organic and UHT semi-skimmed milk at retail in the UK. Food Chem. 2018, 239, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Payling, L.M.; Juniper, D.T.; Drake, C.; Rymer, C.; Givens, D.I. Effect of milk type and processing on iodine concentration of organic and conventional winter milk at retail: Implications for nutrition. Food Chem. 2015, 178, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Flachowsky, G.; Franke, K.; Meyer, U.; Leiterer, M.; Schöne, F. Influencing factors on iodine content of cow milk. Eur. J. Nutr. 2014, 53, 351–365. [Google Scholar] [CrossRef]
- Stergiadis, S.; Bieber, A.; Chatzidimitriou, E.; Franceschin, E.; Isensee, A.; Rempelos, L.; Baranski, M.; Maurer, V.; Cozzi, G.; Bapst, B.; et al. Impact of US Brown Swiss genetics on milk quality from low-input herds in Switzerland: Interactions with season. Food Chem. 2018, 251, 93–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NMKL 186. Trace Elements—As, Cd, Hg, Pb and Other Elements. Determination by ICP-MS after Pressure Digestion. Available online: https://www.nmkl.org/index.php/en/publications/item/tungmetaller-as-cd-hg-og-pb-bestemmelse-med-icp-ms-etter-syreoppslutning-under-trykk-nmkl-186-2007 (accessed on 10 August 2021).
- Gilmour, A.R.; Thompson, R.; Cullis, B.R. Average information REML: An efficient algorithm for variance parameter estimation in linear mixed models. Biometrics 1995, 51, 1440–1450. [Google Scholar] [CrossRef]
- ter Braak, C.; Smilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, Version 5.0. Available online: https://research.wur.nl/en/publications/canoco-reference-manual-and-users-guide-software-for-ordination-v (accessed on 10 August 2021).
- Schwendel, B.H.; Wester, T.J.; Morel, P.C.H.; Tavendale, M.H.; Deadman, C.; Shadbolt, N.M.; Otter, D.E. Invited review: Organic and conventionally produced milk—An evaluation of influence factors on milk composition. J. Dairy Sci. 2015, 98, 721–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, S.M.; DePeters, E.J.; Taylor, S.J.; Rosenberg, M.; Perez-Monti, H.; Capps, V. Milk composition of Holstein, Jersey, and Brown Swiss cows in response to increasing levels of dietary fat. Anim. Feed. Sci. Technol. 2006, 131, 451–473. [Google Scholar] [CrossRef]
- Davis, H.; Stergiadis, S.; Chatzidimitriou, E.; Sanderson, R.; Leifert, C.; Butler, G. Meeting Breeding Potential in Organic and Low-Input Dairy Farming. Front. Vet. Sci. 2020, 7, 544149. [Google Scholar] [CrossRef] [PubMed]
- Couvreur, S.; Hurtaud, C.; Lopez, C.; Delaby, L.; Peyraud, J.L. The linear relationship between the proportion of fresh grass in the cow diet, milk fatty acid composition, and butter properties. J. Dairy Sci. 2006, 89, 1956–1969. [Google Scholar] [CrossRef]
- Bauman, D.E.; Griinari, J.M. Regulation and nutritional manipulation of milk fat: Low-fat milk syndrome. Livest. Prod. Sci. 2001, 70, 15–29. [Google Scholar] [CrossRef]
- O’Callaghan, T.F.; Hennessy, D.; McAuliffe, S.; Kilcawley, K.N.; O’Donovan, M.; Dillon, P.; Ross, R.P.; Stanton, C. Effect of pasture versus indoor feeding systems on raw milk composition and quality over an entire lactation. J. Dairy Sci. 2016, 99, 9424–9440. [Google Scholar] [CrossRef] [PubMed]
- Walker, G.P.; Stockdale, C.R.; Wales, W.J.; Doyle, P.T.; Dellow, D.W. Effect of level of grain supplementation on milk production responses of dairy cows in mid-late lactation when grazing irrigated pastures high in paspalum (Paspalum dilatatum Poir.). Aust. J. Exp. Agric. 2001, 41, 1–11. [Google Scholar] [CrossRef]
- Davis, H.P.; Hathaway, I.L. The Vitamin A content of the Milk of Holstein, Ayrshire, Jersey, and Guernsey Cows; Agricultural Experiment Station of Nebraska: Lincoln, NE, USA, 1931; ISSN 0097-1385. [Google Scholar]
- Walker, G.P.; Dunshea, F.R.; Doyle, P.T. Effects of nutrition and management on the production and composition of milk fat and protein: A review. Aust. J. Agric. Res. 2004, 55, 1009–1028. [Google Scholar] [CrossRef]
- Manuelian, C.L.; Penasa, M.; Visentin, G.; Zidi, A.; Cassandro, M.; De Marchi, M. Mineral composition of cow milk from multibreed herds. Anim. Sci. J. 2018, 89, 1622–1627. [Google Scholar] [CrossRef] [PubMed]
- Rius, A.G.; Appuhamy, J.A.D.R.N.; Cyriac, J.; Kirovski, D.; Becvar, O.; Escobar, J.; McGilliard, M.L.; Bequette, B.J.; Akers, R.M.; Hanigan, M.D. Regulation of protein synthesis in mammary glands of lactating dairy cows by starch and amino acids. J. Dairy Sci. 2010, 93, 3114–3127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Bermudez, R.; Miranda, M.; Baudracco, J.; Fouz, R.; Pereira, V.; Lopez-Alonso, M. Breeding for organic dairy farming: What types of cows are needed? J. Dairy Res. 2019, 86, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Gaucheron, F. The minerals of milk. Reprod. Nutr. Dev. 2005, 45, 473–483. [Google Scholar] [CrossRef]
- Zwierzchowski, G.; Ametaj, B.N. Minerals and Heavy Metals in the Whole Raw Milk of Dairy Cows from Different Management Systems and Countries of Origin: A Meta-Analytical Study. J. Agric. Food Chem. 2018, 66, 6877–6888. [Google Scholar] [CrossRef]
- Pinchen, H.; Powell, N.; Weiner, D.; Finglas, P. McCance and Widdowson’s The Composition of Foods Integrated Dataset. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/788485/McCance_Widdowson_Comp_Foods_Integrated_Dataset_User_Guide_2019__1_.pdf (accessed on 10 August 2021).
- FAO. Evaluation of Certain Food Contaminants: Sixty-Fourth Report of the Joint FAO/WHO Expert Committee on Food Additives; FAO: Rome, Italy, 2006. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Parameters Assessed | Conventional | Organic | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean 1 | SE | p-Value | Mean 1 | SE | p-Value | |||||
| CHP n = 118 | CSP n = 120 | CLP n = 120 | OHP n = 96 | OSP n = 91 | OLP n = 96 | |||||
| Herd composition (% herd) | ||||||||||
| Holstein | 63.8 | 89.1 | 96.4 | 10.19 | 0.077 | 40.2 | 59.8 | 87.3 | 13.62 | 0.070 |
| British Friesian | 2.8 | 10.0 | 1.7 | 6.05 | 0.579 | 2.3 | 0.3 | 1.8 | 1.26 | 0.528 |
| Ayrshire | 0 | 0 | 0.6 | 0.34 | 0.381 | 12.9 | 12.8 | 0.0 | 9.95 | 0.580 |
| New Zealand Friesian | 0.5 | 0 | 0 | 0.24 | 0.371 | 4.2 | 0.0 | 0.0 | 2.42 | 0.387 |
| Jersey | 0.1 | 0 | 1.2 | 0.70 | 0.413 | 0.4 | 4.6 | 2.2 | 2.81 | 0.579 |
| Scandinavian Red | 0.4 | 0 | 0 | 0.20 | 0.371 | 1.0 | 1.1 | 0.8 | 0.74 | 0.944 |
| Shorthorn | 0 | 0 | 0.003 | 0.0019 | 0.384 | 1.0 | 0.1 | 0.6 | 0.36 | 0.271 |
| Brown Swiss | 0 | 0.9 | 0 | 0.55 | 0.381 | 0.3 | 1.0 | 4.9 | 2.90 | 0.490 |
| Guernsey | 0.1 | 0 | 0.1 | 0.10 | 0.563 | n.a.3 | n.a. 3 | n.a. 3 | n.a. 3 | n.a. 3 |
| Other breeds/crossbreds | 32.4 a | 0 b | 0 b | 8.35 | 0.014 | 37.7 | 20.3 | 2.3 | 13.03 | 0.183 |
| Intakes of feed (% dry matter intake unless otherwise stated) | ||||||||||
| Dry matter intake (kg/d) | 19.3 b | 20.8 a | 20.6 a | 0.37 | 0.015 | 17.5 b | 18.3 a,b | 19.8 a | 0.53 | 0.019 |
| Total forage | 70.2 a | 60.1 b | 54.4 c | 1.91 | <0.001 | 77.7 a | 76.5 a | 68.7 b | 1.40 | < 0.001 |
| Total concentrate | 29.8 c | 39.9 b | 45.7 a | 1.91 | <0.001 | 22.3 b | 23.5 b | 31.3 a | 1.40 | <0.001 |
| Pasture intake | 26.0 a | 6.2 b | 0.6 c | 1.23 | <0.001 | 37.3 a | 25.1 b | 13.9 c | 1.48 | <0.001 |
| Grass/clover silage 2 | 21.8 | 23 | 22.6 | 2.77 | 0.948 | 32.2 | 35.4 | 35.2 | 2.72 | 0.654 |
| Maize silage | 18.6 b | 27.1 a | 27.9 a | 2.24 | 0.010 | 0 | 0 | 3.2 | 1.20 | 0.122 |
| Lucerne silage | 0 | 0 | 0.5 | 0.22 | 0.167 | 0 | 0.2 | 0 | 0.08 | 0.379 |
| Other Mixed silage | 0.1 | 0.5 | 0 | 0.21 | 0.225 | 1.7 | 3.7 | 3.6 | 1.44 | 0.542 |
| Wholecrop silage | 1.2 | 0.6 | 0.1 | 0.36 | 0.102 | 4.3 b | 8.9 a,b | 10.8 a | 1.71 | 0.038 |
| Hay and straw | 2.5 | 2.7 | 2.5 | 0.83 | 0.982 | 2.3 | 2.7 | 2.1 | 0.73 | 0.784 |
| Moist by-product | 2.8 | 3.2 | 6.7 | 1.57 | 0.165 | 0.3 | 0.5 | 0.2 | 0.23 | 0.632 |
| Dry straight | 5.0 | 10.4 | 11.2 | 3.66 | 0.436 | 0.5 | 0.3 | 1.5 | 0.42 | 0.136 |
| Cereal | 2.3 | 0.9 | 3.4 | 1.03 | 0.270 | 3.2 | 3.0 | 5.5 | 1.29 | 0.302 |
| Blend | 19.5 | 24.3 | 23.2 | 4.23 | 0.694 | 18.2 | 19.7 | 24.1 | 1.99 | 0.125 |
| Oil | 0.2 b | 1.0 a | 1.2 a | 0.24 | 0.014 | 0.03 | 0 | 0 | 0.017 | 0.391 |
| Mineral (g/cow/day) | 65.2 | 114.1 | 158.3 | 26.04 | 0.057 | 54.8 | 45.4 | 103.1 | 17.32 | 0.059 |
| Vitamin (g/cow/day) | 0 | 0.5 | 11.7 | 5.45 | 0.244 | 0 | 0 | 3.2 | 1.20 | 0.122 |
| Parameters Assessed | Conventional | Organic | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean 1 | SE | p-Value | Mean 1 | SE | p-Value | |||||
| CHP n = 118 | CSP n = 120 | CLP n = 120 | OHP n = 96 | OSP n = 91 | OLP n = 96 | |||||
| Yield and basic composition | ||||||||||
| Milk yield (kg/d) | 26.9 b | 32.3 a | 30.2 a | 1.04 | 0.004 | 19.3 b | 21.4 b | 25.3 a | 1.04 | 0.002 |
| Fat (g/kg milk) | 39.9 | 38.4 | 39.1 | 0.44 | 0.085 | 40.3 a | 40.8 a | 38.5 b | 0.58 | 0.032 |
| Protein (g/kg milk) | 33.8 | 32.9 | 33.2 | 0.28 | 0.100 | 34.1 a | 33.6 a | 32.2 b | 0.46 | 0.023 |
| Lactose (g/kg milk) | 45.0 | 45.2 | 45.4 | 0.13 | 0.172 | 44.4 b | 44.8 a,b | 45.3 a | 0.19 | 0.019 |
| SCC (×1000/mL milk) | 162.9 | 149.3 | 143.8 | 10.00 | 0.492 | 167.5 | 121.6 | 132.5 | 11.12 | 0.076 |
| Macrominerals (mg/kg milk) | ||||||||||
| Ca | 1015.7 a | 966.2 b | 1010.8 a | 9.77 | 0.001 | 1065.5 a | 1057.7 a | 1021.2 b | 9.76 | 0.008 |
| K | 1364.8 | 1352.5 | 1380.1 | 12.74 | 0.246 | 1369.2 | 1386.8 | 1378.3 | 14.27 | 0.857 |
| Mg | 95.5 | 95.4 | 97.4 | 1.03 | 0.511 | 96.7 | 95.0 | 92.8 | 1.02 | 0.069 |
| Na | 341.6 | 340.8 | 348.7 | 5.01 | 0.652 | 360.0 | 346.8 | 331.5 | 6.03 | 0.066 |
| P | 799.7 a | 774.8 b | 803.8 a | 8.37 | 0.045 | 817.3 | 810.5 | 794.0 | 8.85 | 0.246 |
| Essential trace elements (μg/kg milk unless otherwise stated) | ||||||||||
| Cu | 62.1 | 61.9 | 58.4 | 3.52 | 0.815 | 51.7 | 51.2 | 55.8 | 3.72 | 0.198 |
| Fe (mg/kg milk) | 2.58 | 3.01 | 1.67 | 0.779 | 0.557 | 0.44 | 0.68 | 0.95 | 0.168 | 0.185 |
| I | 301.8 | 314.4 | 384.2 | 21.21 | 0.626 | 394.8 | 341.8 | 300.9 | 33.13 | 0.782 |
| Mn | 54.2 | 49.4 | 43.4 | 8.53 | 0.460 | 30.2 | 28.8 | 28.4 | 2.49 | 0.804 |
| Mo | 62.2 | 59.5 | 61.2 | 2.55 | 0.713 | 75.5 | 72.4 | 73.3 | 4.12 | 0.995 |
| Zn (mg/kg milk) | 5.07 | 5.07 | 5.10 | 0.210 | 0.989 | 4.78 | 4.25 | 4.50 | 0.238 | 0.255 |
| Non-essential trace elements | ||||||||||
| Al (mg/kg milk) | 1.37 | 1.65 | 1.08 | 0.440 | 0.736 | 0.24 | 0.31 | 0.39 | 0.075 | 0.421 |
| Sn (μg/kg milk) | 3.26 | 2.60 | 2.10 | 0.394 | 0.128 | 4.05 a | 2.40 b | 1.72 b | 0.625 | 0.016 |
| RNI/PTWI % Contributed by Liquid Milk Intake | ||||
|---|---|---|---|---|
| Element | RNI/PTWI 1 | HP | SP | LP |
| Conventional production system | ||||
| Ca | 350 mg/d | 72.1 | 68.6 | 71.7 |
| P | 270 mg/d | 73.6 | 71.3 | 74.0 |
| Organic production system | ||||
| Ca | 350 mg/d | 75.6 | 75.1 | 72.5 |
| Sn | 175 mg/week | 0.004 | 0.002 | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stergiadis, S.; Qin, N.; Faludi, G.; Beauclercq, S.; Pitt, J.; Desnica, N.; Pétursdóttir, Á.H.; Newton, E.E.; Angelidis, A.E.; Givens, I.; et al. Mineral Concentrations in Bovine Milk from Farms with Contrasting Grazing Management. Foods 2021, 10, 2733. https://doi.org/10.3390/foods10112733
Stergiadis S, Qin N, Faludi G, Beauclercq S, Pitt J, Desnica N, Pétursdóttir ÁH, Newton EE, Angelidis AE, Givens I, et al. Mineral Concentrations in Bovine Milk from Farms with Contrasting Grazing Management. Foods. 2021; 10(11):2733. https://doi.org/10.3390/foods10112733
Chicago/Turabian StyleStergiadis, Sokratis, Nanbing Qin, Gergely Faludi, Stephane Beauclercq, Joe Pitt, Natasa Desnica, Ásta H. Pétursdóttir, Eric E. Newton, Angelos E. Angelidis, Ian Givens, and et al. 2021. "Mineral Concentrations in Bovine Milk from Farms with Contrasting Grazing Management" Foods 10, no. 11: 2733. https://doi.org/10.3390/foods10112733
APA StyleStergiadis, S., Qin, N., Faludi, G., Beauclercq, S., Pitt, J., Desnica, N., Pétursdóttir, Á. H., Newton, E. E., Angelidis, A. E., Givens, I., Humphries, D. J., Gunnlaugsdóttir, H., & Juniper, D. T. (2021). Mineral Concentrations in Bovine Milk from Farms with Contrasting Grazing Management. Foods, 10(11), 2733. https://doi.org/10.3390/foods10112733