Changes in Cell Biology under the Influence of Low-Level Laser Therapy

 , , and
, , and 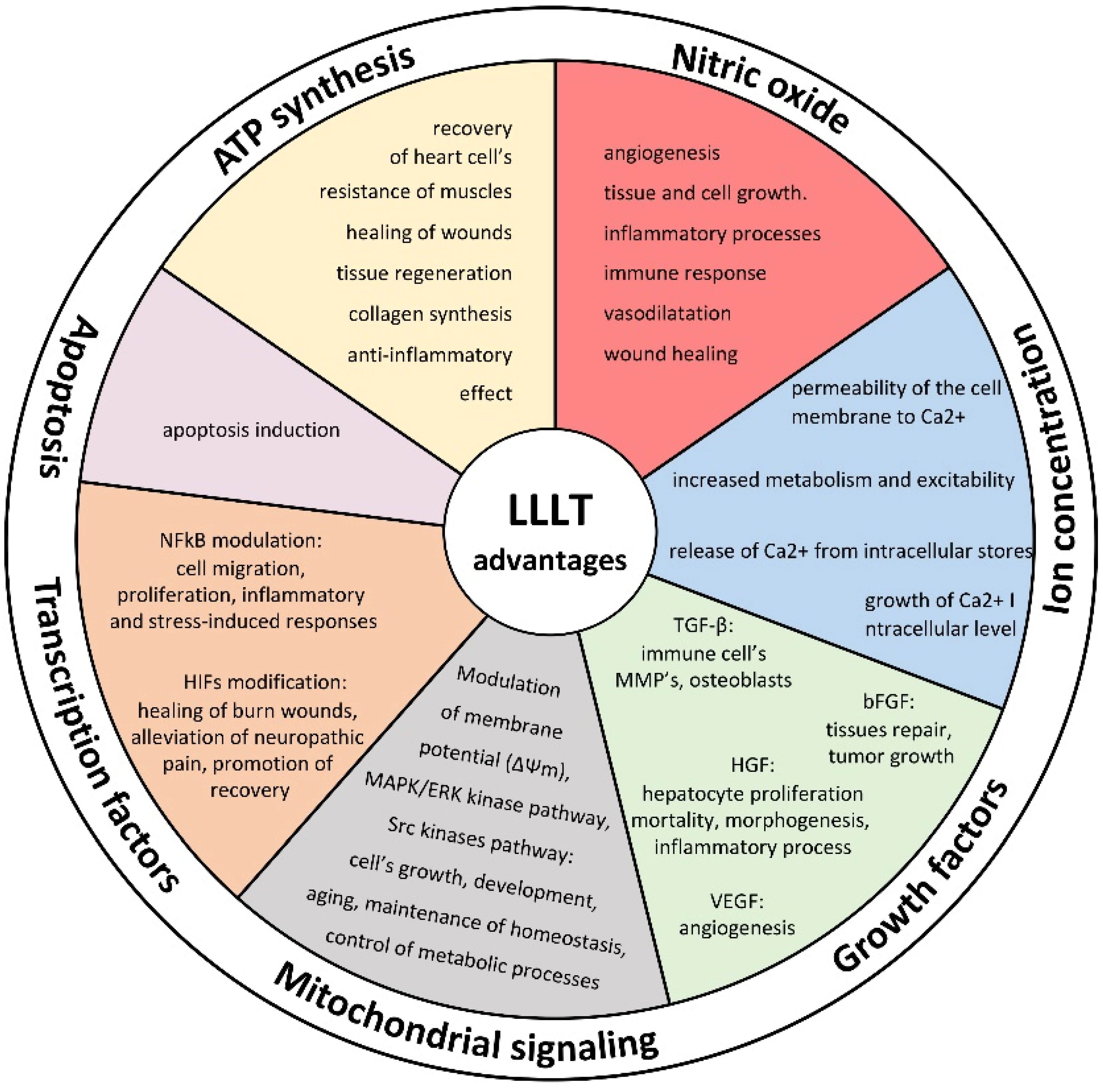{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
Study Aim
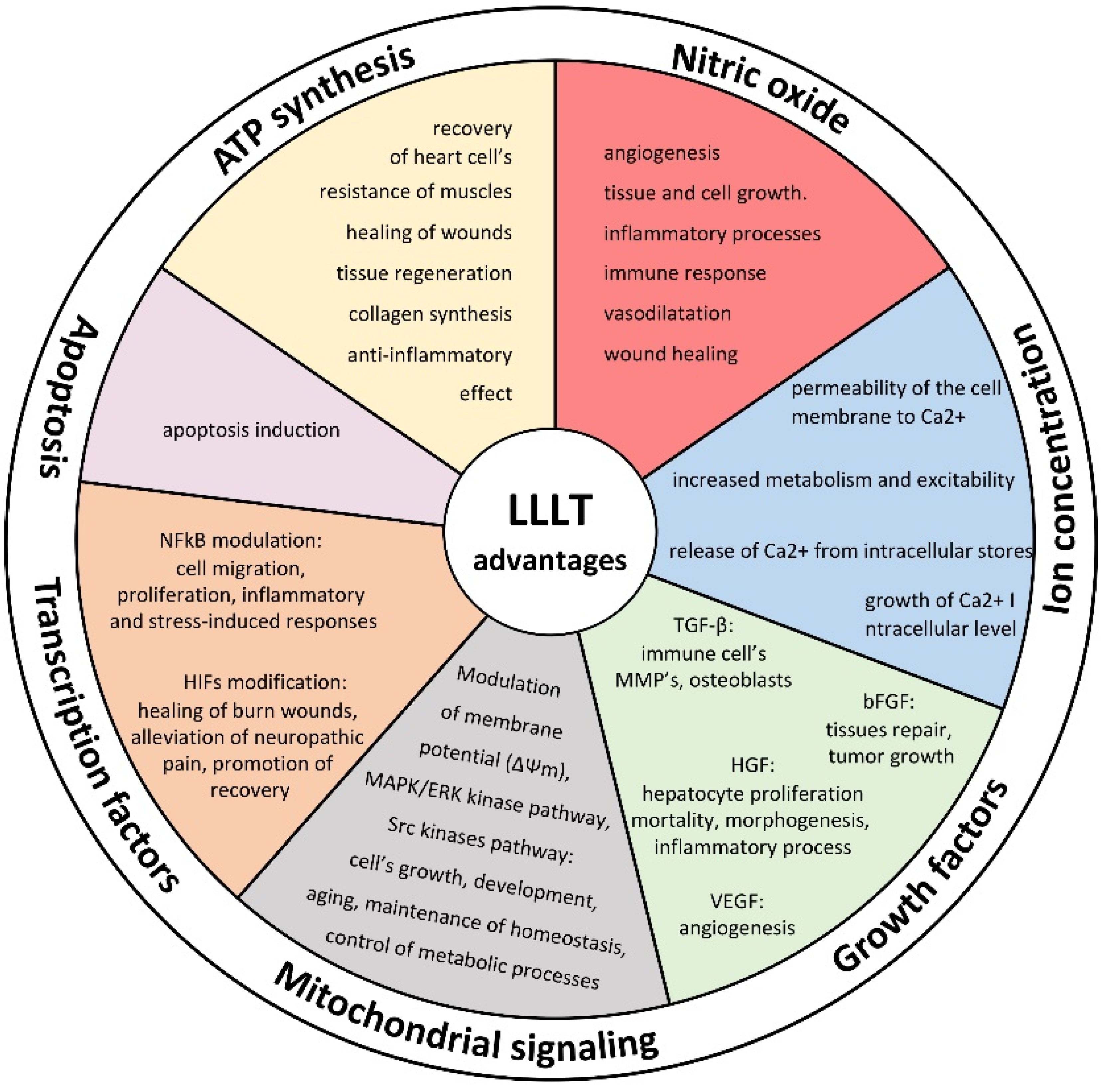
2. ATP Synthesis
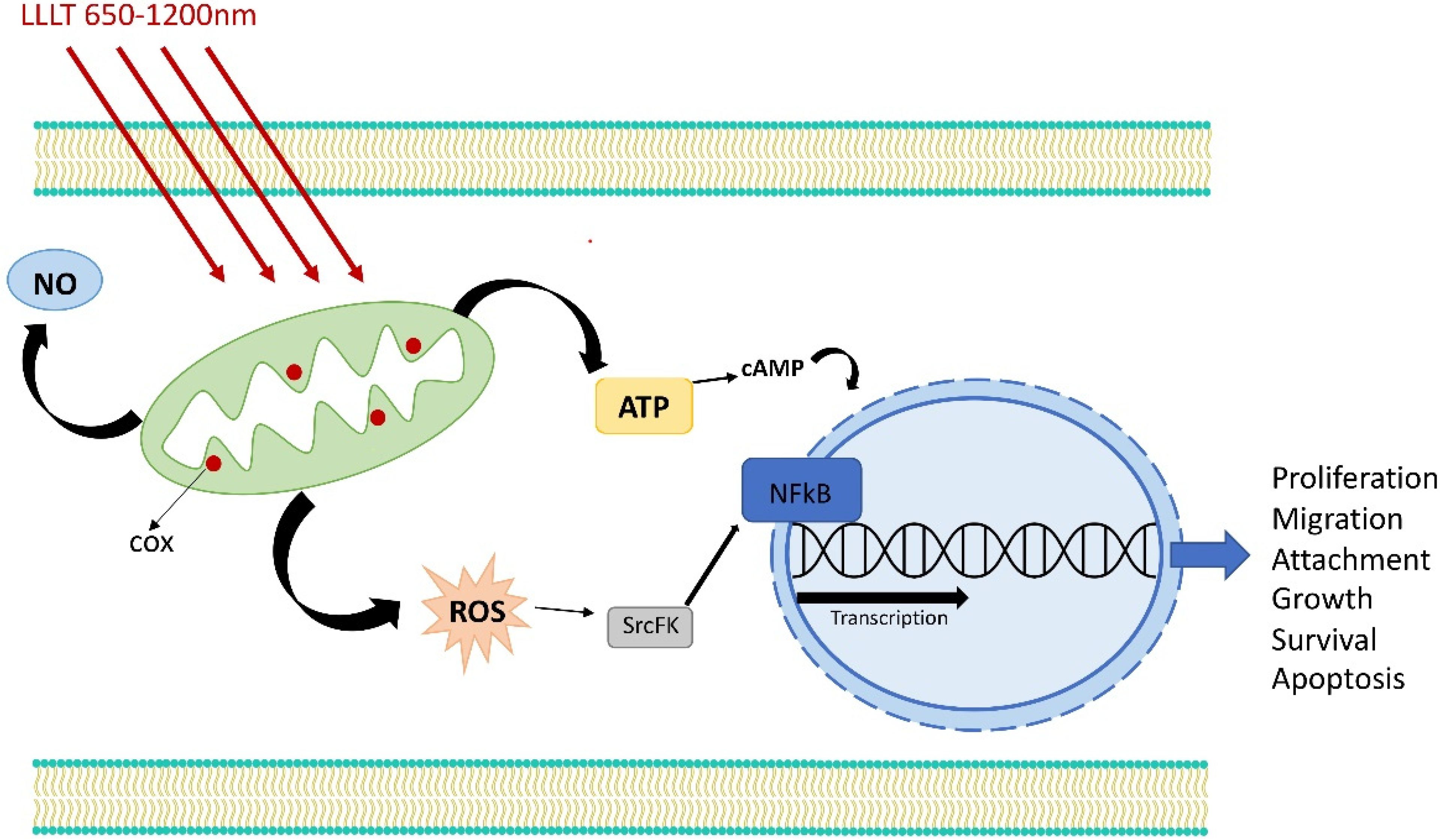
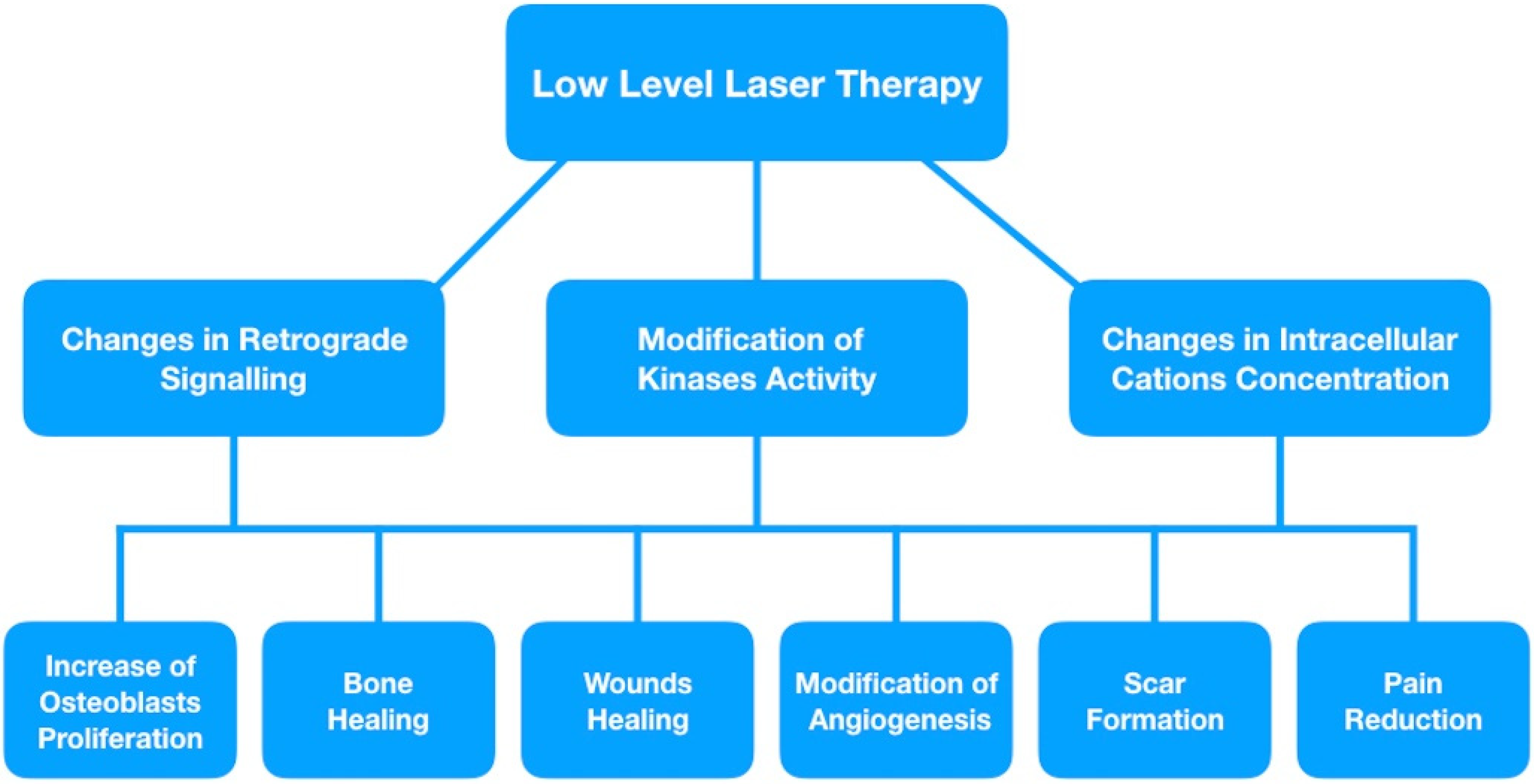
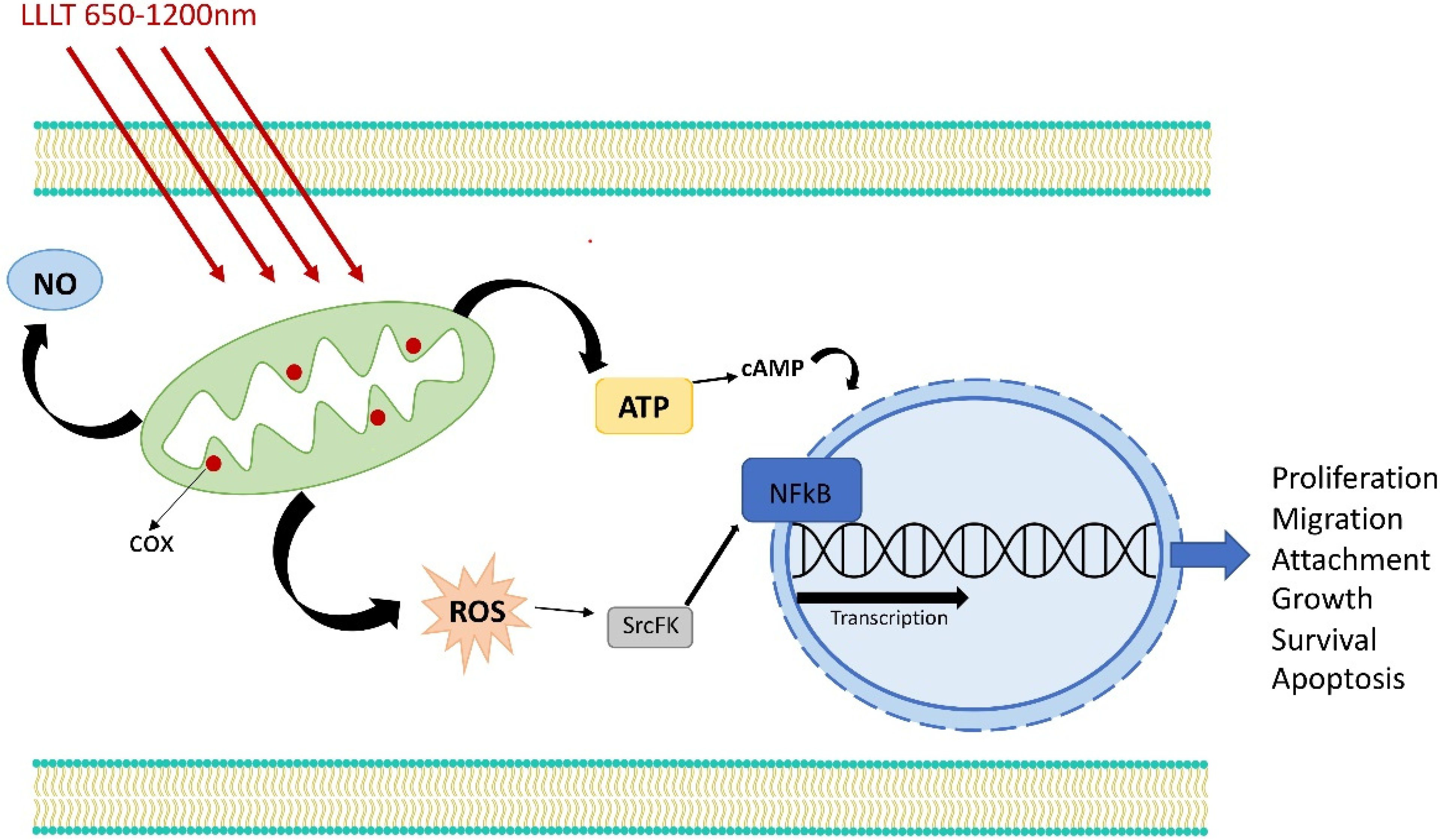
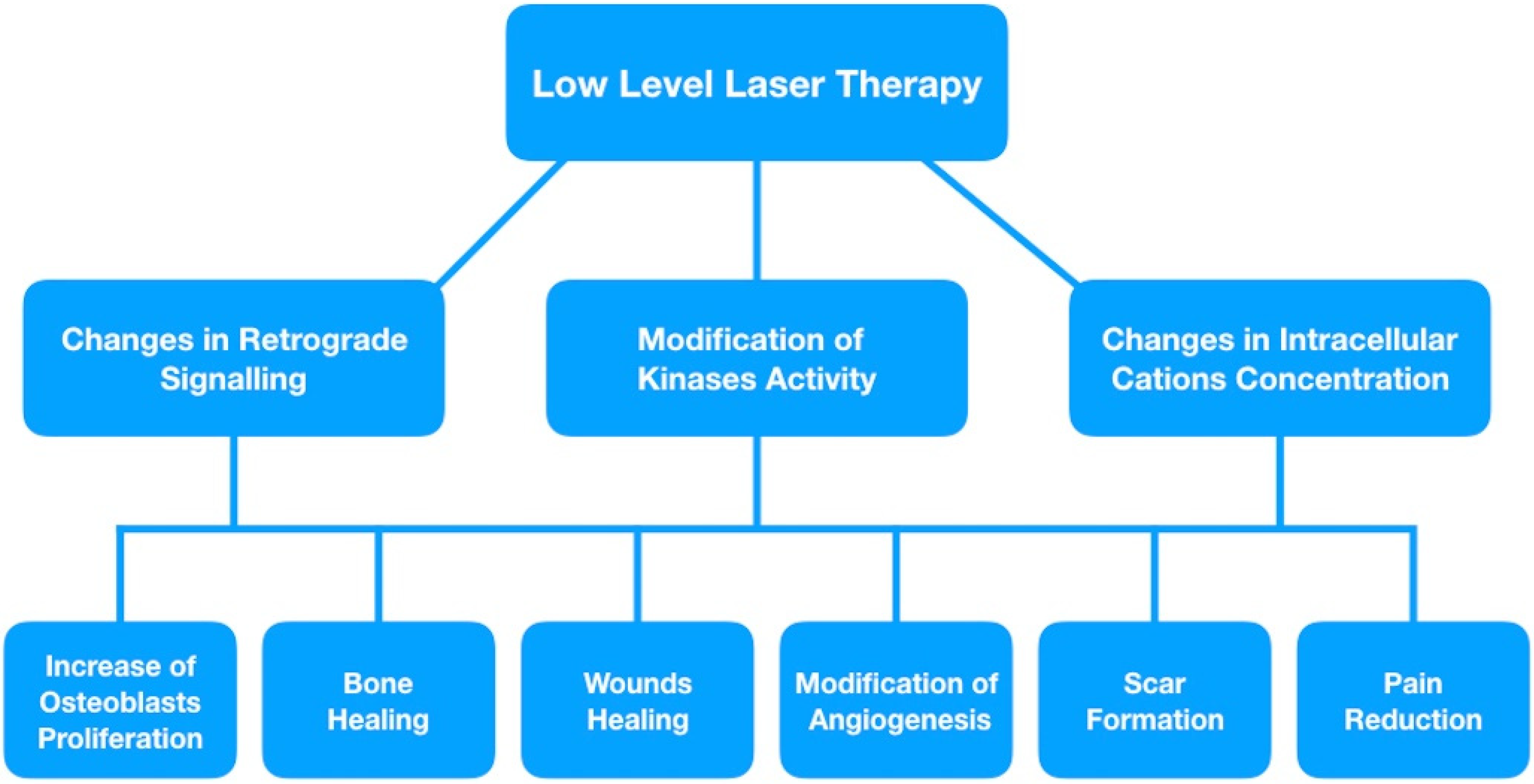
3. Retrograde Mitochondrial Signaling
4. Nitric Oxide
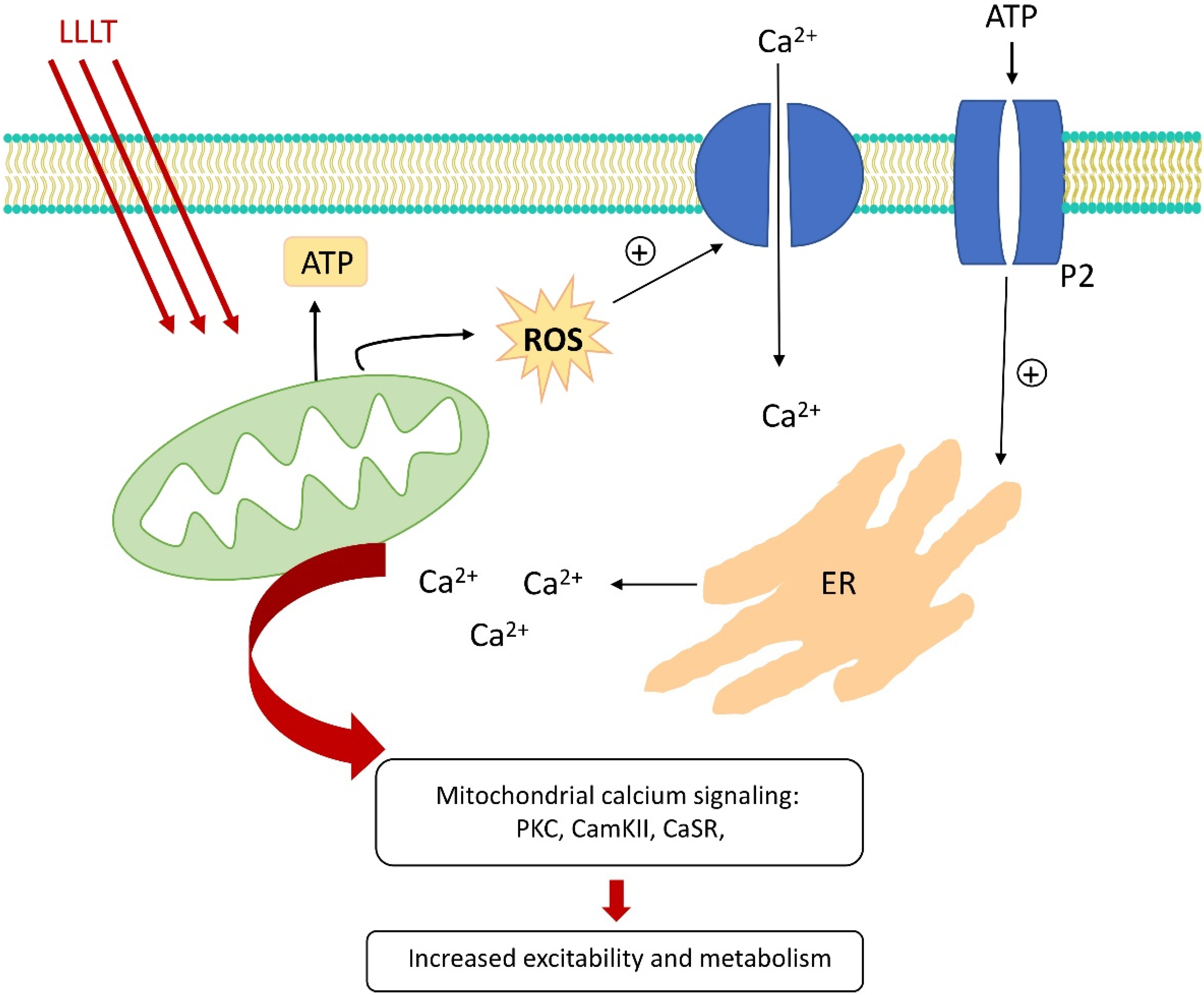
5. Modulation of Ion Concentrations
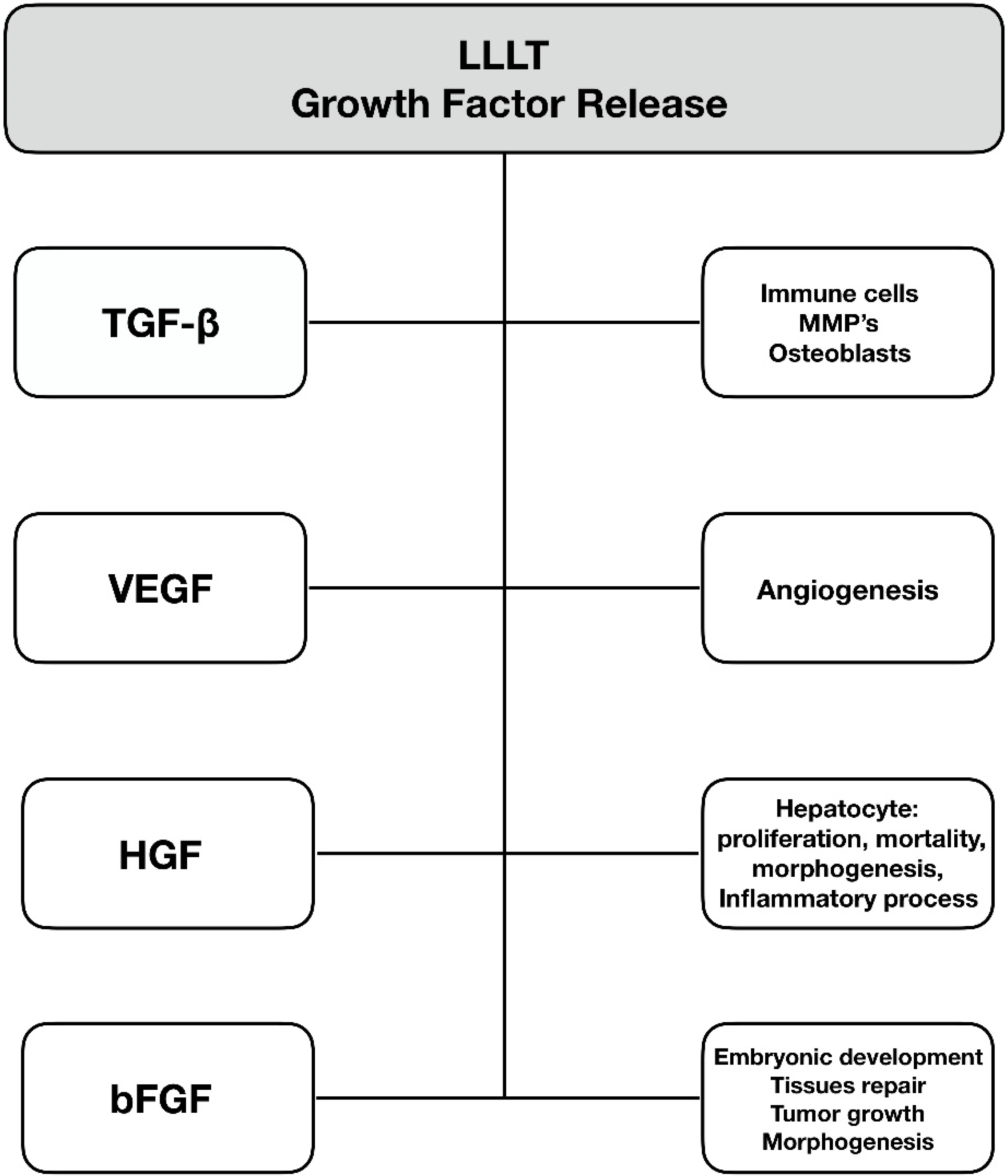
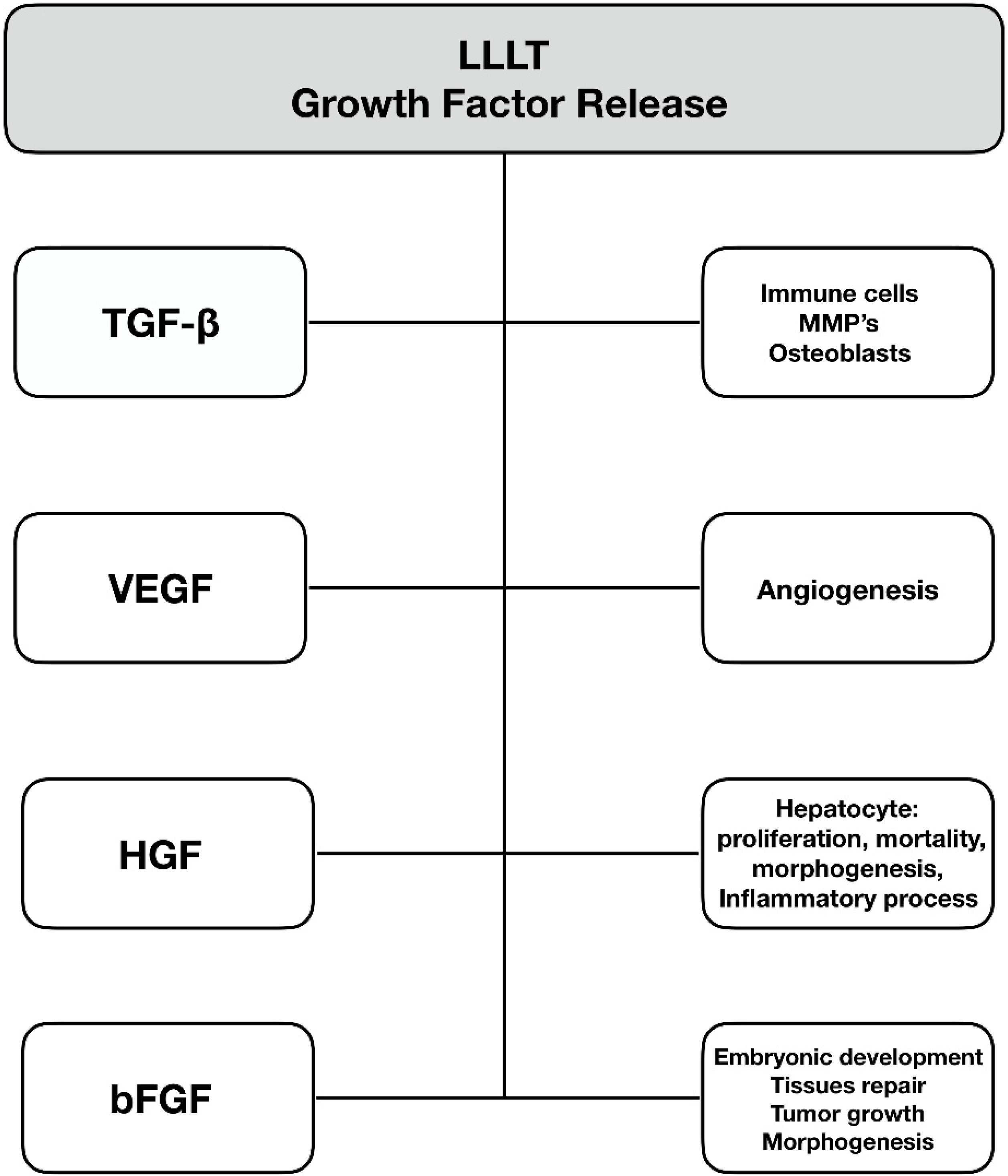
6. Growth Factor Release
6.1. Transforming Growth Factor-Beta
6.2. Vascular Endothelial Growth Factor (VEGF)
6.3. Hepatocyte Growth Factor (HGF)
6.4. Basic Fibroblast Growth Factor (bFGF)
7. Activation of Transcription Factors
8. Influence on Apoptosis
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mester, E.; Szende, B.; Gärtner, P. The effect of laser beams on the growth of hair in mice. Radiobiol. Radiother. 1968, 9, 621–626. [Google Scholar]
- Piotr, R.; Doroszko, A.; Derkacz, A. The Use of Low-Level Energy Laser Radiation in Basic and Clinical Research. Adv. Clin. Exp. Med. 2014, 23, 835–842. [Google Scholar]
- Zecha, J.A.; Raber-Durlacher, J.E.; Nair, R.G.; Epstein, J.B.; Sonis, S.T.; Elad, S.; Hamblin, M.R.; Barasch, A.; Migliorati, C.A.; Milstein, D.M.; et al. Low level laser therapy/photobiomodulation in the management of side effects of chemoradiation therapy in head and neck cancer: Part 1: Mechanisms of action, dosimetric, and safety considerations. Supportive Care Cancer 2016, 24, 2781–2792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, S.Y.; Tam, V.C.W.; Ramkumar, S.; Khaw, M.L.; Law, H.K.W.; Lee, S.W.Y. Review on the Cellular Mechanisms of Low-Level Laser Therapy Use in Oncology. Front. Oncol. 2020, 10, 1255. [Google Scholar] [CrossRef]
- Hashmi, J.T.; Huang, Y.Y.; Sharma, S.K.; Kurup, D.B.; de Taboada, L.; Carroll, J.D.; Hamblin, M.R. Effect of pulsing in low-level light therapy. Lasers Surg. Med. 2010, 42, 450–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balderas, P.M.D.O. Mitochondria-plasma membrane interactions and communication. J. Biol. Chem. 2021, 297, 101164. [Google Scholar] [CrossRef] [PubMed]
- Santucci, R.; Sinibaldi, F.; Cozza, P.; Polticelli, F.; Fiorucci, L. Cytochrome c: An extreme multifunctional protein with a key role in cell fate. Int. J. Biol. Macromol. 2019, 136, 1237–1246. [Google Scholar] [CrossRef]
- Salehpour, F.; Mahmoudi, J.; Kamari, F.; Sadigh-Eteghad, S.; Rasta, S.H.; Hamblin, M.R. Brain Photobiomodulation Therapy: A Narrative Review. Mol. Neurobiol. 2018, 55, 6601–6636. [Google Scholar] [CrossRef]
- Salehpour, F.; Mahmoudi, J.; Kamari, F.; Sadigh-Eteghad, S.; Rasta, S.H.; Hamblin, M.R. Photobiomodulation Therapy Attenuates Hypoxic-Ischemic Injury in a Neonatal Rat Model. J. Mol. Neurosci. 2018, 65, 514–526. [Google Scholar]
- Huang, Y.Y.; Chen, A.C.H.; Carroll, J.D.; Hamblin, M.R. Biphasic dose response in low level light therapy. Dose-Response 2009, 7. [Google Scholar] [CrossRef]
- Karu, T.I. Mitochondrial signaling in mammalian cells activated by red and near-IR radiation. Photochem. Photobiol. 2008, 84, 1091–1099. [Google Scholar] [CrossRef] [PubMed]
- Kadenbach, B.; Huettemann, M. The subunit composition and function of mammalian cytochrome c oxidase. Mitochondrion 2015, 24, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Yu, W.; Yang, X.; Hicks, G.L.; Lanzafame, R.J.; Wang, T. Photo-irradiation improved functional preservation of the isolated rat heart. Lasers Surg. Med. 1997, 20, 332–339. [Google Scholar] [CrossRef]
- Kasprzyk-Kucewicz, T.; Szurko, A.; Stanek, A.; Sieroń, K.; Morawiec, T.; Cholewka, A. Usefulness in Developing an Optimal Training Program and Distinguishing between Performance Levels of the Athlete’s Body by Using of Thermal Imaging. Int. J. Environ. Res. Public. Health 2020, 17, 5698. [Google Scholar] [CrossRef] [PubMed]
- Ferraresi, C.; de Sousa, M.V.P.; Huang, Y.Y.; Bagnato, V.S.; Parizotto, N.A.; Hamblin, M.R. Time response of increases in ATP and muscle resistance to fatigue after low-level laser (light) therapy (LLLT) in mice. Lasers Med. Sci. 2015, 30, 1259–1267. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.; Gupta, A.; Keshri, G.K.; Verma, S.; Sharma, S.K.; Singh, S.B. Photobiomodulatory effects of superpulsed 904 nm laser therapy on bioenergetics status in burn wound healing. J. Photochem. Photobiol. B Biol. 2016, 162, 77–85. [Google Scholar] [CrossRef]
- Xu, Z.; Guo, X.; Yang, Y.; Tucker, D.; Lu, Y.; Xin, N.; Zhang, G.; Yang, L.; Li, J.; Du, X.; et al. Low-level laser irradiation improves depression-like behaviors in mice. Mol. Neurobiol. 2017, 54, 4551–4559. [Google Scholar] [CrossRef]
- Chen, M.-H.; Huang, Y.-C.; Sun, J.-S.; Chao, Y.-H.; Chen, M.-H. Second messengers mediating the proliferation and collagen synthesis of tenocytes induced by low-level laser irradiation. Lasers Med. Sci. 2015, 30, 263–272. [Google Scholar] [CrossRef]
- Hamblin, M.R. Mechanisms and applications of the anti-inflammatory effects of photobiomodulation. AIMS Biophys. 2017, 4, 337. [Google Scholar] [CrossRef]
- Wang, L.; Hu, L.; Grygorczyk, R.; Shen, X.; Schwarz, W. Modulation of extracellular ATP content of mast cells and DRG neurons by irradiation: Studies on underlying mechanism of low-level-laser therapy. Mediat. Inflamm. 2015, 2015, 630361. [Google Scholar] [CrossRef]
- Amaroli, A.; Benedicenti, A.; Ravera, S.; Parker, S.; Selting, W.; Panfoli, I.; Benedicenti, S. Short-pulse neodymium: Yttrium–aluminium garnet (Nd: YAG 1064 nm) laser irradiation photobiomodulates mitochondria activity and cellular multiplication of Paramecium primaurelia (Protozoa). Eur. J. Protistol. 2017, 61, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Rola, P.; Doroszko, A.; Szahidewicz-Krupska, E.; Rola, P.; Dobrowolski, P.; Skomro, R.; Szymczyszyn, A.; Mazur, G.; Derkacz, A. Low-level laser irradiation exerts antiaggregative effect on human platelets independently on the nitric oxide metabolism and release of platelet activation markers. Oxidative Med. Cell. Longev. 2017, 2017, 6201797. [Google Scholar] [CrossRef] [PubMed]
- Mielecki, J.; Gawroński, P.; Karpiński, S. Retrograde Signaling: Understanding the Communication between Organelles. Int. J. Mol. Sci. 2020, 21, 6173. [Google Scholar] [CrossRef] [PubMed]
- Ježek, P.; Holendová, B.; Plecitá-Hlavatá, L. Redox Signaling from Mitochondria: Signal Propagation and Its Targets. Biomolecules 2020, 10, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shefer, G.; Oron, U.; Irintchev, A.; Wernig, A.; Halevy, O. Skeletal muscle cell activation by low-energy laser irradiation: A role for the MAPK/ERK pathway. J. Cell. Physiol. 2001, 187, 73–80. [Google Scholar] [CrossRef]
- Hu, W.-P.; Wang, J.-J.; Yu, C.-L.; Lan, C.-C.E.; Chen, G.-S.; Yu, H.-S. Helium–neon laser irradiation stimulates cell proliferation through photostimulatory effects in mitochondria. J. Investig. Dermatol. 2007, 187, 2048–2057. [Google Scholar]
- Stocker, R.; Keaney, J.F., Jr. Role of oxidative modifications in atherosclerosis. Physiol. Rev. 2004, 84, 1381–1478. [Google Scholar] [CrossRef]
- Zhang, J.; Xing, D.; Gao, X. Low-power laser irradiation activates Src tyrosine kinase through reactive oxygen species-mediated signaling pathway. J. Cell. Physiol. 2008, 217, 518–528. [Google Scholar] [CrossRef]
- Chen, A.C.-H.; Arany, P.R.; Huang, Y.-Y.; Tomkinson, E.M.; Sharma, S.K.; Kharkwal, G.B.; Saleem, T.; Mooney, D.; Yull, F.E.; Blackwell, T.S.; et al. Low-level laser therapy activates NF-kB via generation of reactive oxygen species in mouse embryonic fibroblasts. PLoS ONE 2011, 6, e22453. [Google Scholar]
- De Luca, F. Role of Nuclear Factor Kappa B (NF-κB) in Growth Plate Chondrogenesis. Pediatric Endocrinol. Rev. 2016, 13, 720–730. [Google Scholar]
- Jakubiak, G.K.; Cieślar, G.; Stanek, A. Nitrotyrosine, Nitrated Lipoproteins, and Cardiovascular Dysfunction in Patients with Type 2 Diabetes: What Do We Know and What Remains to Be Explained? Antioxidants 2022, 11, 856. [Google Scholar] [CrossRef]
- Duda, D.G.; Fukumura, D.; Jain, R.K. Role of eNOS in neovascularization: NO for endothelial progenitor cells. Trends Mol. Med. 2004, 10, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation. J. Physiol. Pharm. 2003, 54, 469–487. [Google Scholar]
- Stanek, A.; Cholewka, A.; Wielkoszyński, T.; Romuk, E.; Sieroń, K.; Sieroń, A. Increased Levels of Oxidative Stress Markers, Soluble CD40 Ligand, and Carotid Intima-Media Thickness Reflect Acceleration of Atherosclerosis in Male Patients with Ankylosing Spondylitis in Active Phase and without the Classical Cardiovascular Risk Factors. Oxid. Med. Cell. Longev. 2017, 2017, 9712536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopp-Scheinpflug, C.; Forsythe, I.D. Nitric Oxide Signaling in the Auditory Pathway. Front Neural Circuits. 2021, 15, 114. [Google Scholar] [CrossRef]
- Karu, T.I.; Pyatibrat, L.V.; Afanasyeva, N.I. Cellular effects of low power laser therapy can be mediated by nitric oxide. Lasers Surg. Med. 2005, 36, 307–314. [Google Scholar] [CrossRef]
- Mitchell, U.H.; Mack, G.L. Low-level laser treatment with near-infrared light increases venous nitric oxide levels acutely: A single-blind, randomized clinical trial of efficacy. Am. J. Phys. Med. Rehabil. 2013, 92, 151–156. [Google Scholar] [CrossRef]
- Tuby, H.; Maltz, L.; Oron, U. Modulations of VEGF and iNOS in the rat heart by low level laser therapy are associated with cardioprotection and enhanced angiogenesis. Lasers Surg. Med. 2006, 38, 682–688. [Google Scholar] [CrossRef]
- Samoilova, K.A.; Zhevago, N.A.; Petrishchev, N.N.; Zimin, A.A. Role of nitric oxide in the visible light-induced rapid increase of human skin microcirculation at the local and systemic levels: II. healthy volunteers. Photomed. Laser Surg. 2008, 26, 443–449. [Google Scholar] [CrossRef] [Green Version]
- Poderoso, J.J.; Helfenberger, K.; Poderoso, C. The effect of nitric oxide on mitochondrial respiration. Nitric Oxide 2019, 88, 61–72. [Google Scholar] [CrossRef]
- Karu, T.I.; Pyatibrat, L.V.; Kalendo, G.S. Photobiological modulation of cell attachment via cytochrome c oxidase. Photochem. Photobiol. Sci. 2004, 3, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Hung, H.S.; Hsu, S.H. Low-energy laser irradiation increases endothelial cell proliferation, migration, and eNOS gene expression possibly via PI3K signal pathway. Lasers Surg. Med. 2008, 40, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Kazemikhoo, N.; Sarafnejad, A.F.; Ansari, F.; Mehdipour, P. Modifying effect of intravenous laser therapy on the protein expression of arginase and epidermal growth factor receptor in type 2 diabetic patients. Lasers Med. Sci. 2016, 31, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Lindgård, A.; Hultén, L.M.; Svensson, L.; Soussi, B. Irradiation at 634 nm releases nitric oxide from human monocytes. Lasers Med. Sci. 2007, 22, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Mittermayr, R.; Osipov, A.; Piskernik, C.; Haindl, S.; Dungel, P.; Weber, C.; Vladimirov, Y.A.; Redl, H.; Kozlov, A.V. Blue laser light increases perfusion of a skin flap via release of nitric oxide from hemoglobin. Mol. Med. 2007, 13, 22. [Google Scholar] [CrossRef]
- Lubart, R.; Friedmann, H.; Sinyakov, M.; Cohen, N.; Breitbart, H. Changes in calcium transport in mammalian sperm mitochondria and plasma membranes caused by 780 nm irradiation. Lasers Surg. Med. 1997, 21, 493–499. [Google Scholar] [CrossRef]
- Abdel-Magied, N.; Elkady, A.A.; Fattah, S.M.A. Effect of Low-Level Laser on Some Metals Related to Redox State and Histological Alterations in the Liver and Kidney of Irradiated Rats. Biol Trace Elem. Res. 2020, 194, 410–422. [Google Scholar] [CrossRef]
- Harraz, O.F.; Jensen, L.J. Vascular calcium signalling and ageing. J. Physiol. 2021, 599, 5361–5377. [Google Scholar] [CrossRef]
- Migliario, M.; Sabbatini, M.; Mortellaro, C.; Renò, F. Near infrared low-level laser therapy and cell proliferation: The emerging role of redox sensitive signal transduction pathways. J. Biophotonics 2018, 11, e201800025. [Google Scholar] [CrossRef]
- Kalthof, B.; Bechem, M.; Flocke, K.; Pott, L.; Schramm, M. Kinetics of ATP-induced Ca2+ transients in cultured pig aortic smooth muscle cells depend on ATP concentration and stored Ca2+. J. Physiol. 1993, 466, 245–262. [Google Scholar]
- Kitajima, S.; Ozaki, H.; Karaki, H. Role of different subtypes of P2 purinoceptor on cytosolic Ca2+ levels in rat aortic smooth muscle. Eur. J. Pharmacol. 1994, 266, 263–267. [Google Scholar] [CrossRef]
- Yang, W.-Z.; Chen, J.-Y.; Yu, J.-T.; Zhou, L.-W. Effects of low power laser irradiation on intracellular calcium and histamine release in RBL-2H3 mast cells. Photochem. Photobiol. 2007, 83, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Leyane, T.S.; Jere, S.W.; Houreld, N.N. Cellular Signalling and Photobiomodulation in Chronic Wound Repair. Int. J. Mol. Sci. 2021, 22, 11223. [Google Scholar] [CrossRef] [PubMed]
- Kujawa, J.; Zavodnik, L.; Zavodnik, I.; Buko, V.; Lapshyna, A.; Bryszewska, M. Effect of low-intensity (3.75–25 J/cm2) near-infrared (810 nm) laser radiation on red blood cell ATPase activities and membrane structure. J. Clin. Laser Med. Surg. 2004, 22, 111–117. [Google Scholar] [CrossRef] [PubMed]
- De Lima, H.S.; Rigos, C.F.; Tedesco, A.C.; Ciancaglini, P. Biostimulation of Na, K-ATPase by low-energy laser irradiation (685 nm, 35 mW): Comparative effects in membrane, solubilized and DPPC: DPPE-liposome reconstituted enzyme. J. Photochem. Photobiol. B Biol. 2007, 89, 22–28. [Google Scholar] [CrossRef]
- Lopatina, E.V.; Yachnev, I.L.; Penniyaynen, V.A.; Plakhova, V.B.; Podzorova, S.A.; Shelykh, T.N.; Rogachevsky, I.V.; Butkevich, I.P.; Mikhailenko, V.A.; Kipenko, A.V.; et al. Modulation of signal-transducing function of neuronal membrane Na+, K+-ATPase by endogenous ouabain and low-power infrared radiation leads to pain relief. Med. Chem. 2012, 8, 33–39. [Google Scholar] [CrossRef]
- Hao, Y.; Baker, D.; Dijke, P.T. TGF-β-Mediated Epithelial-Mesenchymal Transition and Cancer Metastasis. Int. J. Mol. Sci. 2019, 20, 2767. [Google Scholar] [CrossRef] [Green Version]
- Nouruzian, M.; Alidoust, M.; Bayat, M.; Bayat, M.; Akbari, M. Effect of low-level laser therapy on healing of tenotomized Achilles tendon in streptozotocin-induced diabetic rats. Lasers Med. Sci. 2013, 28, 399–405. [Google Scholar] [CrossRef]
- Pyo, S.-J.; Song, W.-W.; Kim, I.-R.; Park, B.-S.; Kim, C.-H.; Shin, S.-H.; Chung, I.-K.; Kim, Y.-D. Low-level laser therapy induces the expressions of BMP-2, osteocalcin, and TGF-β1 in hypoxic-cultured human osteoblasts. Lasers Med. Sci. 2013, 28, 543–550. [Google Scholar] [CrossRef]
- Fallahnezhad, S.; Piryaei, A.; Darbandi, H.; Amini, A.; Ghoreishi, S.K.; Jalalifirouzkouhi, R.; Bayat, M. Effect of low-level laser therapy and oxytocin on osteoporotic bone marrow-derived mesenchymal stem cells. J. Cell. Biochem. 2018, 119, 983–997. [Google Scholar] [CrossRef]
- Dang, Y.; Liu, B.; Liu, L.; Ye, X.; Bi, X.; Zhang, Y.; Gu, J. The 800-nm diode laser irradiation induces skin collagen synthesis by stimulating TGF-β/Smad signaling pathway. Lasers Med. Sci. 2011, 26, 837. [Google Scholar] [CrossRef] [PubMed]
- Schardong, J.; Falster, M.; Sisto, I.R.; Barbosa, A.P.O.; Normann, T.C.; De Souza, K.S.; Jaroceski, G.; Bozzetto, C.B.; Baroni, B.M.; Plentz, R.D.M. Photobiomodulation therapy increases functional capacity of patients with chronic kidney failure: Randomized controlled trial. Lasers Med. Sci. 2021, 36, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Guerra, F.D.R.; Vieira, C.P.; Oliveira, L.P.; Marques, P.P.; Almeida, M.D.S.; Pimentel, E.R. Low-level laser therapy modulates pro-inflammatory cytokines after partial tenotomy. Lasers Med. Sci. 2016, 31, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Höfling, D.B.; Chavantes, M.C.; Acencio, M.M.; Cerri, G.G.; Marui, S.; Yoshimura, E.M.; Chammas, M.C. Effects of Low-Level Laser Therapy on the Serum TGF-β1 Concentrations in Individuals with Autoimmune Thyroiditis. Photomed. Laser Surg. 2014, 32, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Barolet, D.; Boucher, A. Prophylactic low-level light therapy for the treatment of hypertrophic scars and keloids: A case series. Lasers Surg. Med. 2010, 42, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Derkacz, A.; Protasiewicz, M.; Rola, P.; Podgorska, K.; Szymczyszyn, A.; Gutherc, R.; Poręba, R.; Doroszko, A. Effects of intravascular low-level laser therapy during coronary intervention on selected growth factors levels. Photomed. Laser Surg. 2014, 32, 582–587. [Google Scholar] [CrossRef]
- Melincovici, C.S.; Boşca, A.B.; Şuşman, S.; Mărginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)—Key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar]
- Karaman, S.; Leppänen, V.-M.; Alitalo, K. Vascular endothelial growth factor signaling in development and disease. Development 2018, 145, dev151019. [Google Scholar] [CrossRef] [Green Version]
- Stanek, A.; Fazeli, B.; Bartuś, S.; Sutkowska, E. The role of endothelium in physiological and pathological states-new data. BioMed Res. Int. 2018, 2018, 1098039. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, T.S.; Serra, A.J.; Manchini, M.T.; Bassaneze, V.; Krieger, J.E.; de Tarso Camillo de Carvalho, P.; Espindola Antunes, D.; Sales Bocalini, D.; Ferreira Tucci, P.J.; Silva, J.A., Jr. Effects of low level laser therapy on attachment, proliferation, and gene expression of VEGF and VEGF receptor 2 of adipocyte-derived mesenchymal stem cells cultivated under nutritional deficiency. Lasers Med. Sci. 2015, 30, 217–223. [Google Scholar] [CrossRef]
- Silveira, P.C.L.; Scheffer, D.D.L.; Glaser, V.; Remor, A.P.; Pinho, R.A.; Junior, A.S.A.; Latini, A. Low-level laser therapy attenuates the acute inflammatory response induced by muscle traumatic injury. Free. Radic. Res. 2016, 50, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Szymczyszyn, A.; Doroszko, A.; Szahidewicz-Krupska, E.; Rola, P.; Gutherc, R.; Jasiczek, J.; Mazur, G.; Derkacz, A. Effect of the transdermal low-level laser therapy on endothelial function. Lasers Med. Sci. 2016, 31, 1301–1307. [Google Scholar] [CrossRef]
- Ribatti, D.; Solimando, A.; Pezzella, F. The Anti-VEGF(R) Drug Discovery Legacy: Improving Attrition Rates by Breaking the Vicious Cycle of Angiogenesis in Cancer. Cancers 2021, 13, 3433. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Umitsu, M.; de Silva, D.M.; Roy, A.; Bottaro, D.P. Hepatocyte growth factor/MET in cancer progression and biomarker discovery. Cancer Sci. 2017, 108, 296–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, Y.T.; Aziz, F.; Guerrero-Castilla, A.; Argüelles, S. Signaling Pathways in Inflammation and Anti-inflammatory Therapies. Curr. Pharm. Des. 2018, 24, 1449–1484. [Google Scholar] [CrossRef]
- Shorning, B.Y.; Dass, M.S.; Smalley, M.J.; Pearson, H.B. The PI3K-AKT-mTOR Pathway and Prostate Cancer: At the Crossroads of AR, MAPK, and WNT Signaling. Int. J. Mol. Sci. 2020, 21, 4507. [Google Scholar] [CrossRef]
- Imamura, R.; Matsumoto, K. Hepatocyte growth factor in physiology and infectious diseases. Cytokine 2017, 98, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-S.; Wang, H.; Zhang, Q.-L.; Yang, Z.-J.; Kong, F.; Wu, C.-T. Hepatocyte Growth Factor Gene Therapy for Ischemic Diseases. Hum. Gene Ther. 2018, 29, 413–423. [Google Scholar] [CrossRef]
- Park, I.-S.; Chung, P.-S.; Ahn, J.C. Enhancement of ischemic wound healing by spheroid grafting of human adipose-derived stem cells treated with low-level light irradiation. PLoS ONE 2015, 10, e0122776. [Google Scholar] [CrossRef]
- Araújo, T.G.; de Oliveira, A.G.; Tobar, N.; Saad, M.J.A.; Moreira, L.R.; Reis, E.R.; Nicola, E.M.D.; de Jorge, G.L.; dos Tártaro, R.R.; Boin, I.F.S.F.; et al. Liver regeneration following partial hepatectomy is improved by enhancing the HGF/Met axis and Akt and Erk pathways after low-power laser irradiation in rats. Lasers Med. Sci. 2013, 28, 1511–1517. [Google Scholar] [CrossRef]
- Miyanaga, T.; Ueda, Y.; Miyanaga, A.; Yagishita, M.; Hama, N. Angiogenesis after administration of basic fibroblast growth factor induces proliferation and differentiation of mesenchymal stem cells in elastic perichondrium in an in vivo model: Mini review of three sequential republication-abridged reports. Cell. Mol. Biol. Lett. 2018, 23, 49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Kang, X.; Jin, L.; Bai, J.; Liu, W.; Wang, Z. Stimulation of wound healing using bioinspired hydrogels with basic fibroblast growth factor (bFGF). Int. J. Nanomed. 2018, 13, 3897–3906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viaña-Mendieta, P.; Sánchez, M.L.; Benavides, J. Rational selection of bioactive principles for wound healing applications: Growth factors and antioxidants. Int. Wound J. 2022, 19, 100–113. [Google Scholar] [CrossRef]
- Chen, M.; Bao, L.; Zhao, M.; Cao, J.; Zheng, H. Progress in Research on the Role of FGF in the Formation and Treatment of Corneal Neovascularization. Front. Pharmacol. 2020, 11, 111. [Google Scholar] [CrossRef] [PubMed]
- Akishima-Fukasawa, Y.; Honma, N.; Ogata, H.; Akasaka, Y.; Mikami, T. Angiogenesis in mammary Paget disease: Histopathological analyses of blood vessel density and angiogenic factors. Diagn. Pathol. 2020, 15, 75. [Google Scholar] [CrossRef]
- Yeung, C.H.C.; Schooling, C.M. Systemic inflammatory regulators and risk of Alzheimer’s disease: A bidirectional Mendelian-randomization study. Int. J. Epidemiol. 2021, 50, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, J.; Zhang, K.; Liu, J.; Li, Y.; Su, W.; Song, N. Advances of Fibroblast Growth Factor/Receptor Signaling Pathway in Hepatocellular Carcinoma and its Pharmacotherapeutic Targets. Front. Pharmacol. 2021, 12, 650388. [Google Scholar] [CrossRef]
- Li, R.; Li, D.-H.; Zhang, H.-Y.; Wang, J.; Li, X.-K.; Xiao, J. Growth factors-based therapeutic strategies and their underlying signaling mechanisms for peripheral nerve regeneration. Acta Pharmacol. Sin. 2020, 41, 1289–1300. [Google Scholar] [CrossRef]
- Panés, J.; Rimola, J. Perianal fistulizing Crohn’s disease: Pathogenesis, diagnosis and therapy. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 652–664. [Google Scholar] [CrossRef]
- Taradaj, J.; Shay, B.; Dymarek, R.; Sopel, M.; Walewicz, K.; Beeckman, D.; Schoonhoven, L.; Gefen, A.; Rosińczuk, J. Effect of laser therapy on expression of angio-and fibrogenic factors, and cytokine concentrations during the healing process of human pressure ulcers. Int. J. Med. Sci. 2018, 15, 1105. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Sun, Z.; Zhang, L.; Li, X.; Dong, Y.; Liu, T.C.-Y. Effects of low-level laser therapy on ROS homeostasis and expression of IGF-1 and TGF-β1 in skeletal muscle during the repair process. Lasers Med. Sci. 2013, 28, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Saygun, I.; Nizam, N.; Ural, A.U.; Serdar, M.A.; Avcu, F.; Tozum, T. Low-level laser irradiation affects the release of basic fibroblast growth factor (bFGF), insulin-like growth factor-I (IGF-I), and receptor of IGF-I (IGFBP3) from osteoblasts. Photomed. Laser Surg. 2012, 30, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, N.; Mayahara, K.; Kiyosaki, T.; Yamaguchi, A.; Ozawa, Y.; Abiko, Y. Low-intensity laser irradiation stimulates bone nodule formation via insulin-like growth factor-I expression in rat calvarial cells. Lasers Surg. Med. 2007, 39, 551–559. [Google Scholar] [CrossRef]
- Gaptulbarova, K.A.; Tsyganov, M.M.; Pevzner, A.M.; Ibragimova, M.K.; Litviakov, N.V. NF-kB as a potential prognostic marker and a candidate for targeted therapy of cancer. Exp. Oncol 2020, 42, 263–269. [Google Scholar] [CrossRef]
- Thoma, A.; Lightfoot, A.P. NF-kB and Inflammatory Cytokine Signalling: Role in Skeletal Muscle Atrophy. Muscle Atrophy 2018, 1088, 267–279. [Google Scholar] [CrossRef]
- Rizzi, C.F.; Mauriz, J.L.; Freitas Correa, D.S.; Moreira, A.J.; Zettler, C.G.; Filippin, L.I.; Marroni, N.P.; Gonzalez-Gallego, J. Effects of low-level laser therapy (LLLT) on the nuclear factor (NF)-κB signaling pathway in traumatized muscle. Lasers Surg. Med. 2006, 38, 704–713. [Google Scholar] [CrossRef]
- Tamura, A.; Matsunobu, T.; Tamura, R.; Kawauchi, S.; Sato, S.; Shiotani, A. Photobiomodulation rescues the cochlea from noise-induced hearing loss via upregulating nuclear factor κB expression in rats. Brain Res. 2016, 1646, 467–474. [Google Scholar] [CrossRef]
- Yin, K.; Zhu, R.; Wang, S.; Zhao, R.C. Low level laser (LLL) attenuate LPS-induced inflammatory responses in mesenchymal stem cells via the suppression of NF-κB signaling pathway in vitro. PLoS ONE 2017, 12, e0179175. [Google Scholar] [CrossRef]
- Gupta, A.; Keshri, G.K.; Yadav, A.; Gola, S.; Chauhan, S.; Salhan, A.K.; Singh, S.B. Superpulsed (Ga-As, 904 nm) low-level laser therapy (LLLT) attenuates inflammatory response and enhances healing of burn wounds. J. Biophotonics 2015, 8, 489–501. [Google Scholar] [CrossRef]
- Cury, V.; Moretti, A.I.S.; Assis, L.; Bossini, P.; Crusca, J.D.S.; Neto, C.B.; Fangel, R.; de Souza, H.P.; Hamblin, M.; Parizotto, N.A. Low level laser therapy increases angiogenesis in a model of ischemic skin flap in rats mediated by VEGF, HIF-1α and MMP-2. J. Photochem. Photobiol. B Biol. 2013, 125, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xu, Q.; Shi, M.; Gan, P.; Huang, Q.; Wang, A.; Tan, G.; Fang, Y.; Liao, H. Low-level laser therapy induces human umbilical vascular endothelial cell proliferation, migration and tube formation through activating the PI3K/Akt signaling pathway. Microvasc. Res. 2020, 129, 103959. [Google Scholar] [CrossRef] [PubMed]
- Rhee, Y.-H.; Moon, J.-H.; Choi, S.-H.; Ahn, J.-C. Low-level laser therapy promoted aggressive proliferation and angiogenesis through decreasing of transforming growth factor-β1 and Increasing of Akt/hypoxia inducible factor-1α in anaplastic thyroid cancer. Photomed. Laser Surg. 2016, 34, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Zou, Z.; Li, J.; Shen, Q.; Liu, L.; An, X.; Yang, S.; Xing, D. Photoactivation of mitochondrial reactive oxygen species-mediated Src and protein kinase C pathway enhances MHC class II-restricted T cell immunity to tumours. Cancer Lett. 2021, 523, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Ambasta, R.K.; Kumar, P. Autophagy and apoptosis cascade: Which is more prominent in neuronal death? Cell. Mol. Life Sci. 2021, 78, 8001–8047. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Chen, T.; Xing, D.; Wang, F.; Pei, Y.; Wei, X. Single cell analysis of PKC activation during proliferation and apoptosis induced by laser irradiation. J. Cell. Physiol. 2006, 206, 441–448. [Google Scholar] [CrossRef]
- Wang, F.; Chen, T.-S.; Xing, D.; Wang, J.-J.; Wu, Y.-X. Measuring dynamics of caspase-3 activity in living cells using FRET technique during apoptosis induced by high fluence low-power laser irradiation. Lasers Surg. Med. 2005, 36, 2–7. [Google Scholar] [CrossRef]
- Izadi, M.; Ali, T.A.; Pourkarimi, E. Over Fifty Years of Life, Death, and Cannibalism: A Historical Recollection of Apoptosis and Autophagy. Int. J. Mol. Sci. 2021, 22, 12466. [Google Scholar] [CrossRef]
- Martines-Arano, H.; Palacios-Barreto, S.; Castillo-Cruz, J.; Meda-Campaña, J.A.; García-Pérez, B.E.; Torres-Torres, C. Fractional photodamage triggered by chaotic attractors in human lung epithelial cancer cells. Int. J. Therm. Sci. 2022, 181, 107734. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rola, P.; Włodarczak, S.; Lesiak, M.; Doroszko, A.; Włodarczak, A. Changes in Cell Biology under the Influence of Low-Level Laser Therapy. Photonics 2022, 9, 502. https://doi.org/10.3390/photonics9070502
Rola P, Włodarczak S, Lesiak M, Doroszko A, Włodarczak A. Changes in Cell Biology under the Influence of Low-Level Laser Therapy. Photonics. 2022; 9(7):502. https://doi.org/10.3390/photonics9070502
Chicago/Turabian StyleRola, Piotr, Szymon Włodarczak, Maciej Lesiak, Adrian Doroszko, and Adrian Włodarczak. 2022. "Changes in Cell Biology under the Influence of Low-Level Laser Therapy" Photonics 9, no. 7: 502. https://doi.org/10.3390/photonics9070502
APA StyleRola, P., Włodarczak, S., Lesiak, M., Doroszko, A., & Włodarczak, A. (2022). Changes in Cell Biology under the Influence of Low-Level Laser Therapy. Photonics, 9(7), 502. https://doi.org/10.3390/photonics9070502