1. Introduction
Gold compounds have a long history in medicinal chemistry. The thiophilic nature of gold suggests sulfur-rich proteins, such as thioredoxin, as reasonable cellular targets for gold action [
1]. Au(III) is quite labile in its chemistry and is susceptible to reduction to Au(I) [
1]. Apart from the inherent difficulty in understanding speciation of an Au(III) complex in a biological medium, nonselective interactions are also highly likely. This is especially true when highly nucleophilic sulfur sites in proteins are considered as viable targets. Formally, substitution-inert coordination spheres, such as MN
4 or MN
3L (where M = Pt, Au and N
4 may be an N
3 chelate such as diethylenetriamine (dien) and L a purine or pyrimidine ligand), have been explored by our group for greater specificity with biomolecules [
2]. In principle, we suggested that highly nucleophilic cysteines of biomolecules can be targeted using weak nucleophiles, such as platinum-nucleobase PtN
4 complexes for selective peptide reactions [
2]. Although Au(III) is more chemically reactive than Pt(II), the same principle applies [
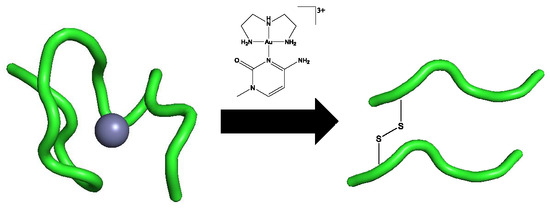
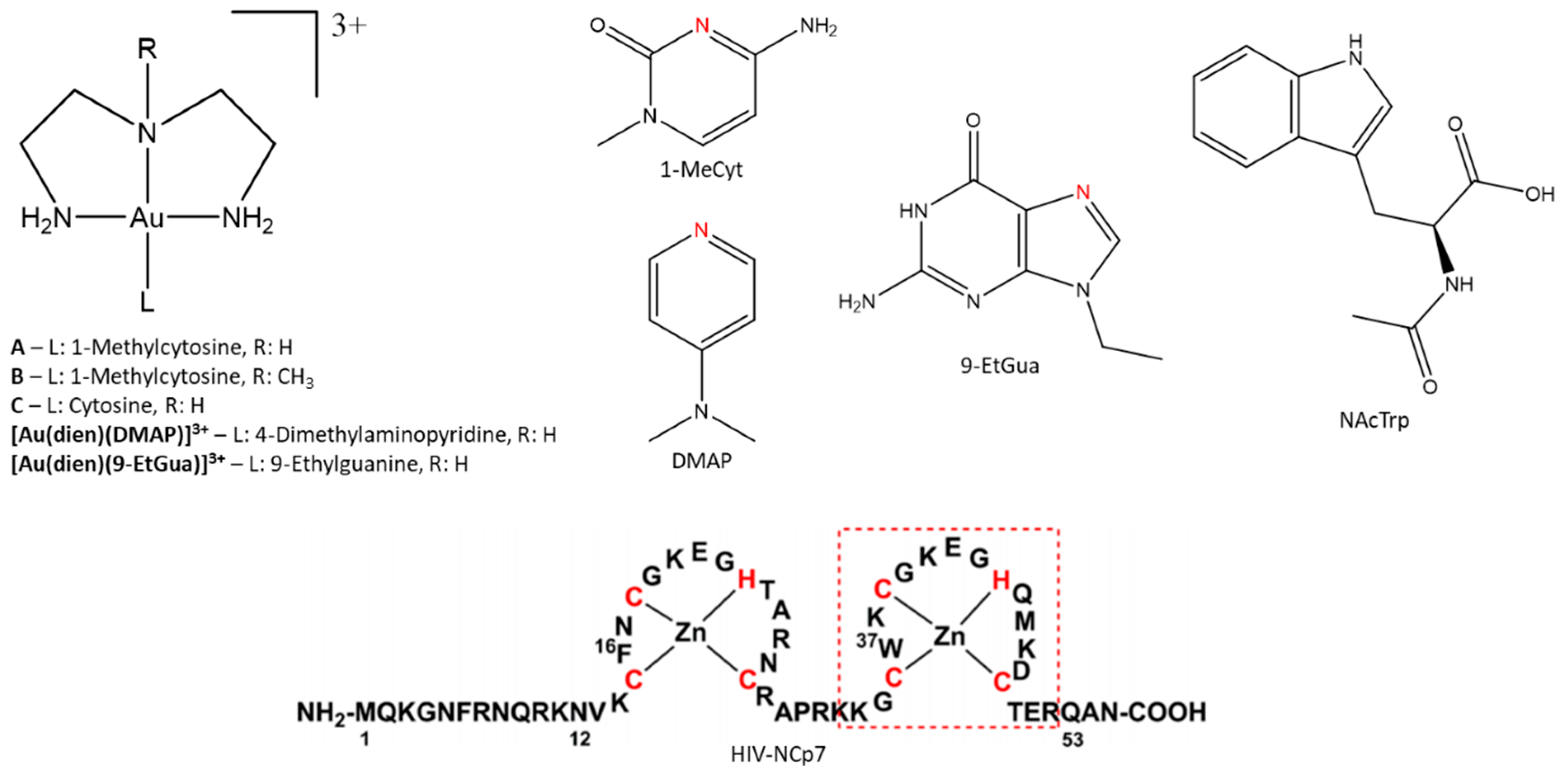
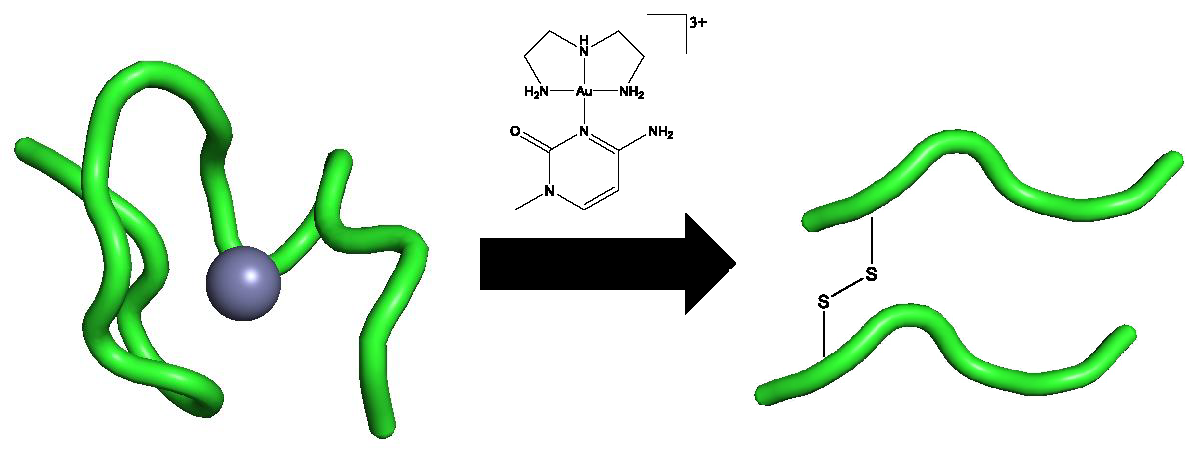
3]. A complementary approach to enhance selectivity in metal ion-biomolecule interactions involves using an inherent property of the protein along with appropriate design of the small molecule. A two-stage approach to selectivity can then be envisaged: Molecular recognition followed by “fixation”, where the inorganic moiety forms covalent linkages to the protein of interest. In this contribution, we summarize how this approach might work using formally substitution-inert platinum-metal nucleobase complexes and a tryptophan-containing zinc finger peptide from the HIV nucleocapsid protein HIVNCp7. Specifically, we describe the synthesis and properties of Au(III)-1-MeCytosine complexes and their interactions with the C-terminal NCp7-F2 targeting the essential tryptophan of the protein.
Zinc binding proteins are one of the most common types of metalloprotein and make up as much as 10% of proteins found in humans [
4]. In these proteins, zinc can serve a catalytic role, a structural role, and even a recently discovered inhibitory role [
5,
6]. In zinc-finger proteins, the zinc serves a structural role, binding in a tetrahedral geometry to four surrounding amino acids [
7]. These amino acids are generally a combination of cysteines and histidines, and take the forms Cys
2His
2, Cys
2HisCys (Cys
3His), or Cys
4. The modification of the zinc-coordinating residues can have a dramatic effect on protein function [
8]. The HIV nucleocapsid protein (NCp7), a 55 amino acid protein that contains two Cys
3His zinc fingers (
Figure 1), has been identified as a potential target for chemical intervention through the modification of these structural elements. It was previously shown that alteration of the zinc coordination sphere significantly hinders viral replication [
5]. NCp7 interacts with its polynucleotide substrate, viral RNA, through guanine stacking with both W37 on the second C-terminal zinc finger motif, as well as through the phenylalanine (F16) located on the first zinc finger [
5,
9].
NCp7 recognition by small molecules can be optimized through a noncovalent π–π stacking interaction between the “essential” tryptophan residue and metallated nucleobases, analogous to the RNA(DNA) interaction. Previously reported gold (III) complexes based on [Au(dien)L]
3+ (dien = diethylenetriamine; L =
N-heterocycle) were investigated for their ability to stabilize the Au(III) oxidation state, as well as their ability to eject zinc from zinc-finger proteins [
10,
11,
12,
13]. In the case of coordination complexes, this interaction brings the metal center in close contact with the highly nucleophilic zinc-bound cysteine residues, allowing for a secondary electrophilic attack by the metal center, resulting in zinc ejection [
2,
10]. Changing the
N-heterocyclic ligand can be used to fine-tune the electronic properties of the metal center in order to impart higher selectivity for the protein of interest. To this end, the [Au(dien)L]
3+ coordination sphere was studied with the previously uninvestigated ligand 1-methylcytosine (1-MeCyt). Compared to the
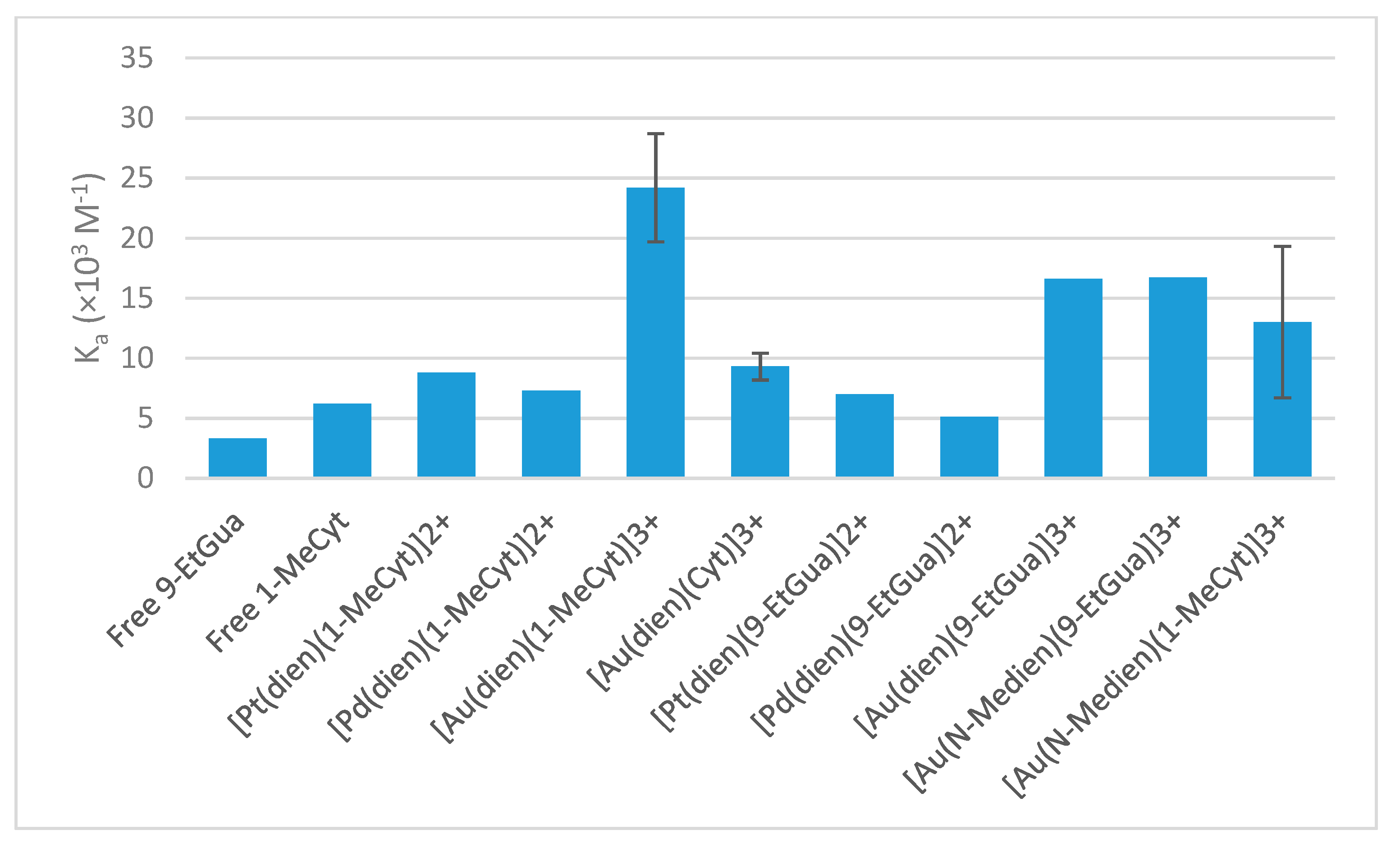
N-heterocycles 4-Dimethylaminopyridine (DMAP) and 9-Etylguanine (9-EtGua), the free ligand 1-MeCyt has a higher association constant with tryptophan [
14]. Additionally, 1-MeCyt has an intermediate basicity (pK
a = 4.45) relative to DMAP (pK
a = 9.1) and 9-EtGua (pK
a(N7) = 2.7) [
15]. Previously, basicity of the heterocycle was thought to be proportional to association constant in the [Au(dien)(
N-heterocycle)]
3+ motif [
10]. The [Au(dien)(1-MeCyt)]
3+ is, to our knowledge, the first 1-MeCytosine compound of Au(III) to be synthesized, and it completes the series of Pt(II) and Pd(II) complexes [M(dien)L]
n+ (L = 9-EtGua, 1-MeCyt) studied for their isostructural and isoelectronic similarities [
14,
16,
17]. We report the synthesis of the [Au(dien)(1-MeCyt)]
3+ (
A), [Au(
N-Medien)(1-MeCyt)]
3+ (
B), and [Au(dien)(Cyt)]
3+ (
C) (
Figure 1). The associative interaction with
N-Acetyl-L-tryptophan (NAcTrp) as a model for the recognition site of NCp7 (W37) was probed through the use of a fluorescence quenching assay. Additionally, the interactions with the C-terminal zinc finger of NCp7 (
Figure 1) were investigated by mass spectrometry and circular dichroism spectroscopy.
3. Discussion
The synthesis of complexes A and B serves to complete the series of complexes of the form [M(dien)(1-MeCyt)]n+, in which the Pt(II) and Pd(II) complexes, both isoelectronic and isostructural to the Au(III) complexes investigated in this work, had been previously investigated. However, the Au(III) analog had remained unreported up to this point.
Tryptophan is present on the C-terminal zinc-finger of NCp7 and plays an essential role in protein-DNA/RNA recognition, which occurs through a π–π stacking interaction between the aromatic tryptophan (or in the case of the full peptide, phenylalanine) residues and the nucleobases in DNA/RNA [
14,
19]. Previously, it was shown that association with free
N-Acetyl-L-tryptophan (NAcTrp) is a good indicator of a compound’s affinity for the full protein. The fluorescent emission of NAcTrp can be quenched by a complex that interacts with NAcTrp in a π–π stacking interaction, and the degree to which the fluorescent emission is quenched is directly related to the strength of the interaction [
3,
10,
11,
12,
14,
15].
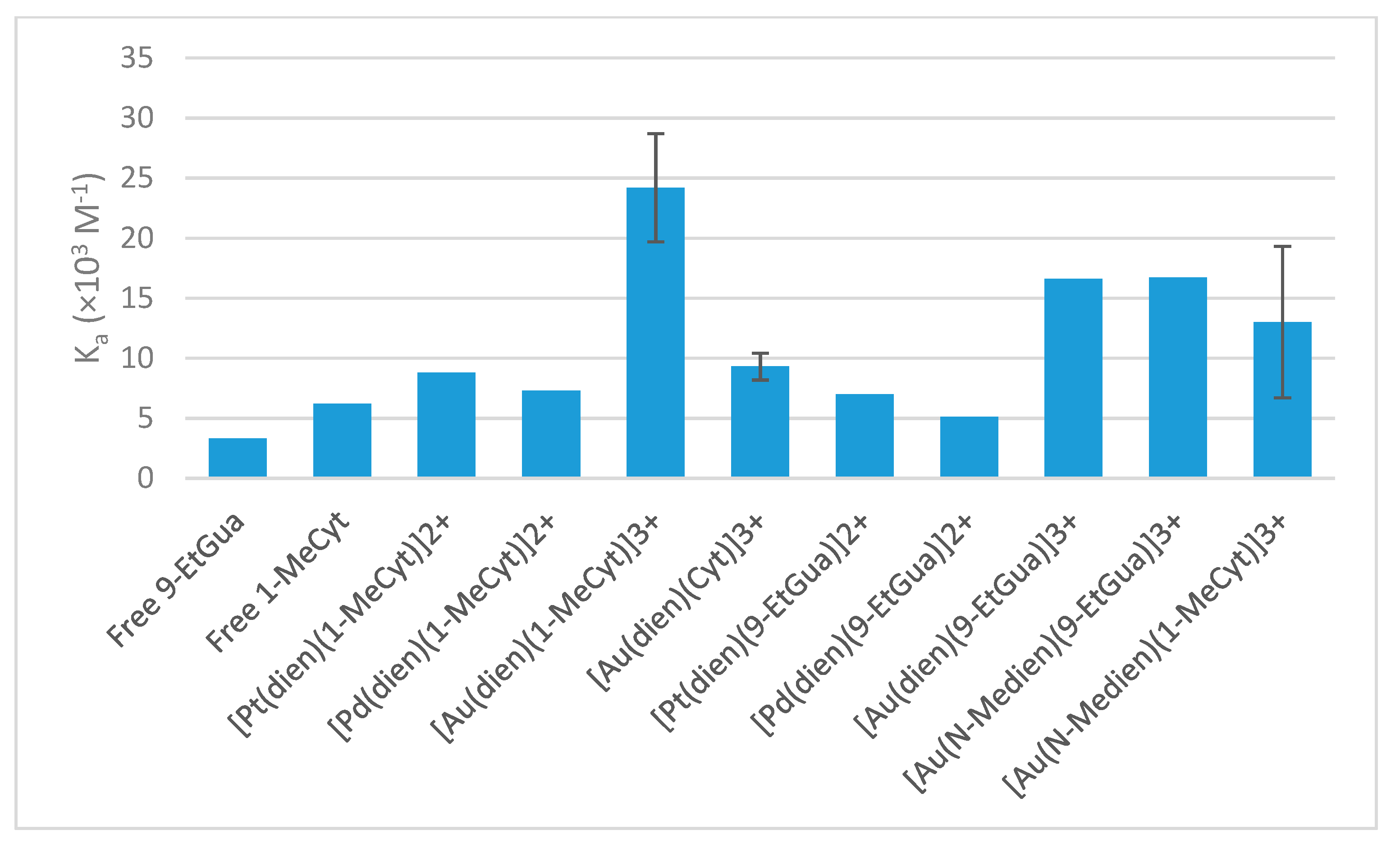
Compared to the Pt(II) and Pd(II) analogs, the Au(III) complexes show a higher affinity for NAcTrp, consistent with the previous literature [
5,
20]. The lower affinity of
B for NAcTrp than the DMAP analog and similar affinity compared to the 9-EtGua analog is surprising given the intermediate basicity of 1-MeCyt. The result for
A is as expected, as the basicity of free 1-MeCyt is intermediate between that of DMAP and 9-EtGua, but the results obtained for
B and
C show that there are factors other than the basicity that influence the associative interaction. Therefore, it is likely that the
N-methyl group present on the chelating ligand in
B causes a lower association constant for the π–π stacking interaction with NAcTrp than in
A. This shows that the modulation of the electronic environment of the metal center can influence the interactions between the complex and the protein recognition element. However, this reduction upon methylation of the chelating ligand is only observed to a lesser extent in the 9-EtGua analog.
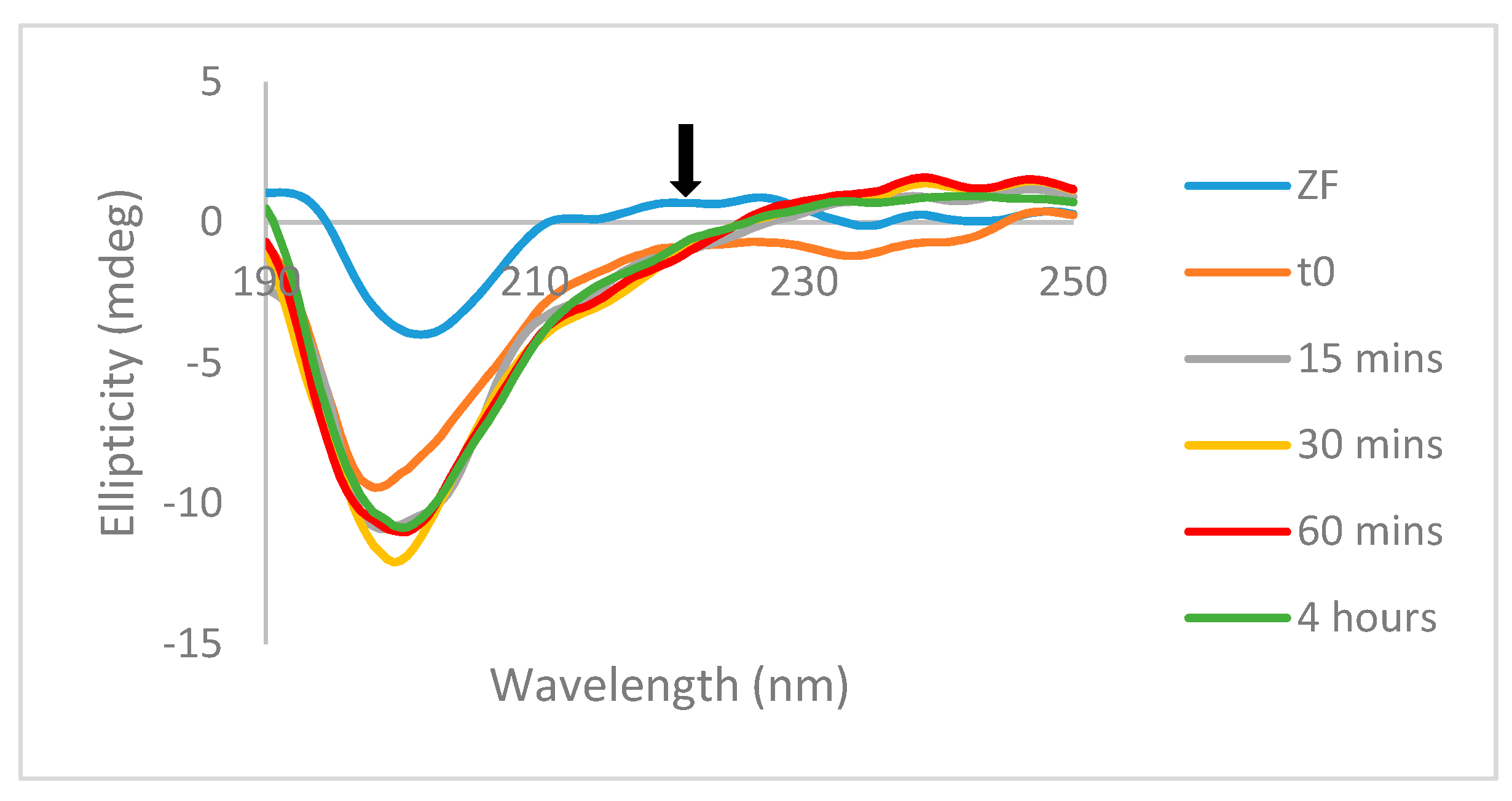
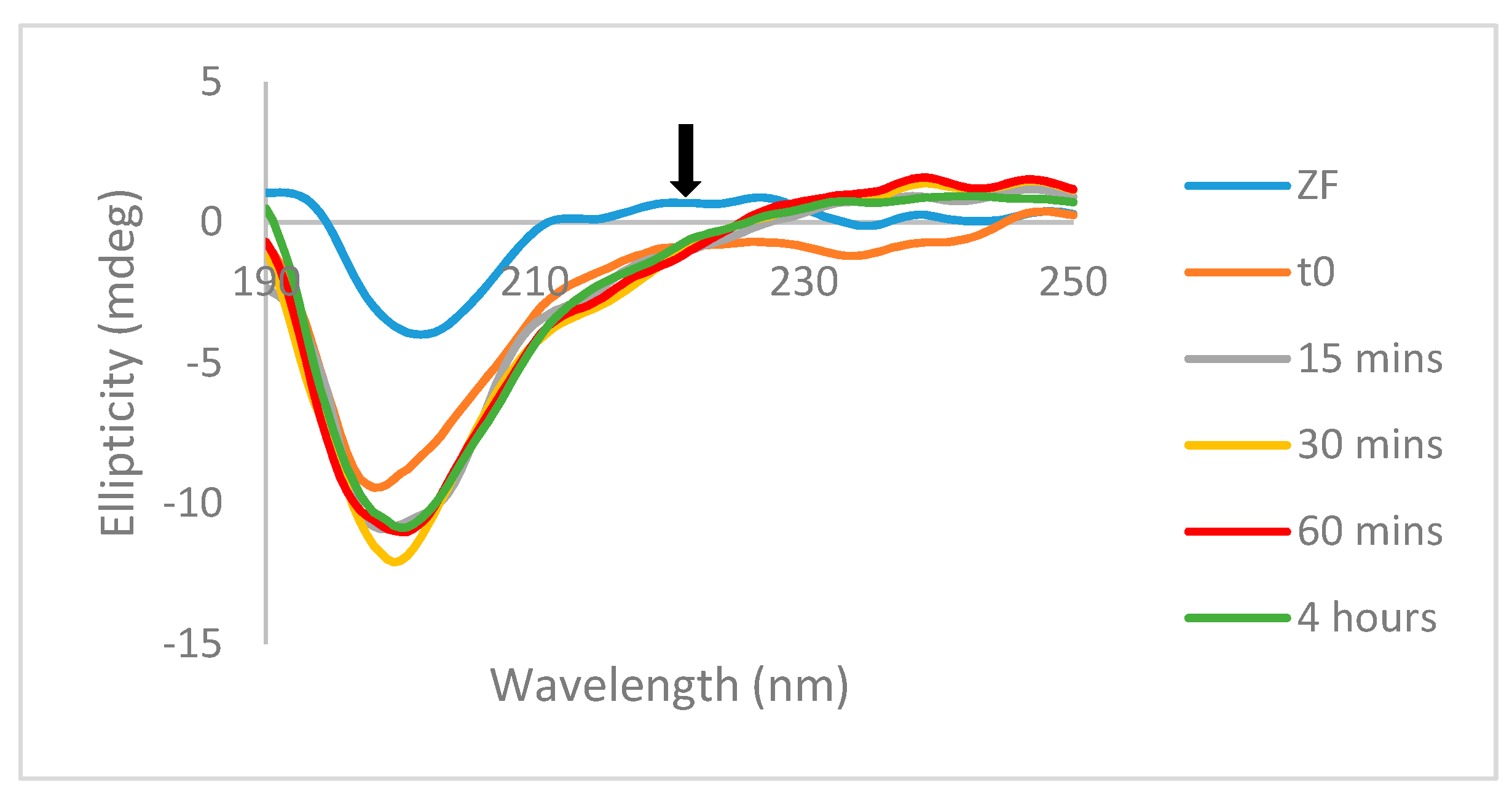
Mass spectrometry studies with the previously reported Au(III) analogs showed that the formation of random coil observed by CD was accompanied by the formation of “gold fingers,” in which one or more atoms of gold replaced the zinc in the zinc-finger structure upon zinc ejection [
10]. The observation of oxidized apopeptide is indicative of disulfide bond formation between cysteine molecules of the same peptide, while the observation of oxidized dimerized apopeptide is indicative of disulfide bond formation between cysteine residues of different protein molecules. These interactions would likely be facilitated by the reduction of the metal center from Au(III) to Au(I)/Au(0). This interaction is in contrast to the interactions observed in the Pt(II) analogs, in which reduction and incorporation of the Pt(II) metal center is not seen. Instead, the metal center is observed with the ligands intact and with the zinc still incorporated [
3,
5,
20]. Like previously reported aurated DMAP and 9-EtGua analogs, but unlike the platinated analogs, peaks representing the unreacted zinc-finger protein are not seen at the initial time point, highlighting the increased reactivity of the Au(III) metal center. The incorporation of four equivalents of gold was thought to be due to a difference in the redox properties of
A and
B when compared to the previously studied Au(III) analogs. Therefore, CV studies were carried out in order to assess these differences.
CV showed that it is significantly easier to reduce cytosine and thymine than it is to reduce adenine and guanine [
21]. In the context of interaction with zinc-finger peptides, this reduction would be accompanied by disulfide bond formation. In this case, the greater ease of reduction may cause the occurrence of a charge transfer mechanism, in which the disulfide bond formation occurs across two equivalents of the peptide, forming the oxidized dimerized peptide uniquely observed in the case of
A and
B, in contrast to the simple oxidation of one equivalent of zinc finger. Whereas this latter oxidation is easily understood with reduction of Au(III) to Au(I) and concomitant oxidation of two Zn-bound cysteines, the first step in peptide dimerization may be an electron transfer between the 1-MeCyt ligand to the tryptophan ligand. This suggestion would explain why similar products would not be seen in the isostructural Pt(II) complexes, as the platinum center is not reduced upon interaction with zinc-finger peptides, while a charge transfer mechanism must occur with concomitant reduction of the metal center [
22]. No evidence for self-association was observed by NMR studies for
A or for free ligands (
Figure S6). Previously published work discussed the reduction potential and accessibility of the cysteine residues of the C-terminal zinc finger, and this data suggests that disulfide bond formation would likely occur between C39 residues (
Figure 5), as C39 has a reduction potential nearly as low as that of C49 (−33.35 vs. −35.91 mV) but has a much greater accessibility (52.88 vs. 15.17) [
22]. It is also possible that reduction of multiple molecules of
A or
B could also result in the formation of multiple disulfide bonds, in which case both C49 and C39 could form disulfide bonds to form the dimer.
4. Materials and Methods
[AuCl(dien)]Cl
2 and [AuCl(
N-Medien)]Cl
2 were synthesized according to previously published procedures [
23]. The 1-methylcytosine was synthesized and purified according to previously published procedures [
24,
25]. The addition of the
N-heterocycles were carried out analogously to previously published procedures [
10]. The apoNCp7-F2 was purchased from GenScript (Piscataway, NJ, USA). NCp7-F2 was prepared according to previously published procedures [
10]. All nuclear magnetic resonance (NMR) studies were carried out in deuterium oxide on a Bruker AVANCE III (400 MHz) (Bruker, Billerica, MA, USA). CD studies were carried out at 50 µM concentrations in water on a Jasco J-1500 spectrometer (Jasco, Easton, MD, USA). Mass spectrometry studies were carried out on a Thermo Electron Corporation Orbitrap Velos mass spectrometer (Thermo Scientific, Waltham, MA, USA). Mass spectrometry samples were prepared at 10 µM concentrations in 10% methanol, and were sprayed at 2.30 kV and 0.9 µL/minute at 230 °C.
Synthesis of [Au(dien)(1-Mecyt)](NO3)3 (A): [AuCl(dien)]Cl2 (101.2 mg, 0.249 mmol) was dissolved in dH2O (10 mL).The 1-methylcytosine (30.2 mg, 0.242 mmol) and silver nitrate (125.4 mg, 0.738 mmol) were added, and the reaction mixture was stirred under light protection for 72 h. The precipitated silver chloride was removed by filtration through celite, and the filtrate was evaporated to dryness under reduced pressure. The orange/yellow residue was dissolved in a small amount of dH2O, and acetone was added to the cloud point. The orange/yellow solid product was collected by vacuum filtration and characterized by 1H-NMR, infrared spectroscopy, and elemental analysis (experimental (theoretical)): C: 14.46 (14.62) H: 2.81 (2.86) N: 17.70 (17.17). 1H NMR (D2O, 400 MHz): 7.83, 7.73 (d, 1H), 6.14 (d, 1H), 3.69 (broad, 6H), 3.45 (s, 3H), 3.18 (broad, 2H). UV–Vis (1 mM, H2O): 237 nm (Abs = 3.98), 357 nm (Abs = 0.408).
Synthesis of [Au(N-Medien)(1-MeCyt)](NO3)3 (B): [AuCl(N-MeDien)]Cl2 (100.12 mg, 0.238 mmol) was dissolved in dH2O (10 mL). Silver nitrate (121.4 mg, 0.715 mmol) and 1-methylcytosine (29.8 mg, 0.238 mmol) were added and the reaction mixture was stirred under light protection for 4 days. The reaction mixture was filtered through celite and the solvent was evaporated under reduced pressure. The resulting crude product was dissolved in a small amount of water and excess acetone was added to precipitate the yellow product, characterized by 1H-NMR, Far-IR, and elemental analysis (experimental (theoretical)): C: 16.77 (17.20) H: 3.33 (3.46) N: 17.69 (18.05). 1H-NMR (D2O, 400 MHz): 7.82, 7.72 (d,1H), 6.13(d, 1H), 4.21, 4.09 (m, 4H), 3.65 (m, 3H), 3.46 (d, 2H), 3.38 (s, 3H), 3.19 (m, 2H). UV–Vis (1 mM, H2O): 237 nm (Abs = 3.94), 352 nm (Abs = 0.921).
Synthesis of [Au(dien)(Cyt)](NO3)3 (C): [AuCl(dien)]Cl2 (100.44 mg, 0.168 mmol) was dissolved in dH2O (10 mL). Cytosine (28.2 mg, 0.164 mmol) and silver nitrate (85.61 mg, 0.506 mmol) were added, and the reaction mixture was stirred under light protection overnight. The precipitated silver chloride was removed by filtration through celite, and the filtrate was evaporated to dryness under reduced pressure. The orange residue was dissolved in a small amount of dH2O, and acetone was added to the cloud point. The orange solid product was collected by vacuum filtration. 1H-NMR (D2O, 400 MHz): 7.64 (d, 1H), 6.06 (d, 1H), 3.52 (t, 4H), 2.99 (broad, 4H).
Fluorescence quenching of tryptophan: Tryptophan quenching methods were adapted from those previously published [
10]. A solution of
N-acetyl-L-tryptophan (5 µM) was titrated with the Au(III) complexes (7.5 mM) in amounts ranging from 1-10 equivalents of quencher. The decrease in fluorescence intensity of
N-acetyl-L-tryptophan was plotted using the Eadie-Hofstee method in order to determine the association constant.
Determination of self-association by NMR spectroscopy: Varying concentrations (0.1–2.5 mM) of
N-heterocycle and gold(III) complex were analyzed by
1H-NMR. The degree of self-association was determined by previously published methods [
26]. The change in chemical shift with increasing concentration was fitted to the equation
. Microsoft Excel Solver was used to solve the three-variable equation.
Cyclic voltammetry: CV studies were carried out following the previously published procedures [
10]. A three-electrode cell was employed, containing a platinum disk working electrode, a platinum mesh counter electrode, and a silver/silver chloride reference electrode (0.1 M KCl). Solutions were degassed with nitrogen for 15 minutes before analysis. Measurements were obtained in 50 mM phosphate buffer (4 mM NaCl, pH = 7.4).
5. Conclusions
In conclusion, the previously unreported 1-methylcytosine complexes based on the [Au(dien)(
N-heterocycle)]
3+ motif,
A, and
B, have been synthesized and investigated for their ability to interact with the C-terminal finger of the HIV nucleocapsid protein NCp7 as a model for a protein with highly nucleophilic cysteines. Gold compounds are considered to primarily interfere with protein, rather than DNA, function. In considering the series [M(dien)L]
n+, the electronic properties can be modulated systematically for optimal protein interaction. A key feature leading to the rational design of complexes of the form [Au(dien)L]
3+ is the enhancement of the redox stability of the complexes relative to the precursor [AuCl(dien)]
2+, due to the incorporation of a more substitution inert nitrogen donor in the form of an
N-heterocycle [
9]. In the case of the more sterically hindered complex
B, the reduction potential is greater than the chloride analog [AuCl(
N-Medien)]
2+, and is in line with the reduction in stabilization of the gold(III) oxidation state seen in the complex [Au(
N-Medien)(DMAP)]
3+. The aurated nucleobase may interact preferentially with an RNA(DNA) recognition motif, such as tryptophan. Formation of oxidized dimerized peptide was observed in the mass spectrometry studies conducted with NCp7-F2, and indicated a difference in the reactivity profile of
A and
B compared to the previously studied DMAP and 9-EtGua analogs. These results further confirm that tuning the electronic and steric properties of the ligands on the Au(III) coordination sphere can modulate the strength of interaction between complexes of the [Au(dien)(
N-heterocycle)]
3+ motif and a zinc finger a systematic manner.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}