Abstract
Helicobacter pylori is a cause of gastric cancer. We extracted the exopolysaccharide (EPS) of Lactobacillus plajomi PW-7 for antibacterial activity versus H. pylori, elucidating its biological activity and structural characteristics. The minimum inhibitory concentration (MIC) of EPS against H. pylori was 50 mg/mL. Disruption of the cell membranes of pathogenic bacteria by EPS was indicated via the antibacterial mechanism test and confirmed through electron microscopy. EPS also has antioxidant capacity. The IC50 of EPS for 2,2-Diphenyl-1-picrylhydrazyl (DPPH) radical, superoxide anions, and hydroxyl radicals were 300 μg/mL, 180 μg/mL, and 10 mg/mL, respectively. The reducing power of EPS was 2 mg/mL, equivalent to 20 μg/mL of ascorbic acid. EPS is a heteropolysaccharide comprising six monosaccharides, with an approximate molecular weight of 2.33 × 104 Da. Xylose had a significant effect on H. pylori. EPS from L. plajomi PW-7 showed potential as an antibacterial compound and antioxidant, laying a foundation for the development of EPS-based foods.
1. Introduction
Fifty percent of the global population have gastric Helicobacter pylori. The International Cancer Society identify it as a category I carcinogen. The prevalence of gastric cancer in Asia, particularly East Asia, is significantly higher than the average worldwide [1,2]. H. pylori can only survive in gastric acid for a short time. It parasitizes the gastric antrum, resulting in chronic inflammation. The resulting destruction of the mucosal barrier is long lasting and leads to gastritis, gastric and duodenal ulcers, and gastric cancer [3,4]. Current H. pylori treatment is mainly based on quadruple antibiotic therapy. However, there are problems with this approach [5]. Antibiotics such as clarithromycin and amoxicillin impact the intestinal flora and have many shortcomings, including varying side effects and antibiotic resistance [6,7]. Caring for a wide range of infected people is problematic. There is thus an unmet medical need for molecules with low antibiotic resistance and reduced side effects able to reduce or prevent H. pylori in high-risk groups on a large scale.
Lactic acid bacteria (LAB) are beneficial intestinal microorganisms [8,9]. They are present in many foods, including yogurt, sauerkraut, soy milk, kimchi, and fermented vegetables [10]. LAB have been used in the pharmacy industry to regulate intestinal flora, and even to treat dyspepsia, enteritis, and diarrhea [11]. Moreover, LAB could be used as tanning agents and biodegradable materials in the chemical industry [12]. The literature showed that LAB can assist in the treatment of H. pylori. When bismuth quadruple therapy, standard triple therapy, and high-dose PPI Bismuth-containing quadruple therapy were used with LAB, the side effects of antibiotics (diarrhea, constipation, etc.) were reduced and the treatment could be sped up [5,13,14]. In the literature, some lactic acid bacteria have an antibacterial effect on H. pylori directly; some studies reported that the supernatant contained the substance that inhibited H. pylori [15,16].
Metabolites of LAB, such as EPS, organic acids, bacteriocins, and fatty acids, display beneficial biological activities, such as antibacterial, antioxidant, anticancer, and cholesterol-lowering activities [17,18]. Many studies indicated LAB can inhibit the growth of pathogenic bacteria by secreting acid and bacteriocins [18,19]. LAB exopolysaccharide (EPS) is also receiving increased attention due to the potential health benefits of EPS [20,21,22]. Ryan et al. found that the acid and bacteriocin secreted by L. salivarius UCC119 could not inhibit H. pylori, suggesting the involvement of other mediators [23]. Liu et al. found that EPS enhanced rUreB-specific protective immunity against H. pylori [24]. L. fermentum UCO-979C EPS also inhibits H. pylori colonization [25].
As well as antibacterial activity, LAB EPS also possesses other biological activities, including antioxidant, anti-tumor, and anti-diabetic properties [20,26,27]. It has also found wide application in health care and the food industry. A systematic evaluation of the available literature indicates that EPS can inhibit pathogenic bacteria, although its antibacterial mechanism against H. pylori is underexplored.
We isolated a strain of lactic acid bacteria from Sichuan pickles and found that its supernatant had a strong inhibitory effect on H. pylori. Our study aimed to elucidate the H. pylori-inhibiting components that are fermented by L. plajomi PW-7 and to clarify its activity on H. pylori. Furthermore, the mechanism of the antibacterial and antioxidant activity of the substances were investigated.
2. Materials and Methods
2.1. Materials
H. pylori was obtained from the American Type Culture Collection (ATCC). H. pylori growth medium consisted of ATCC®18 media (trypsin bean soup) and ATCC®260 media (tryptic soy agar, 5% defibrated sheep blood (Nanjing maojie microbial technology Co., Ltd., Nanjing, China)) (Qingdao hope bio-technology Co., Ltd., Qingdao, China). A micro aerobic gas bag (Mitsubishi gas chemical Co., Ltd., Tokyo, Japan). Aeromonas hydrophila, Aeromonas salmonicida, Bacillus subtilis, Escherichia coli, Pseudomonas aeruginosa, Staphylococcus aureus, and Vibrio parahaemolyticus were provided by the Provincial Marine Resources Development Research Institute (Lianyungang, China) and were cultivated using Luria–Bertani (LB) (Aobox, Beijing, China). Lactobacillus plajomi PW-7 was selected from indigenous Sichuan pickles (vegetables: radish, cauliflower, carrot, cowpea, celery, ginger, and green and red pepper with 5% of NaCl) and cultivated in MRS broth (Beijing luqiao technology Co., Ltd., Beijing, China) at 37 °C.
2.2. Screening of LAB
Ten milliliters of Sichuan pickle water was added to MRS broth to form a 3% inoculation, and subsequently cultured at 37 °C for 24 h. The enrichment broth was then diluted, using gradient dilution, using concentrations of 10−3, 10−4, and 10−5. Plates were spread with MRS medium in 50 μL aliquots. Plates were put into an anaerobic tank and incubated for 24 h at 37 °C. The colonies with a calcified zone were selected for subsequent experiments [28].
Selected colonies were inoculated into MRS broth and incubated at 37 °C for 24 h. The fermentation broth was then centrifuged at 8000 rpm for 15 min, and the supernatant was used for Oxford cup bacteriostasis tests, with H. pylori as the indicator bacterium; is the test was based on the appearance of transparent circle. Strains inhibiting H. pylori were selected and pure strains isolated using streak plates [29].
2.3. Morphology of Strains
Colony characteristics: the purified single colony was cultured at 37 °C and observed. The cell morphology was recorded using scanning electron microscopy (SEM, model JFC-1600; JSM-6390LA; JEOL, Tokyo, Japan).
2.4. 16S rDNA Gene Identification
The genomic DNA of strains was extracted using a PCR kit. PCR amplification was performed using universal primers 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-GGTTACCTTGTTACGCTT-3′). PCR products were purified and sequenced by Sangon Biotech (Shanghai). NCBI Blast (https://blast.ncbi.nlm.nih.gov, 3 September 2020) was used to analyze the resulting sequences. Phylogenetic trees were drawn using MEGA 6 with Neighbor-Joining [30].
2.5. Analysis of Components of Strain PW-7 Inhibiting H. pylori
2.5.1. Hydrogen Peroxide Detection and Elimination
The pH of the fermentation supernatant of strain PW-7 was adjusted to 7.0. Catalase was added, and the final concentration adjusted to 1 mg/mL, before being incubated for 2 h at 37 °C. The solution was then placed in boiling water for 5 min, before the pH was adjusted to 5.0 for hydrogen peroxide detection. Supernatant without catalase treatment (Aladdin, Shanghai, China) was used as a control. H. pylori was the indicator bacterium, and 107 CFU/mL suspension was applied to TCC®260 medium, cultured at 37 °C for 12 h. An Oxford cup antibacterial test was conducted [31].
2.5.2. Detection and Elimination of Organic Acids
The pH of the PW-7 fermentation supernatant was adjusted to 7.0 using NaOH. Untreated supernatant was used as a control. H. pylori was used as the indicator bacteria, and the method was the same as that of 2.5.1 [32].
2.5.3. Bacteriostatic Test of Bacteriocin
The fermentation supernatant was precipitated using ammonium sulfate (20–70%) (Aladdin, Shanghai, China), dialyzed, and then lyophilized (Labconco FreeZon®2.5L USA). After Gram-negative and Gram-positive bacteria reached the logarithmic growth phase, 107 CFU/mL bacterial suspension was coated on LB, put on Oxford cup, and PW-7 bacteriocin (100 mg/mL) was added and cultured at 37 °C for 12 h. The culture of H. pylori was the same as 2.5.1. The bacteriostatic ability of PW-7 bacteriocin was determined for indicator bacteria using the Oxford cup method [33].
2.5.4. Bacteriostatic Test of EPS
The fermentation supernatant was precipitated for 12 h using 3× ethanol at 4 °C. The precipitate was dissolved, and 4% Trichloroacetic acid (TCA) (Aladdin, Shanghai, China) was added. The resulting solution was left to stand for 30 min before being centrifuged (Thermo Heraeus Multifuge X3R Centrifuge, Eppendorf, Hamburg, Germany), and the supernatant was isolated. As with the bacteriocin antibacterial test, the inhibitory effect of EPS on each indicator bacterium was determined [34].
2.6. Structure Characterization of EPS
2.6.1. Monosaccharide Composition Analysis
A chromatographic DionexCarbopacTMPA20 (3 × 150) column (Dionex™ ICS-5000+ TC Thermal Compartment, ThermoFisher, Waltham, MA, USA) was used to determine monosaccharide composition. The mobile phase was: A: H2O, B: 15 mM NaOH, C: 15 mM NaOH and 100 mM NaOAc; flow rate: 0.3 mL/min; injection volume: 5 μL; column temperature: 30 °C. Sixteen monosaccharide standards (fucose, rhamnose, arabinose, galactose, glucose, xylose, mannose, fructose, ribose, galacturonic acid, glucuronic acid, galactosamine hydrochloride, glucosamine hydrochloride, N-acetyl-D-glucosamine, gulosaldehyde acid, and mannuronic acid) (Borui sugar Biotechnology Co., Ltd., Yangzhou, China) were dissolved in standard solution. Ten micrograms of polysaccharide were placed in ampoules. An amount of 10 mL of Trifluoroacetic acid (3 M) was added, and the solution then underwent hydrolysis for 3 h at 120 °C. The resulting solution was accurately absorbed, transferred to a tube, and dried under nitrogen. An amount of 10 mL of water was added; before vortexing and being absorbed at 100 μL, 900 μL of deionized water was added, and the solution was centrifuged at 12,000 rpm for 5 min. The supernatant was isolated for ion chromatography (IC) analysis [35].
2.6.2. Determination of Molecular Weight
Polysaccharide molecular weight and purity were determined at 25 °C using high-performance gel permeation chromatography (HPGPC) (Shimadzu LC-10A HPLC, Shimadzu, Kyoto, Japan), with ultrapure water as the mobile phase and the flow rate set to 1.0 mL/min. A 2 mg/mL polysaccharide sample solution was prepared using a disposable microporous filter membrane (0.22 μm). After filtration, 100 μL was injected into the chromatograph. By using the elution peak times and multiplicity, the sample purity and average molecular weight were calculated using GPC software (Agilent, Palo Alto, CA, USA ) [36].
2.7. Minimum Inhibitory Concentration (MIC)
Suspensions of E. coli, S. aureus, and H. pylori were diluted gradually. A suspension at 107 CFU/mL was used as a standard. In the experimental group, 2 mL of LB culture solution and then 40 μL of spare bacterial solution were added to test tubes. EPS was added to each tube to reach a series of final concentrations: 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100 mg/mL. The control group was prepared similarly, without the addition of EPS. Test tubes were incubated at 37 °C for 24 h. After culture, 100 μL of the bacterial broth was spread on LB plates. Observations were made after 24 h of inverted culture at 37 °C. The EPS concentration of the first plate with a single observable colony was taken as the lowest inhibitory concentration [37].
2.8. Antibacterial Mechanism
2.8.1. Cell Membrane Permeability
Using the method of Qiao et al. [38], E. coli, H. pylori, and S. aureus in their logarithmic growth phase were inoculated into LB with an EPS concentration of 1× MIC and 2× MIC at 2% inoculum volume. After mixing, samples were cultured at 37 °C. LB without added samples acted as controls. Samples were measured every 2 h for 8 h. Supernatant was centrifuged at 3500 rpm for 10 min. Absorbance was measured at 450 nm using a full wavelength microplate reader (Multiskan GO, Thermo Scientific, Waltham, MA, USA). A bacterial culture medium without samples was used as the control.
2.8.2. Cell Membrane Integrity
Cell membrane integrity was determined by measuring the release of cell components. E. coli, H. pylori and S. aureus were cultured to the logarithmic phase, then centrifuged at 5000 rpm for 5 min. The precipitate was washed with 0.1 M PBS (pH 7.4) (Aladdin, Shanghai, China) three times before resuspension. To measure protein and reduce sugar concentrations in the supernatant, two milliliters of cell suspension were cultured with blank, 1× MIC, and 2× MIC for 4 h before being centrifuged at 10,000 rpm for 5 min. The released nucleic acid concentration was determined at 260 nm [39].
2.8.3. NPN Uptake
The uptake of N-phenyl-1-naphthylamine (NPN) was determined using the method of Zhang et al. [40]. Briefly, E. coli, H. pylori, and S. aureus in the logarithmic phase were centrifuged at 5000 rpm for 10 min, then washed with 0.5% NaCl three times before being re-suspended to an OD600nm of 0.5. The suspension was treated with different EPS concentrations (1× MIC, 2× MIC) for 1 h. After centrifugation, the suspension was washed and re-suspended using 0.5% NaCl. An amount of 200 mL of bacterial suspension was mixed with 1.5 μL of NPN (100 mM) (Aladdin, Shanghai, China). Fluorescence was measured at 350 nm and 420 nm using a full wavelength multifunctional microplate reader (Infinite M1000 Pro, TECAN, Homrechtikon, Switzerland).
2.8.4. Scanning Electron Microscopy
For the experimental group, 2 mL of LB was added to each test tube, then logarithmic phase broth (2% inoculation), before EPS was added to reach a final EPS concentration of 2× MIC. Samples were cultured at 37 °C for 1 h, then centrifuged at 5000 rpm for 10 min to remove supernatant. For the control group, logarithmic phase broth was centrifuged directly. The precipitate was removed before being washed three times with 0.2 M PBS (pH 7.4) and then fixed with 2.5% glutaraldehyde (Aladdin, Shanghai, China). Mixing occurred overnight. Slides were soaked in 75% ethanol and then allowed to dry. Different gradients of bacterial solution were applied to the slide before the addition of 0.2 M PBS (pH 7.4) followed by spreading, drying, and washing with 30%, 50%, 70%, 80%, 90%, and 100% ethanol as required by SEM [41].
2.9. Antioxidant Activity of Exopolysaccharides In Vitro
2.9.1. DPPH Free Radical Scavenging Experiment
The rate of DPPH free radical scavenging was determined using the method of Dong [42]. Two hundred microliters of EPS at different concentrations were added to 200 μL of DPPH (0.1 mM of absolute ethanol) (Aladdin, Shanghai, China). After mixing, absorbance was measured at 517 nm after equilibrating at room temperature for 30 min. Ascorbic acid (Vc) (Aladdin, Shanghai, China) was used as a positive control. DPPH free radical scavenging activity was expressed as:
where Aa is the absorbance of the DPPH free radical solution and polysaccharide sample; Ab is the absorbance of anhydrous ethanol and polysaccharide; and Ac is the absorbance of the DPPH radical solution and absolute ethanol.
Scavenging ability (%) = [1 − (Aa − Ab)/Ac] × 100
2.9.2. Hydroxyl Radical Scavenging Experiment
Amounts of 100 μL of FeSO4·7H2O (9 mM), 100 μL of salicylic acid anhydrous ethanol solution (9 mM), 100 μL of 0.03% H2O2 (Aladdin, Shanghai, China), and 100 μL of polysaccharide samples at different concentrations were mixed for 15 m at 37 °C. Absorbance was measured at 510 nm, with water as a blank [43]. Hydroxyl radical scavenging activity was calculated using:
where Aa is the absorbance value of the polysaccharide sample; Ab is the absorbance value without H2O2; and Ac was the blank control.
Scavenging ability (%) = [(Ac − (Aa − Ab))/Ac] × 100
2.9.3. Superoxide Anion Radical Scavenging Experiment
Superoxide anion was modified using the method of Zhang [44]. Two hundred microliters of polysaccharide samples at different concentrations were mixed with 1 mL of Tris HCl (50 mM pH 8.2) and then incubated for 10 min at 25 °C. Thirty microliters of pyrogallol (6 mM) (Aladdin, Shanghai, China) was added. The reaction was then kept in the dark for 30 min. Thirty microliters of HCl (6 mM) was added to terminate the reaction. Absorbance was measured at 320 nm, with water used as a control. The scavenging rate of superoxide anion radical was calculated as:
where Aa is the absorbance of polysaccharide; Ab is the absorbance without pyrogallol; and Ac is the absorbance of the control.
Scavenging activity (%) = [1 − (Aa − Ab)/Ac] × 100
2.9.4. Reducing Power
Amounts of 200 μL of polysaccharide samples with different concentrations, 200 μL of PBS (0.2 M, pH 6.6), and 200 μL of potassium hexacyanoferrate (1%, w/v) (Aladdin, Shanghai, China) were mixed, then cultured at 50 °C for 20 min, with 200 μL of TCA (10%) added to terminate the reaction. After centrifugation at 5000 rpm for 10 min, 500 μL of supernatant was mixed for 10 min with 100 μL of FeCl3 (0.2%) (Aladdin, Shanghai, China) and 2.5 mL of water. Absorbance was measured at 700 nm [44].
2.10. Statistical Significance
Experiments were repeated three times. Data are presented as means ± SD. SPSS statistics, version 17.0 for windows, was used for statistical analysis. p < 0.05 was the threshold for statistical significance.
3. Results and Discussions
3.1. Screening and Identification of Strain
3.1.1. Screening of Strain
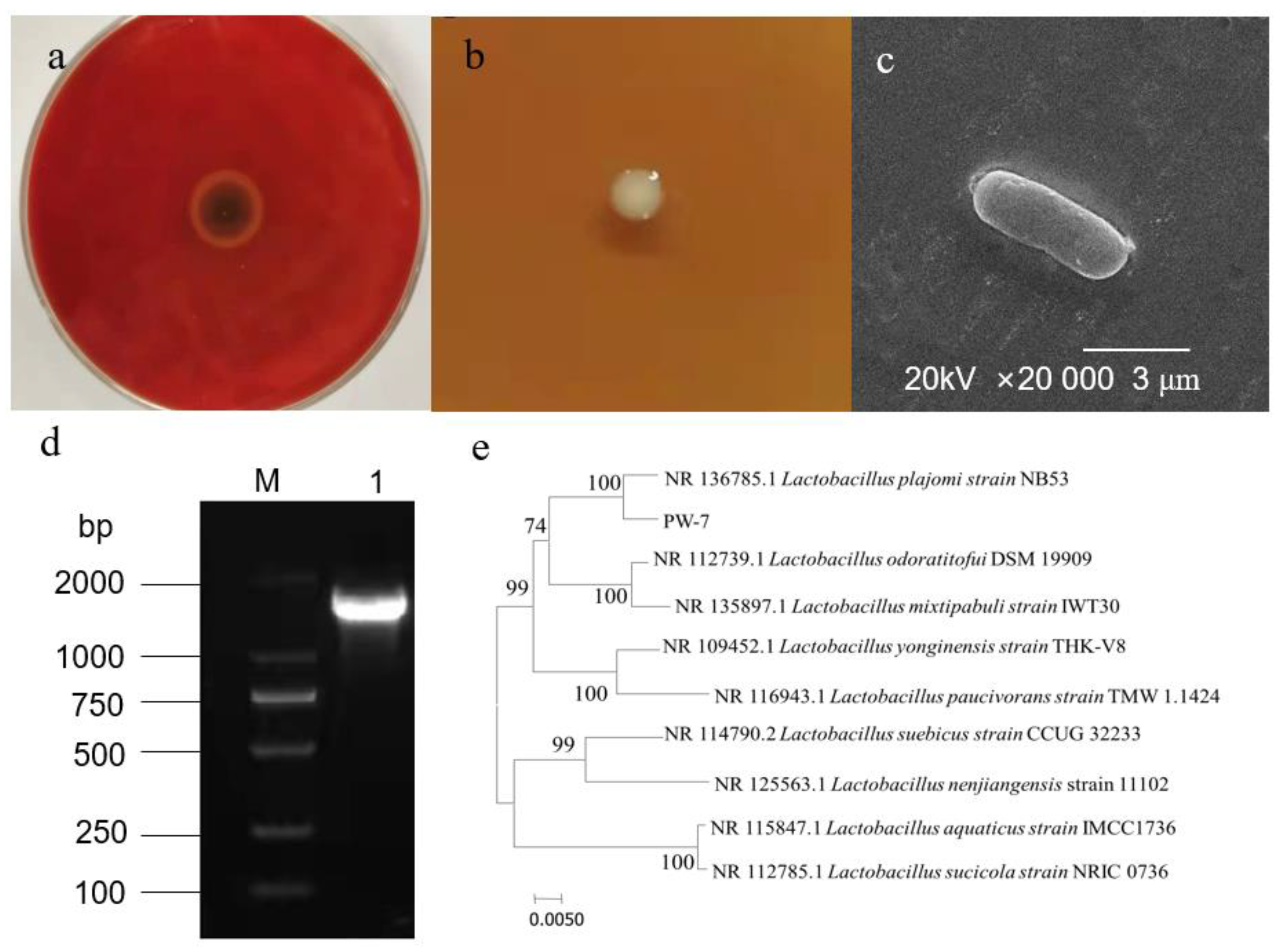
Ten strains of LAB were isolated from pickle samples. Four strains displaying an inhibition zone against H. pylori were screened further. An optimal strain exhibiting enhanced inhibition was selected. We named this strain PW-7 (Figure 1a).
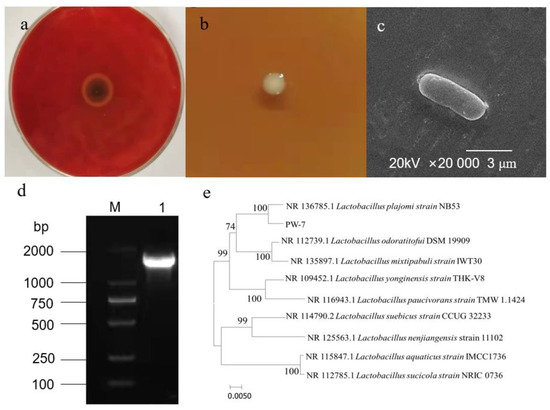
Figure 1.
(a): Antibacterial effect of PW-7 on H. pylori; (b): colony morphology of strain PW-7; (c): scanning electron microscopy of PW-7 morphology; (d): agarose gel electrophoresis of 16S rDNA PCR products; M: DNA marker; 1: strain PW-7 16S rDNA PCR amplification product; (e): phylogenetic tree based on 16S rDNA gene sequences.
3.1.2. Morphological Characteristics of Strain PW-7
3.1.3. Sequence Analysis of 16S rDNA of Strain PW-7
The genome of strain PW-7 was extracted, and 16S rDNA was amplified using universal primers and detected using agarose gel electrophoresis (see Figure 1d). The fragment size was approximately 1.5 kb. The resulting PW-7 gene sequence was submitted to GenBank. By using sequence similarity comparison, it was determined that PW-7 belonged to the genus Lactobacillus. Sequences from closely related strains were compared using MEGA 7, with the neighbor-joining method used to construct a phylogenetic tree. This showed that PW-7 and Lactobacillus plajomi were closely related (Figure 1e). On the basis of our analysis of its cellular morphology, its gene sequence, and its biochemical and physiological characteristics, strain PW-7 was classified as Lactobacillus plajomi PW-7.
3.2. Analysis of Components of Strain PW-7 Inhibiting H. pylori
When compared to untreated PW-7 fermentation broth, catalase treatment did not decrease the inhibitory effect of the broth, indicating that inhibition was not caused by hydrogen peroxide (see Table S1). When the pH of the fermentation supernatant was adjusted to 7.0, its antibacterial ability did not decrease, indicating inhibition was not caused by release of acid (Table S2). Bacteriocins did not inhibit Gram-negative and Gram-positive bacteria (Table S3), suggesting that bacteriocin was not responsible for the observed antimicrobial activity, and that other bacteriostatic substances may be present. EPS can inhibit Gram-negative and Gram-positive bacteria (Table 1).

Table 1.
Antibacterial spectrum of EPS of strain PW-7.
A review of the literature suggested that EPS from LAB can inhibit pathogenic bacteria. EPS from Lactobacillus plantarum WLPL04 and EPS from Lactobacillus rhamnosus are known to inhibit pathogenic micro-organisms [43,45]. We conclude that the EPS of L. plajomi PW-7 is a substance with antibacterial effect.
3.3. Structure Analysis of PW-7 EPS
3.3.1. Monosaccharide Composition of EPS
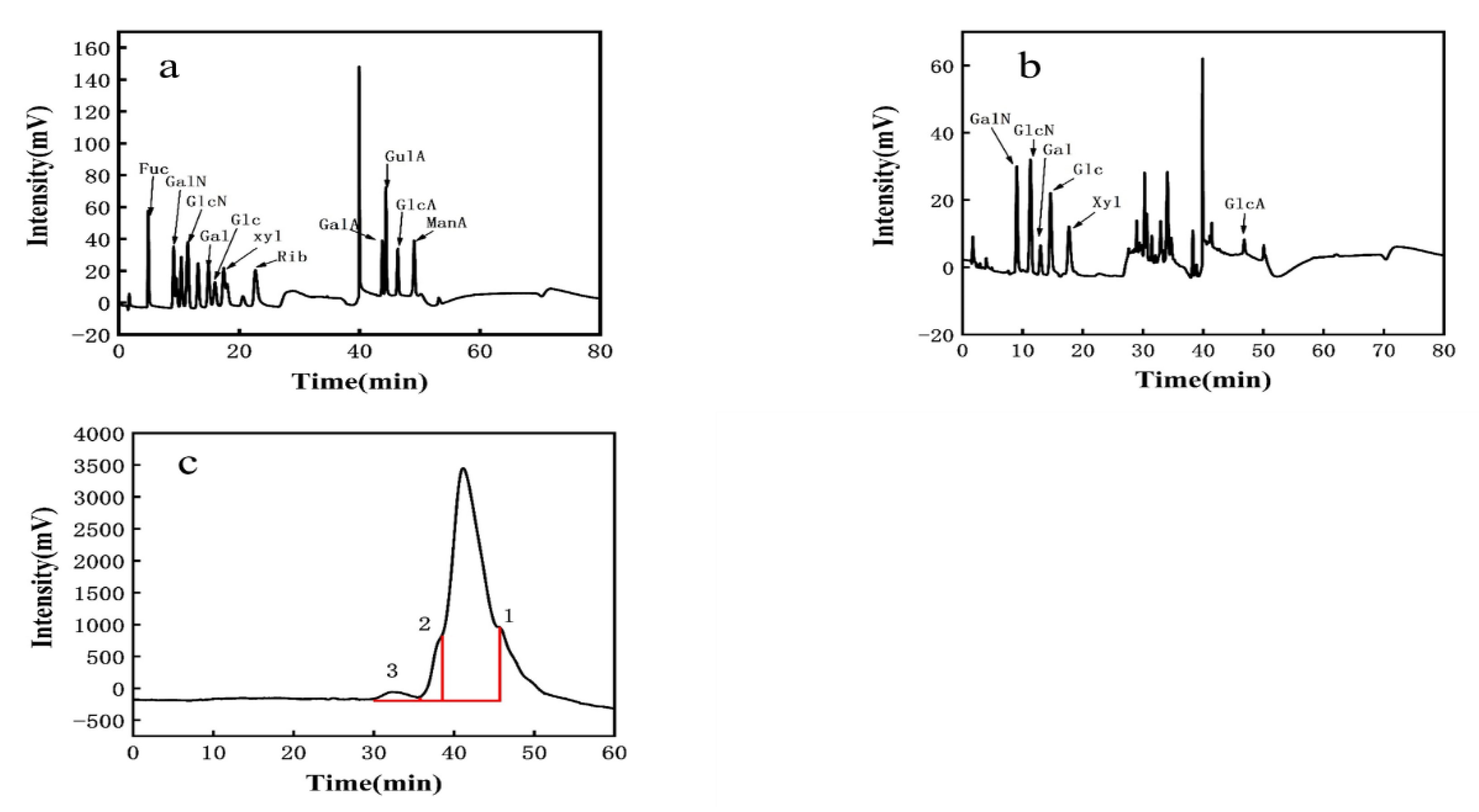
The observed elution peak was 2.0 min for sodium hydroxide and 40 min for sodium acetate. This was compared to standard monosaccharide chromatograms (Figure 2a). PW-7 EPS mainly comprised galactosamine hydrochloride, glucosamine hydrochloride, galactose, glucose, xylose, and glucuronic acid. The molar ratio was 2.804:4.217:1.978:6.304:5.435:1 (Figure 2b, Table 2). Therefore, PW-7 EPS was a heteropolysaccharide. The monosaccharide composition of LAB EPS may affect the biological activity [46,47]. The EPS of L. plantarum KX041 consisted of rhamnose, fucose, arabinose, xylose, mannose, glucose, galactose, and galacturonic acid [48]. In contrast, the EPS of L. helveticus LZ-R-5 was composed of galactose and glucose [49]. Most LAB EPS comprised glucose, galactose, and mannose among others, and a small amount of EPS contained xylose and glucuronic acid [36].
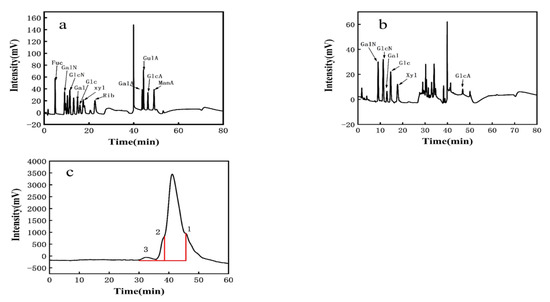
Figure 2.
(a): The HPLC chromatograms of monosaccharides standard; (b): the HPLC chromatograms of PW-7 EPS; (c): GPC analysis of EPS.
Table 2.
Analysis of monosaccharide composition of PW-7 EPS.
3.3.2. Molecular Weight of EPS
Three distinct molecular weights of EPS were determined using GPC analysis (see Figure 2c). Peak 1 corresponded to 2.33 × 104 Da, with a peak area of 93.07%. The polydispersity index (PDI, Mw/Mn) of EPS was 1.404. Peak 2 corresponded to 8.76 × 104 Da, with a peak area of 5.445% and an EPS PDI of 1.545. Peak 3 corresponded to 1.45 × 106 Da, with a peak area of 1.483% and an EPS PDI of 1.893. According to Wang et al. [50], PDI, as a measure of the molecular weight distribution of EPS, has a significant impact on the characteristics of EPS. Kieran M Lynch found that the molecular weight of LAB EPS is generally between 104–106 Da [51]. Based on peak 1, strain PW-7 could produce heteropolysaccharides, the molecular weight of which was over 104 Da.
3.4. Minimum Inhibitory Concentration (MIC)
The minimum inhibitory concentration of extracellular polysaccharide against E. coli was 40 mg/mL. The minimum inhibitory concentration against S. aureus and H. pylori was 50 mg/mL (see Table 3).
Table 3.
MIC of EPS against indicator bacteria.
3.5. Antibacterial Mechanism
3.5.1. Cell Membrane Permeability
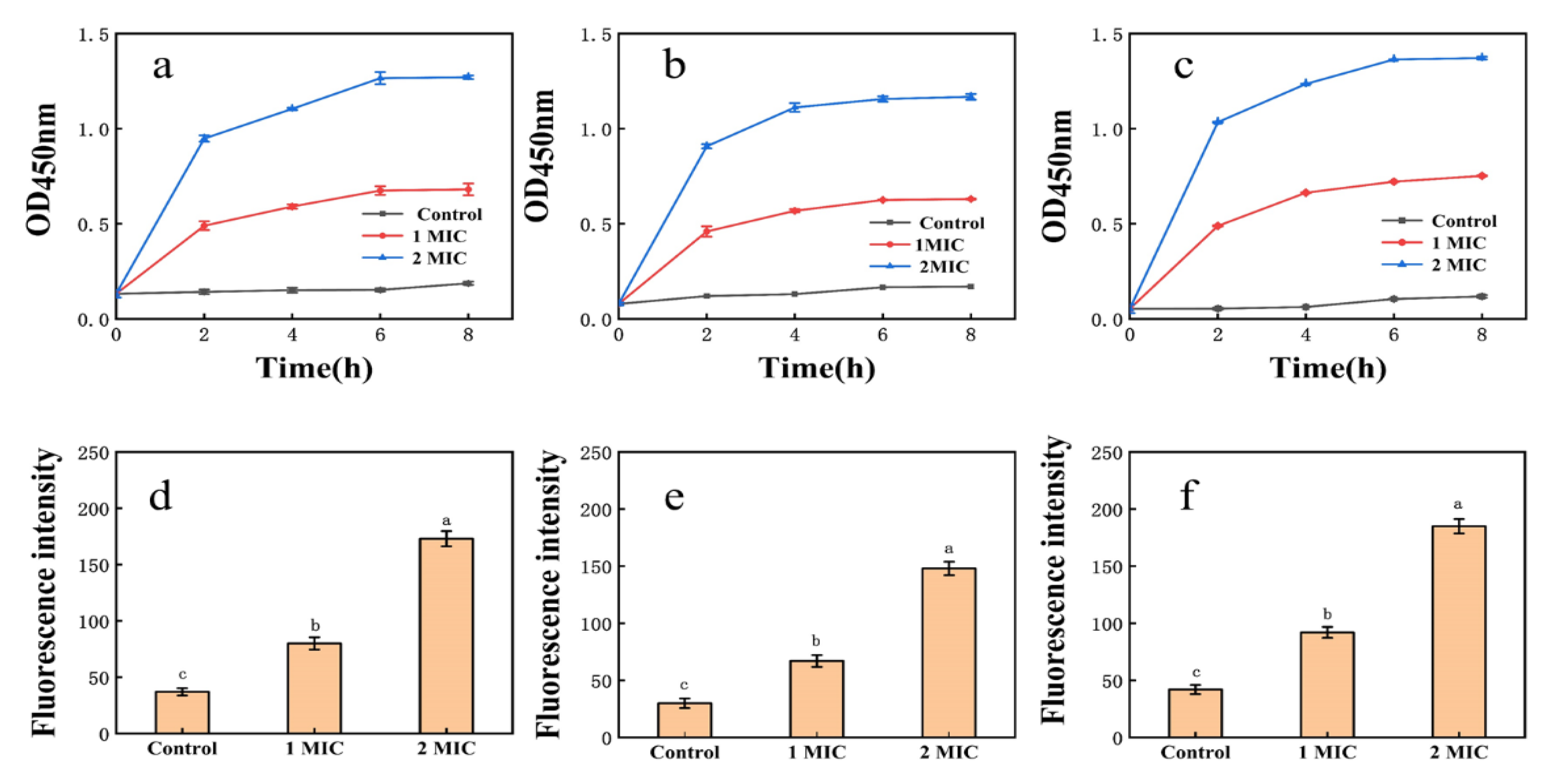
EPS affected the membrane permeability of E. coli, S. aureus, and H. pylori (Figure 3). In the control groups of E. coli, S. aureus, and H. pylori, conductivity measurements increased only slightly with time, perhaps resulting from normal bacterial lysis and cell death [52]. When the EPS concentration was greater than or equal to 1× MIC, conductivity measurements increased immediately, rising rapidly with time and polysaccharide concentration. Growth tended to be flat at 6–8 h, suggesting that EPS destroys the normal structure of bacterial cell membranes. Increased cell permeability indicates the leakage of intracellular components, particularly electrolytes such as K+, Ca2+, and Na+ [40].
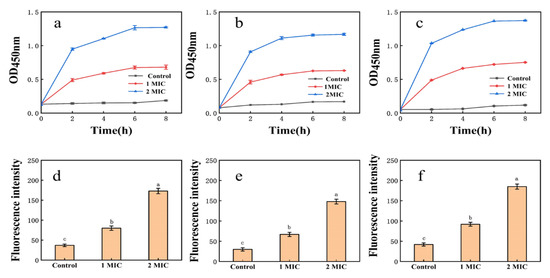
Figure 3.
(a): Effect of EPS on the membrane permeability of E. coli; (b): effect of EPS on the membrane permeability of S. aureus; (c): effect of EPS on the membrane permeability of H. pylori; (d): NPN uptake of E. coli treated with EPS; (e): NPN uptake of S. aureus treated with EPS; (f): NPN uptake of H. pylori treated with EPS. Values are expressed as the mean ± SD of triplicates. Different lowercase letters indicate statistically significant differences between means (p < 0.05) for EPS of different concentration.
3.5.2. Integrity of Cell Membranes
As EPS concentration rose, the release of cell components increased significantly (Table 4). When the concentration of EPS was greater than or equal to 1× MIC, the concentration of cell components (OD260nm), reducing sugar, and soluble protein increased with increasing EPS concentration. The leakage of nucleic acids, proteins, and other cell components resulted from a loss of membrane integrity [52]. Pradeepa et al. found that LAB EPS alters negatively-charged proteins and glycoproteins on the bacterial cell surface [53]. EPS may damage Staphylococcus aureus cells, leading to protein leakage and cell death.
Table 4.
Effects of EPS on cell components’ release of H. pylori.
3.5.3. Cell Membrane NPN Uptake
Fluorescence intensity measurements of E. coli, S. aureus, and H. pylori suspensions increased sharply after 1× MIC and 2× MIC treatment (Figure 3). Cell damage was dose-dependent, consistent with membrane integrity assays. Exposure of bacteria to EPS solution may induce structural changes in the bacterial cell membrane and concomitant functional damage. The principal component of cell membranes is phospholipid, and NPN may combine with phospholipid, resulting in significantly increased fluorescence [40].
3.5.4. Electron Microscopic Observations
In the log phase, 2× MIC EPS was reacted for 1h with indicator bacteria. SEM images indicated untreated E. coli, S. aureus, and H. pylori were regular and complete (Figure 4a–c), with normal cell morphology, an intact cell wall, and undisrupted cell membranes. In contrast, bacteria treated with EPS (Figure 4d–f) were pitted, irregular, and sunken, with altered cell morphology. Some cells were broken, with absent or ruptured membranes. Our results show EPS may seriously disrupt the cell membranes of H. pylori, resulting in intracellular content leakage [44].

Figure 4.
The SEM photography, control: (a): E. coli, (b): S. aureus, (c): H. pylori. Treated with EPS at 2× MIC: (d): E. coli, (e): S. aureus, (f): H. pylori.
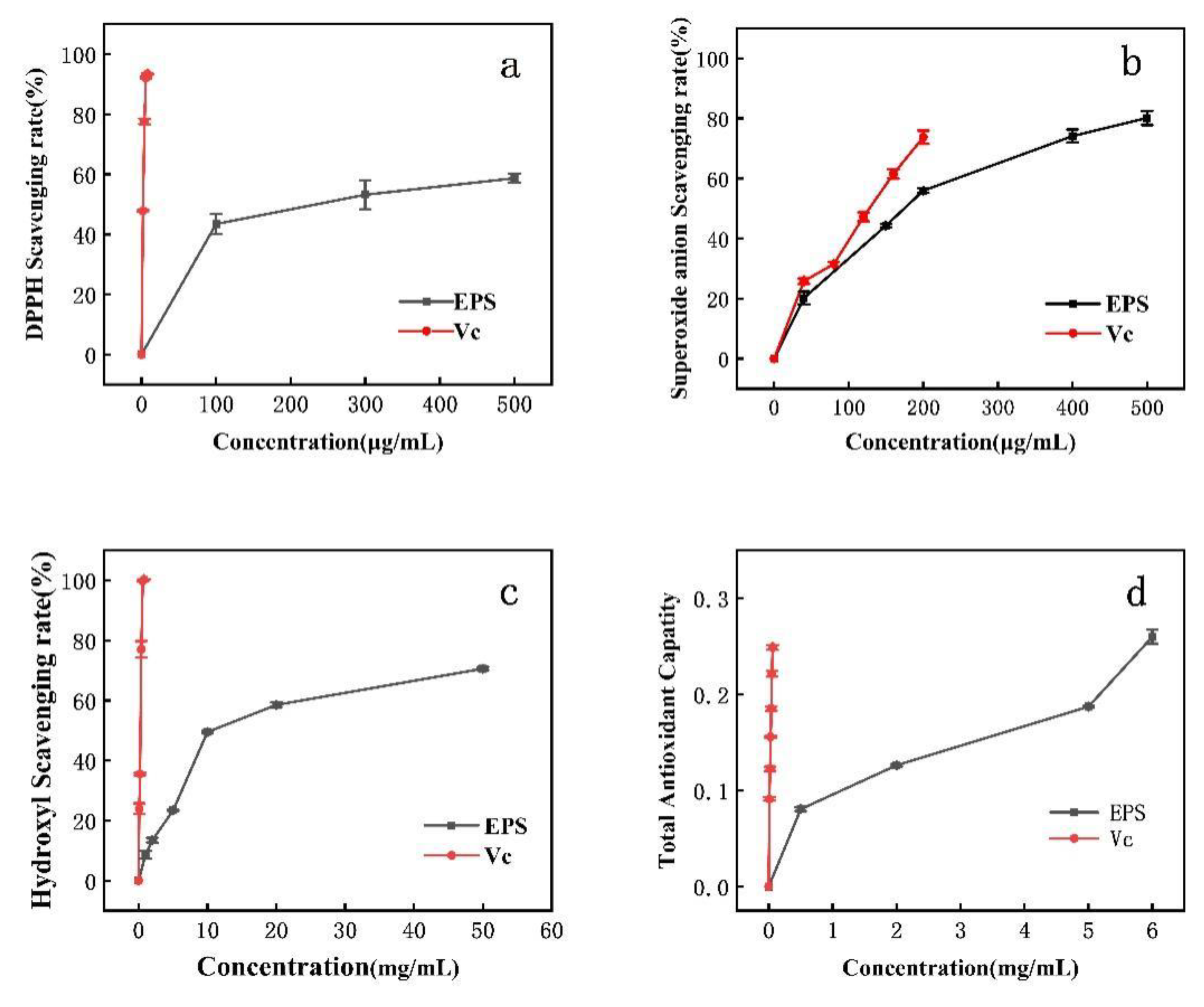
3.6. Antioxidant Activity of EPS
DPPH clearance increased with increasing EPS concentration. The IC50 for DPPH removal was 300 μg/mL (Figure 5a). DPPH is a relatively stable lipid free radical, able to inhibit the oxidation of free radicals after the addition of antioxidant. The scavenging of superoxide anion via EPS is concentration-dependent. At an EPS concentration of 180 μg/mL, the scavenging rate of superoxide anion reached the IC50. When the concentration of Lactobacillus helveticus MB 2-1 EPS was 4 mg/mL, the superoxide radical scavenging rate was 71.82%. When EPS was 6.83 mg/mL, the DPPH scavenging rate was 50% [24]. When the Lactobacillus rhamnosus EPS concentration was 4 mg/mL, the DPPH scavenging rates reached (EPS 114) 37.9% and (EPS 111) 63.4%, while the superoxide radical scavenging rates were 5.9% (EPS 115) and 29.4% (EPS 111). However, the DPPH and superoxide radical scavenging rates of PW-7 EPS were higher [43].
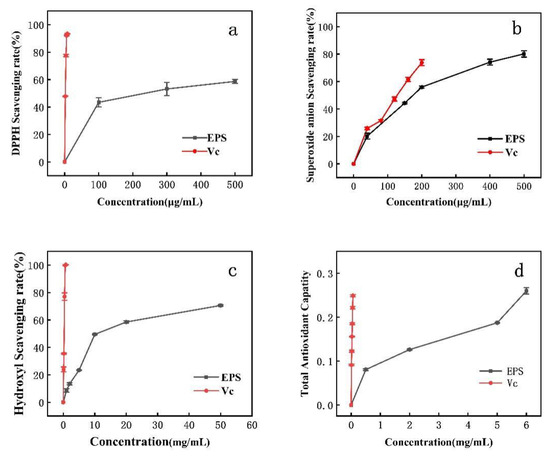
Figure 5.
Antioxidant activity of EPS. (a): DPPH radical scavenging activity; (b): superoxide radical scavenging activity; (c): hydroxyl radical scavenging activity; (d): reducing power.
3.7. Effect of Monosaccharide on Cell Membrane Permeability
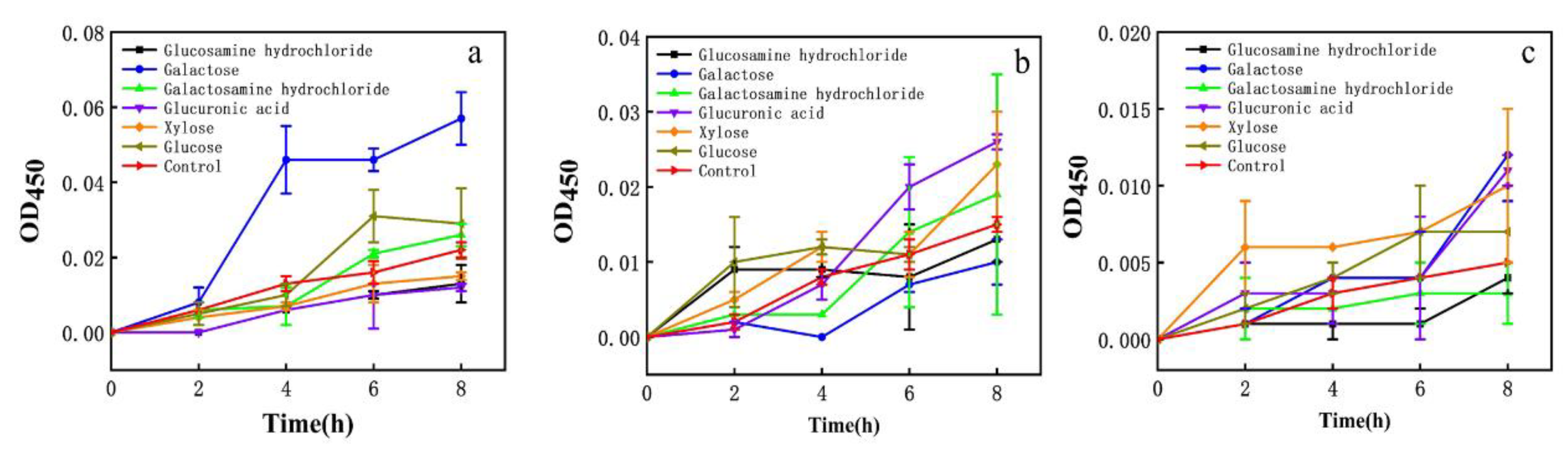
The effect on the membrane permeability of each EPS monosaccharide component was evaluated. Galactose had a greater effect on E. coli and glucuronic acid had a greater effect on S. aureus. Xylose had a stronger effect on H. pylori (Figure 6). EPS had more than 7.6% xylose.
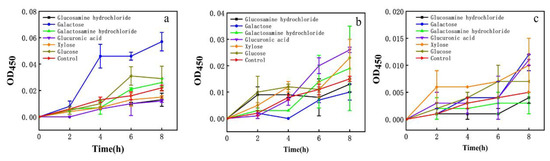
Figure 6.
Effect of different monosaccharides on cell membrane impermeability. (a) E. coli; (b) S. aureus; (c) H. pylori.
Prior work suggested EPS had an inhibitory effect on H. pylori; however, the antibacterial effect of EPS components on H. pylori was not further analyzed [24,25]. It was found that the EPS (EPSWLD and EPSMLD) produced by wild-type and mutant L. delbrueckii contained monosaccharides such as ribose, xylose, arabinose, rhamnose, fructose, glucose, mannose, and galactose, and the EPS inhibited Bacillus subtilis and S. aureus. EPS also has antioxidant capacity. When the concentration was 10 mg/mL, EPSWLD and EPSMLD have the highest DPPH activity (73.4% and 65.6%) and H2O2 scavenging activity (88.5% and 78.6%) [54]. The EPS of L. plantarum WLPL04 was composed of xylose, glucose, and galactose, and the molar ratio was 3.4:1.8:1. It can inhibit the pathogenic biofilm of Pseudomonas aeruginosa, E. coli, Salmonella typhimurium, and S. aureus [45]. Therefore, we speculated that the EPS (heteropolysaccharides) with a higher portion of xylose might be a factor to inhibit H. pylori, which paves the way for future research on inhibiting H. pylori.
4. Conclusions
To conclude, EPS is a heteropolysaccharide extracted from L. plajomi PW-7 with antibacterial activity against H. pylori, S. aureus, and E. coli. The MIC of the EPS for H. pylori, S. aureus, and E. coli was 50, 50, and 40 mg/mL, respectively. Antibacterial activity exhibited by EPS increases with increasing concentration and time. EPS exerts its antibacterial effect by disrupting the cell membranes of H. pylori, S. aureus, and E. coli. The molecular weight of EPS was approximately 2.33 × 104 Da (93.07%). The EPS from xylose may exert important effects on anti-H. pylori. EPS is also a potent antioxidant. To summarize, the EPS from L. plajomi PW-7 has clear antibacterial and anti-oxidation properties. The PW-7 strain should be useful in health care and as a food additive, along with a range of other important applications.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/foods10102453/s1, Table S1: Experiment on Removal of Hydrogen Peroxide from the Fermentation Supernatant of Strain PW-7, Table S2: Experiment on Removal of organic acid from the Fermentation Supernatant of Strain PW-7, Table S3: Antibacterial spectrum of bacteriocin of strain PW-7.
Author Contributions
J.H.: Conceptualization, methodology, writing—original draft preparation. X.T.: Data curation, resources. T.W.: Visualization, investigation. H.W.: Supervision. J.L.: Software, validation. M.L.: Writing—reviewing and editing. S.W.: Funding acquisition, project administration. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Key R&D Program of China (2018YFC0311106), the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), and the Research and Practice Innovation Program of Jiangsu (SJCX20_1321).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The interaction data used to support the findings of this study are included within the article and the supporting information file. Also, all the data used to support the findings of this study are available from the corresponding author upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Juvale, K.; Purushothaman, G.; Singh, V.; Shaik, A.; Ravi, S.; Thiruvenkatam, V.; Kirubakaran, S. Identification of selective inhibitors of Helicobacter pylori IMPDH as a targeted therapy for the infection. Sci. Rep. 2019, 9, 190. [Google Scholar] [CrossRef] [Green Version]
- Miqueleiz-Zapatero, A.; Alba-Rubio, C.; Domingo-Garcia, D.; Canton, R.; Gomez-Garcia de la Pedrosa, E.; Aznar-Cano, E.; Leiva, J.; Montes, M.; Sanchez-Romero, I.; Rodriguez-Diaz, J.C.; et al. First national survey of the diagnosis of Helicobacter pylori infection in Clinical Microbiology Laboratories in Spain. Enferm. Infecc. Microbiol. Clin. (Engl. Ed.) 2020, 38, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Ghobadi, E.; Ghanbarimasir, Z.; Emami, S. A review on the structures and biological activities of anti-Helicobacter pylori agents. Eur. J. Med. Chem. 2021, 223, 113669. [Google Scholar] [CrossRef]
- Gutierrez-Zamorano, C.; Gonzalez-Avila, M.; Diaz-Blas, G.; Smith, C.T.; Gonzalez-Correa, C.; Garcia-Cancino, A. Increased anti-Helicobacter pylori effect of the probiotic Lactobacillus fermentum UCO-979C strain encapsulated in carrageenan evaluated in gastric simulations under fasting conditions. Food Res. Int. 2019, 121, 812–816. [Google Scholar] [CrossRef]
- Karbalaei, M.; Keikha, M. Rescue effects of Lactobacillus-containing bismuth regimens after Helicobacter pylori treatment failure. New Microbes New Infect 2021, 42, 100904. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tian, F.; Liu, X.; Zhao, J.; Zhang, H.P.; Zhang, H.; Chen, W. In vitro screening of lactobacilli with antagonistic activity against Helicobacter pylori from traditionally fermented foods. J. Dairy Sci. 2010, 93, 5627–5634. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Hormazabal, P.; Arenas, A.; Serrano, C.; Pizarro, M.; Fuentes-Lopez, E.; Arnold, J.; Berger, Z.; Musleh, M.; Valladares, H.; Lanzarini, E.; et al. Prevalence of Helicobacter pylori Antimicrobial Resistance Among Chilean Patients. Arch. Med. Res. 2021, 52, 529–534. [Google Scholar] [CrossRef]
- Azizian, K.; Osquee, H.O.; Pourlak, T.; Hosseinpour, R.; Asgharzadeh, M.; Ganvarov, K.; Kafil, H.S. Genetic diversity of Lactobacillus spp. isolates from oral cavity and their probiotic and antimicrobial properties. Gene Rep. 2021, 24, 101231. [Google Scholar] [CrossRef]
- Jiang, B.; Tian, L.; Huang, X.; Liu, Z.; Jia, K.; Wei, H.; Tao, X. Characterization and antitumor activity of novel exopolysaccharide APS of Lactobacillus plantarum WLPL09 from human breast milk. Int. J. Biol. Macromol. 2020, 163, 985–995. [Google Scholar] [CrossRef]
- Xu, Y.; Cui, Y.; Yue, F.; Liu, L.; Shan, Y.; Liu, B.; Zhou, Y.; Lu, X. Exopolysaccharides produced by Lactic Acid Bacteria and Bifidobacteria: Structures, Physiochemical Functions and Applications in the Food Industry. Food Hydrocoll. 2019, 94, 475–499. [Google Scholar] [CrossRef]
- Sauer, M.; Russmayer, H.; Grabherr, R.; Peterbauer, C.K.; Marx, H. The Efficient Clade: Lactic Acid Bacteria for Industrial Chemical Production. Trends Biotechnol. 2017, 35, 756–769. [Google Scholar] [CrossRef]
- Abdullah, A.; Winaningsih, I.; Hadiyarto, A. Lactic Acid Fermentation from Durian Seeds (Durio zibethinus Murr.) Using Lactobacillus plantarum. In IOP Conference Series: Materials Science and Engineering; IOP Publishing: Bristol, UK, 2021; Volume 1053, p. 012032. [Google Scholar]
- Yu, M.; Zhang, R.; Ni, P.; Chen, S.; Duan, G. Efficacy of Lactobacillus-supplemented triple therapy for H. pylori eradication: A meta-analysis of randomized controlled trials. PLoS ONE 2019, 14, e0223309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poonyam, P.; Chotivitayatarakorn, P.; Vilaichone, R.K. High Effective of 14-Day High-Dose PPI- Bismuth-Containing Quadruple Therapy with Probiotics Supplement for Helicobacter Pylori Eradication: A Double Blinded-Randomized Placebo-Controlled Study. Asian Pac. J. Cancer Prev. 2019, 20, 2859–2864. [Google Scholar] [CrossRef] [PubMed]
- García, A.; Navarro, K.; Sanhueza, E.; Pineda, S.; Pastene, E.; Quezada, M.; Henríquez, K.; Karlyshev, A.; Villena, J.; González, C. Characterization of Lactobacillus fermentum UCO-979C, a probiotic strain with a potent anti-Helicobacter pylori activity. Electron. J. Biotechnol. 2017, 25, 75–83. [Google Scholar] [CrossRef]
- Wu, D.; Cao, M.; Zhou, J.; Yan, S.; Peng, J.; Yu, Z.; Zhang, A.; Wu, J.; Yan, X.; Zhao, J. Lactobacillus casei T1 from kurut against Helicobacter pylori-induced inflammation and the gut microbial disorder. J. Funct. Foods 2021, 85, 104611. [Google Scholar] [CrossRef]
- Zhang, X.; Ali Esmail, G.; Fahad Alzeer, A.; Valan Arasu, M.; Vijayaraghavan, P.; Choon Choi, K.; Abdullah Al-Dhabi, N. Probiotic characteristics of Lactobacillus strains isolated from cheese and their antibacterial properties against gastrointestinal tract pathogens. Saudi J. Biol. Sci. 2020, 27, 3505–3513. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.X.; Fang, H.Y.; Yang, H.B.; Tien, N.Y.; Wang, M.C.; Wu, J.J. Lactobacillus pentosus strain LPS16 produces lactic acid, inhibiting multidrug-resistant Helicobacter pylori. J. Microbiol. Immunol. Infect. 2016, 49, 168–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.W.; Xiang, Y.Z.; Zhang, M.; Jiang, Y.H.; Zhang, Q.L. A novel bacteriocin from Lactobacillus salivarius against Staphylococcus aureus: Isolation, purification, identification, antibacterial and antibiofilm activity. LWT-Food Sci. Technol. 2020, 140, 110826. [Google Scholar] [CrossRef]
- Ale, E.C.; Rojas, M.F.; Reinheimer, J.A.; Binetti, A.G. Lactobacillus fermentum: Could EPS production ability be responsible for functional properties? Food Microbiol. 2020, 90, 103465. [Google Scholar] [CrossRef]
- Rahbar Saadat, Y.; Yari Khosroushahi, A.; Pourghassem Gargari, B. A comprehensive review of anticancer, immunomodulatory and health beneficial effects of the lactic acid bacteria exopolysaccharides. Carbohydr. Polym. 2019, 217, 79–89. [Google Scholar] [CrossRef]
- Ayyash, M.; Abu-Jdayil, B.; Itsaranuwat, P.; Galiwango, E.; Tamiello-Rosa, C.; Abdullah, H.; Esposito, G.; Hunashal, Y.; Obaid, R.S.; Hamed, F. Characterization, bioactivities, and rheological properties of exopolysaccharide produced by novel probiotic Lactobacillus plantarum C70 isolated from camel milk. Int. J. Biol. Macromol. 2020, 144, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Ryan, K.A.; Daly, P.; Li, Y.; Hooton, C.; O’Toole, P.W. Strain-specific inhibition of Helicobacter pylori by Lactobacillus salivarius and other lactobacilli. J. Antimicrob. Chemother. 2008, 61, 831–834. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Luo, J.; Xue, R.Y.; Guo, L.; Nie, L.; Li, S.; Ji, L.; Ma, C.J.; Chen, D.Q.; Miao, K.; et al. The mucosal adjuvant effect of plant polysaccharides for induction of protective immunity against Helicobacter pylori infection. Vaccine 2019, 37, 1053–1061. [Google Scholar] [CrossRef]
- Garcia-Castillo, V.; Marcial, G.; Albarracin, L.; Tomokiyo, M.; Clua, P.; Takahashi, H.; Kitazawa, H.; Garcia-Cancino, A.; Villena, J. The Exopolysaccharide of Lactobacillus fermentum UCO-979C Is Partially Involved in Its Immunomodulatory Effect and Its Ability to Improve the Resistance against Helicobacter pylori Infection. Microorganisms 2020, 8, 479. [Google Scholar] [CrossRef] [Green Version]
- Kšonžeková, P.; Bystrický, P.; Vlčková, S.; Pätoprstý, V.; Pulzová, L.; Mudroňová, D.; Kubašková, T.; Csank, T.; Tkáčiková, Ľ. Exopolysaccharides of Lactobacillus reuteri: Their influence on adherence of E. coli to epithelial cells and inflammatory response. Carbohydr. Polym. 2016, 141, 10–19. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, X.; Jiang, Y.; Zhao, W.; Guo, T.; Cao, Y.; Teng, J.; Hao, X.; Zhao, J.; Yang, Z. Antioxidant status and gut microbiota change in an aging mouse model as influenced by exopolysaccharide produced by Lactobacillus plantarum YW11 isolated from Tibetan kefir. J. Dairy Sci. 2017, 100, 6025–6041. [Google Scholar] [CrossRef] [PubMed]
- Chen, O.; Hong, Y.; Ma, J.; Deng, L.; Yi, L.; Zeng, K. Screening lactic acid bacteria from pickle and cured meat as biocontrol agents of Penicillium digitatum on citrus fruit. Biol. Control 2021, 158, 104606. [Google Scholar] [CrossRef]
- Pan, M.; Wan, C.; Xie, Q.; Huang, R.; Tao, X.; Shah, N.P.; Wei, H. Changes in gastric microbiota induced by Helicobacter pylori infection and preventive effects of Lactobacillus plantarum ZDY 2013 against such infection. J. Dairy Sci. 2016, 99, 970–981. [Google Scholar] [CrossRef]
- Lorn, D.; Nguyen, T.K.; Ho, P.H.; Tan, R.; Licandro, H.; Wache, Y. Screening of lactic acid bacteria for their potential use as aromatic starters in fermented vegetables. Int. J. Food Microbiol. 2021, 350, 109242. [Google Scholar] [CrossRef]
- Snow, M.J.; Manley-Harris, M. On the nature of non-peroxide antibacterial activity in New Zealand manuka honey. Food Chem. 2004, 84, 145–147. [Google Scholar] [CrossRef]
- Le, V.T.; Leelakriangsak, M.; Lee, S.W.; Panphon, S.; Utispan, K.; Koontongkaew, S. Characterization and safety evaluation of partially purified bacteriocin produced by Escherichia coli E isolated from fermented pineapple Ananas comosus (L.) Merr. Braz. J. Microbiol. 2019, 50, 33–42. [Google Scholar] [CrossRef]
- Ndlovu, B.; Schoeman, H.; Franz, C.M.; du Toit, M. Screening, identification and characterization of bacteriocins produced by wine-isolated LAB strains. J. Appl. Microbiol. 2015, 118, 1007–1022. [Google Scholar] [CrossRef]
- Mahdhi, A.; Leban, N.; Chakroun, I.; Chaouch, M.A.; Hafsa, J.; Fdhila, K.; Mahdouani, K.; Majdoub, H. Extracellular polysaccharide derived from potential probiotic strain with antioxidant and antibacterial activities as a prebiotic agent to control pathogenic bacterial biofilm formation. Microb. Pathog. 2017, 109, 214–220. [Google Scholar] [CrossRef]
- Bomfim, V.B.; Pereira Lopes Neto, J.H.; Leite, K.S.; de Andrade Vieira, É.; Iacomini, M.; Silva, C.M.; Olbrich dos Santos, K.M.; Cardarelli, H.R. Partial characterization and antioxidant activity of exopolysaccharides produced by Lactobacillus plantarum CNPC003. LWT 2020, 127, 109349. [Google Scholar] [CrossRef]
- Xu, Y.; Cui, Y.; Wang, X.; Yue, F.; Shan, Y.; Liu, B.; Zhou, Y.; Yi, Y.; Lü, X. Purification, characterization and bioactivity of exopolysaccharides produced by Lactobacillus plantarum KX041. Int. J. Biol. Macromol. 2019, 128, 480–492. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Kim, J.H.; Lee, N.K.; Paik, H.D. Inhibitory effects of Lactobacillus brevis KU15153 against Streptococcus mutans KCTC 5316 causing dental caries. Microb. Pathog. 2021, 157, 104938. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Z.; Chen, J.; Zhou, Q.; Wang, X.; Shan, Y.; Yi, Y.; Liu, B.; Zhou, Y.; Lu, X. Purification, characterization, and mode of action of a novel bacteriocin BM173 from Lactobacillus crustorum MN047 and its effect on biofilm formation of Escherichia coli and Staphylococcus aureus. J. Dairy Sci. 2021, 104, 1474–1483. [Google Scholar] [CrossRef] [PubMed]
- Ju, H.; Chen, H.; Xiang, A.; Wang, Y.; Yue, T.; Yuan, Y. Identification and characterization of Lactobacillus paracasei strain MRS-4 antibacterial activity against Alicyclobacillus acidoterrestris. LWT 2021, 150, 111991. [Google Scholar] [CrossRef]
- Zhang, L.-L.; Zhang, L.-F.; Hu, Q.-P.; Hao, D.-L.; Xu, J.-G. Chemical composition, antibacterial activity of Cyperus rotundus rhizomes essential oil against Staphylococcus aureus via membrane disruption and apoptosis pathway. Food Control 2017, 80, 290–296. [Google Scholar] [CrossRef]
- Xing, K.; Chen, X.G.; Kong, M.; Liu, C.S.; Cha, D.S.; Park, H.J. Effect of oleoyl-chitosan nanoparticles as a novel antibacterial dispersion system on viability, membrane permeability and cell morphology of Escherichia coli and Staphylococcus aureus. Carbohydr. Polym. 2009, 76, 17–22. [Google Scholar] [CrossRef]
- Dong, D.; Wang, X.; Deng, T.; Ning, Z.; Tian, X.; Zu, H.; Ding, Y.; Wang, C.; Wang, S.; Lyu, M. A novel dextranase gene from the marine bacterium Bacillus aquimaris S5 and its expression and characteristics. FEMS Microbiol. Lett. 2021, 368, fnab007. [Google Scholar] [CrossRef] [PubMed]
- Riaz Rajoka, M.S.; Jin, M.; Haobin, Z.; Li, Q.; Shao, D.; Jiang, C.; Huang, Q.; Yang, H.; Shi, J.; Hussain, N. Functional characterization and biotechnological potential of exopolysaccharide produced by Lactobacillus rhamnosus strains isolated from human breast milk. LWT 2018, 89, 638–647. [Google Scholar] [CrossRef]
- Zhang, N.; Zhao, Y.; Shi, Y.; Chen, R.; Fu, X.; Zhao, Y. Polypeptides extracted from Eupolyphaga sinensis walker via enzymic digestion alleviate UV radiation-induced skin photoaging. Biomed. Pharmacother. 2019, 112, 108636. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, Z.; Qiu, L.; Zhang, F.; Xu, X.; Wei, H.; Tao, X. Characterization and bioactivities of the exopolysaccharide from a probiotic strain of Lactobacillus plantarum WLPL04. J. Dairy Sci. 2017, 100, 6895–6905. [Google Scholar] [CrossRef]
- Caggianiello, G.; Kleerebezem, M.; Spano, G. Exopolysaccharides produced by lactic acid bacteria: From health-promoting benefits to stress tolerance mechanisms. Appl. Microbiol. Biotechnol. 2016, 100, 3877–3886. [Google Scholar] [CrossRef]
- Das, D.; Baruah, R.; Goyal, A. A food additive with prebiotic properties of an alpha-d-glucan from lactobacillus plantarum DM5. Int. J. Biol. Macromol. 2014, 69, 20–26. [Google Scholar] [CrossRef]
- Do, T.B.T.; Tran, T.A.L.; Tran, T.V.T.; Le, T.H.; Jayasena, V.; Nguyen, T.H.C.; Nguyen, C.C.; Kim, S.Y.; Le, Q.V. Novel Exopolysaccharide Produced from Fermented Bamboo Shoot-Isolated Lactobacillus Fermentum. Polymers 2020, 12, 1531. [Google Scholar] [CrossRef]
- You, X.; Li, Z.; Ma, K.; Zhang, C.; Chen, X.; Wang, G.; Yang, L.; Dong, M.; Rui, X.; Zhang, Q.; et al. Structural characterization and immunomodulatory activity of an exopolysaccharide produced by Lactobacillus helveticus LZ-R-5. Carbohydr. Polym. 2020, 235, 115977. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, X.; Tian, Z.; Yang, Y.; Yang, Z. Characterization of an exopolysaccharide produced by Lactobacillus plantarum YW11 isolated from Tibet Kefir. Carbohydr. Polym. 2015, 125, 16–25. [Google Scholar] [CrossRef]
- Lynch, K.M.; Zannini, E.; Coffey, A.; Arendt, E.K. Lactic Acid Bacteria Exopolysaccharides in Foods and Beverages: Isolation, Properties, Characterization, and Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 155–176. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Wang, Y.; Jiang, P.; Quek, S. Antibacterial activity and mechanism of cinnamon essential oil against Escherichia coli and Staphylococcus aureus. Food Control 2016, 59, 282–289. [Google Scholar] [CrossRef]
- Pradeepa; Shetty, A.D.; Matthews, K.; Hegde, A.R.; Akshatha, B.; Mathias, A.B.; Mutalik, S.; Vidya, S.M. Multidrug resistant pathogenic bacterial biofilm inhibition by Lactobacillus plantarum exopolysaccharide. Bioact. Carbohydr. Diet. Fibre 2016, 8, 7–14. [Google Scholar] [CrossRef]
- Adebayo-Tayo, B.; Fashogbon, R. In vitro antioxidant, antibacterial, in vivo immunomodulatory, antitumor and hematological potential of exopolysaccharide produced by wild type and mutant Lactobacillus delbureckii subsp. bulgaricus. Heliyon 2020, 6, e03268. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).