Single-Molecule Clustering for Super-Resolution Optical Fluorescence Microscopy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
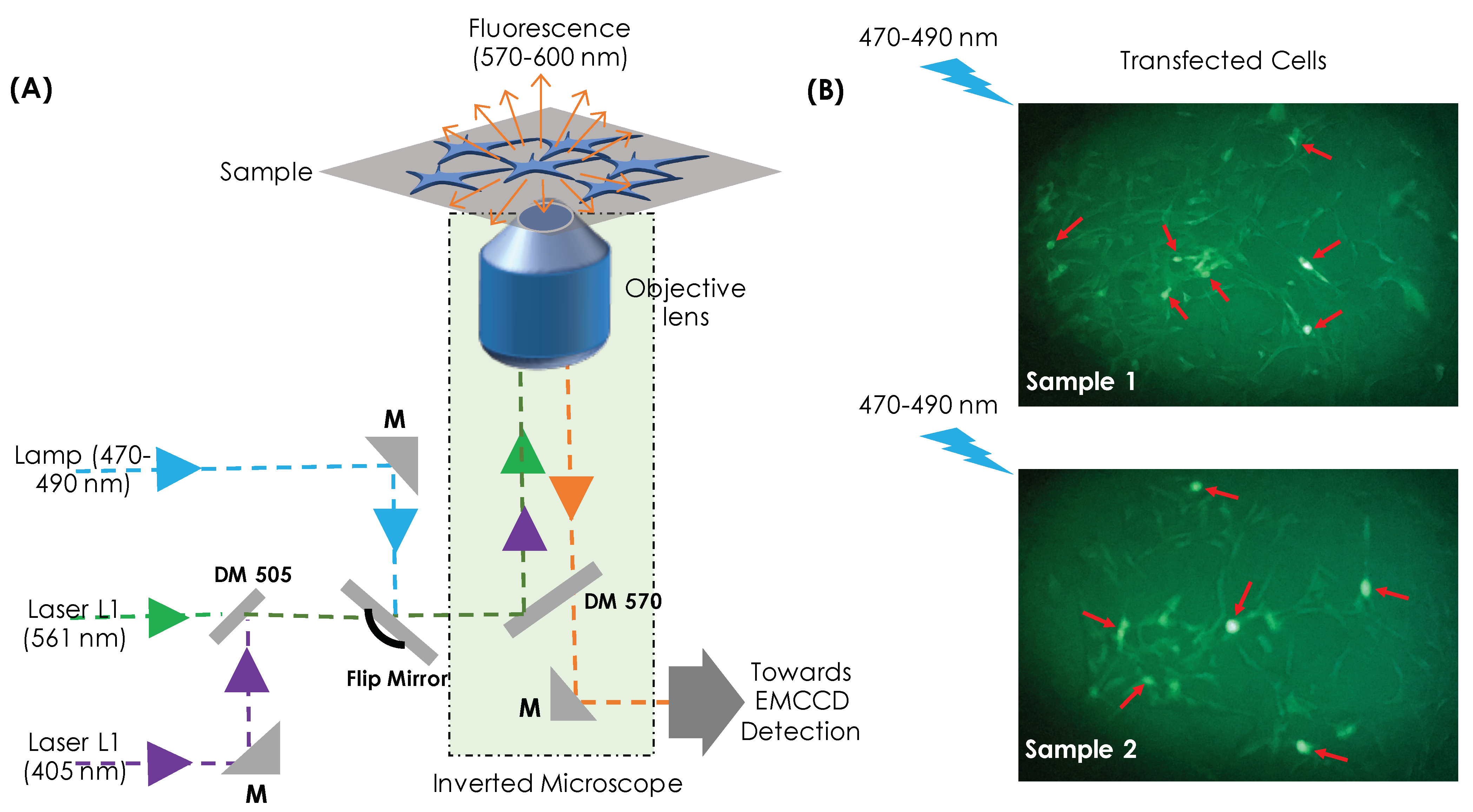
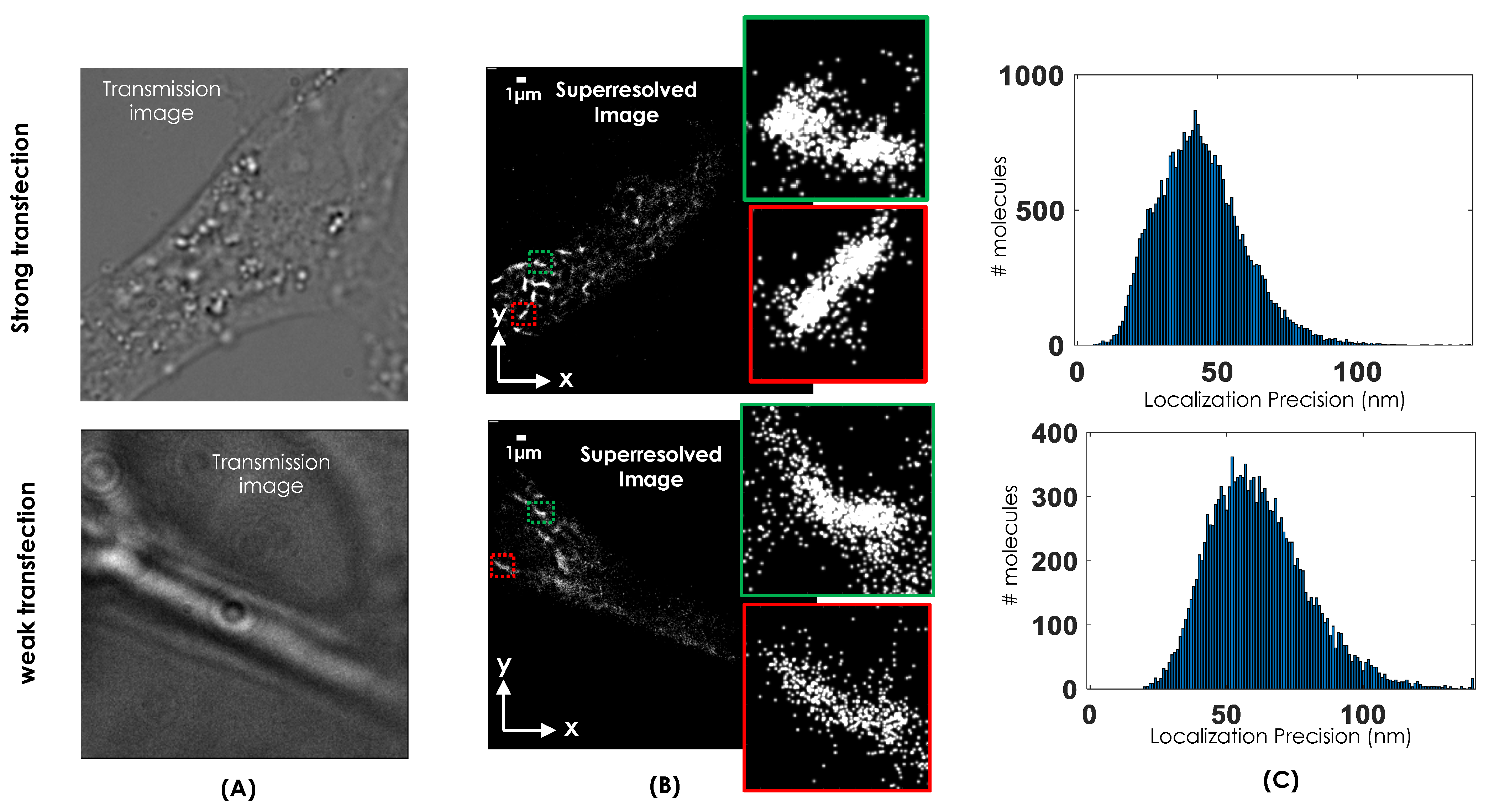
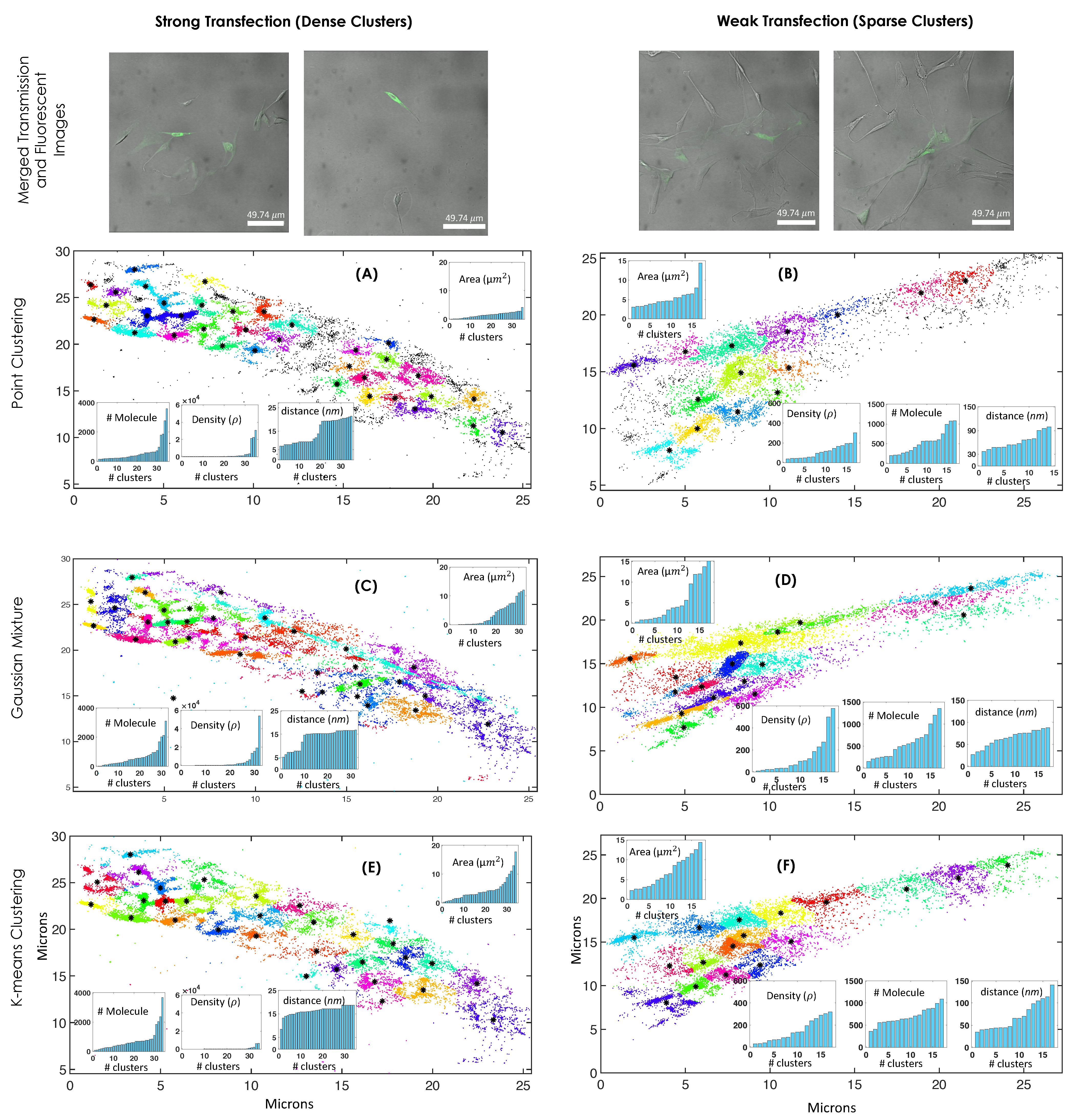
2. Results
3. Discussion
4. Methods and Protocols
4.1. Cell Culture
4.2. Cell Transfection and Fixing
4.3. Optical Setup
4.4. Superresolution Imaging and Image Reconstruction
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Platzer, R.; Rossboth, B.K.; Schneider, M.C. Unscrambling fluorophore blinking for comprehensive cluster detection via photoactivated localization microscopy. Nat. Commun. 2020, 11, 4993. [Google Scholar] [CrossRef]
- Gudheti, M.V.; Curthoys, N.M.; Gould, T.J.; Kim, D.; Gunewardene, M.S.; Gabor, K.A.; Gosse, J.A.; Kim, C.H.; Zimmerberg, J.; Hess, S.T. Actin mediates the nanoscale membrane organization of the clustered membrane protein influenza hemagglutinin. Biophys. J. 2013, 104, 2182–2192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Civitci, F.; Shangguan, J.; Zheng, T. Fast and multiplexed superresolution imaging with DNA-PAINT-ERS. Nat. Commun. 2020, 11, 4339. [Google Scholar] [CrossRef] [PubMed]
- Curthoys, N.M.; Mlodzianoski, M.J.; Parent, M.; Butler, M.B.; Raut, P.; Wallace, J.; Lilieholm, J.; Mehmood, K.; Maginnis, M.S.; Waters, H.; et al. Influenza Hemagglutinin Modulates Phosphatidylinositol 4, 5-Bisphosphate Membrane Clustering. Biophys. J. 2019, 116, 893–909. [Google Scholar] [CrossRef] [Green Version]
- Deniz, A.A.; Mukhopadhyay, S.; Lemke, E.A. Single-molecule biophysics: At the interface of biology, physics and chemistry. J. R. Soc. Interface 2008, 5, 15–45. [Google Scholar] [CrossRef]
- Mondal, P.P. Probabilistic Optically-Selective Single-molecule Imaging Based Localization Encoded (POSSIBLE) microscopy for ultra-superresolution imaging. PLoS ONE 2020, 15, e0242452. [Google Scholar]
- Yasui, M.; Hiroshima, M.; Kozuka, J.; Sako, Y.; Ueda, M. Automated single-molecule imaging in living cells. Nat. Commun. 2018, 9, 3061. [Google Scholar] [CrossRef] [PubMed]
- Gustavsson, A.K.; Petrov, P.N.; Lee, M.Y.; Shechtman, Y.; Moerner, W.E. 3D single-molecule super-resolution microscopy with a tilted light sheet. Nat. Commun. 2018, 9, 123. [Google Scholar] [CrossRef] [Green Version]
- Mondal, P.P.; Hess, S.T. Total internal reflection fluorescence based multiplane localization microscopy enables super-resolved volume imaging. Appl. Phys. Lett. 2017, 110, 211102. [Google Scholar] [CrossRef]
- Greiss, F.; Deligiannaki, M.; Jung, C.; Gaul, U.; Braun, D. Single-Molecule Imaging in Living Drosophila Embryos with Reflected Light-Sheet Microscopy. Biophys. J. 2016, 110, 939–946. [Google Scholar] [CrossRef] [Green Version]
- Andrecka, J.; Arroyo, J.O.; Lewis, K.; Cross, R.A.; Kukura, P. Label-free Imaging of Microtubules with Sub-nm Precision Using Interferometric Scattering Microscopy. Biophys. J. 2016, 110, 214–217. [Google Scholar] [CrossRef] [Green Version]
- Luo, F.; Qin, G.; Xia, T.; Fang, X. Single-Molecule Imaging of Protein Interactions and Dynamics. Ann. Rev. Anal. Chem. 2020, 13, 337. [Google Scholar] [CrossRef] [PubMed]
- Moerner, W.E.; Kador, L. Optical detection and spectroscopy of single molecules in a solid. Phys. Rev. Lett. 1989, 62, 2535. [Google Scholar] [CrossRef] [Green Version]
- Hess, S.T.; Girirajan, T.P.; Mason, M.D. Ultra-high resolution imaging by fluorescence photoactivation localization microscopy. Biophys. J. 2006, 91, 4258–4272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valles, M.; Hess, S.T. A Cross Beam Excitation Geometry for Localization Microscopy. iSci. Notes 2017, 2, 1. [Google Scholar]
- Hell, S.W.; Wichmann, J. Breaking the diffraction resolution limit by stimulated emission: Stimulated-emission-depletion fluorescence microscopy. Opt. Lett. 1994, 19, 780–782. [Google Scholar] [CrossRef] [PubMed]
- Rust, M.; Bates, M.; Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy (STORM). Nat. Methods 2006, 3, 793–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betzig, E.; Patterson, G.H.; Sougrat, R.; Lindwasser, O.W.; Olenych, S.; Bonifacino, J.S.; Davidson, M.W.; Lippincott-Schwartz, J.; Hess, H.F. Imaging intracellular fluorescent proteins at nanometer resolution. Science 2006, 313, 1642–1645. [Google Scholar] [CrossRef] [Green Version]
- Gustafsson, M.G.L. Nonlinear structured-illumination microscopy: Wide-field fluorescence imaging with theoretically unlimited resolution. Proc. Natl. Acad. Sci. USA 2005, 102, 13081–13086. [Google Scholar] [CrossRef] [Green Version]
- Cella Zanacchi, F.; Lavagnino, Z.; Perrone Donnorso, M.; Del Bue, A.; Furia, L.; Faretta, M.; Diaspro, A. Live-cell 3D super-resolution imaging in thick biological samples. Nat. Methods 2011, 8, 1047–1049. [Google Scholar] [CrossRef]
- Fölling, J.; Bossi, M.; Bock, H.; Medda, R.; Wurm, C.A.; Hein, B.; Jakobs, S.; Eggeling, C.; Hell, S.W. Fluorescence nanoscopy by ground-state depletion and single-molecule return. Nat. Methods 2008, 5, 943–945. [Google Scholar] [CrossRef] [PubMed]
- Dertinger, T.; Colyer, R.; Iyer, G.; Weiss, S.; Enderlein, J. Fast, background-free, 3D super-resolution optical fluctuation imaging (SOFI). Proc. Natl. Acad. Sci. USA 2009, 106, 22287–22292. [Google Scholar] [CrossRef] [Green Version]
- Sharonov, A.; Hochstrasser, R.M. Wide-field subdiffraction imaging by accumulated binding of diffusing probes Alexey Sharonov and Robin M. Hochstrasser. Proc. Natl. Acad. Sci. USA 2006, 103, 18911–18916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannone, G.; Hosy, E.; Levet, F.; Constals, A.; Schulze, K.; Sobolevsky, A.I.; Rosconi, M.P.; Gouaux, E.; Tampé, R.; Choquet, D.; et al. Dynamic superresolution imaging of endogenous proteins on living cells at ultra-high density. Biophys. J. 2010, 99, 1303–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, P.P. Simultaneous multiplane imaging-based localization encoded (SMILE) microscopy for super-resolution volume imaging. Microsc. Res. Tech. 2017, 80, 333. [Google Scholar] [CrossRef]
- Gwosch, K.C.; Pape, J.K.; Balzarotti, F.; Hoess, P.; Ellenberg, J.; Ries, J.; Hell, S.W. MINFLUX nanoscopy delivers 3D multicolor nanometer resolution in cells. Nat. Methods 2020, 217–224. [Google Scholar] [CrossRef]
- Zhang, Y.; Schroeder, L.K.; Lessard, M.D.; Kidd, P.; Chung, J.; Song, Y.; Benedetti, L.; Li, Y.; Ries, J.; Grimm, J.B.; et al. Nanoscale subcellular architecture revealed by multicolor three-dimensional salvaged fluorescence imaging. Nat. Methods 2020, 17, 225–231. [Google Scholar] [CrossRef]
- AbuZineh, K.; Joudeh, L.I.; Al Alwan, B.; Hamdan, S.M.; Merzaban, J.S.; Habuchi, S. Microfluidics-based super-resolution microscopy enables nanoscopic characterization of blood stem cell rolling. Sci. Adv. 2018, 4, eaat5304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mlodzianoski, M.J. Future Considerations for Localizationbased Super-resolution Fluorescence Microscopy. iSci. NoteS 2016, 1, 5. [Google Scholar]
- Thevathasan, J.V.; Kahnwald, M.; Cieśliński, K.; Hoess, P.; Peneti, S.K.; Reitberger, M.; Heid, D.; Kasuba, K.C.; Hoerner, S.J.; Li, Y.; et al. Nuclear pores as versatile reference standards for quantitative superresolution microscopy. Nat. Methods 2019, 16, 1045–1053. [Google Scholar] [CrossRef]
- Habuchi, S. Microfluidics-based Super-Resolution Imaging: A New Tool for Nanoscopic Charactertization of Cellular Interactions. iSci. Notes 2019, 4, 1. [Google Scholar] [CrossRef]
- Ma, H.; Xu, J.; Liu, Y. WindSTORM: Robust online image processing for high-throughput nanoscopy. Sci. Adv. 2019, 5, eaaw0683. [Google Scholar] [CrossRef] [Green Version]
- Turcotte, R.; Liang, Y.; Tanimoto, M.; Zhang, Q.; Li, Z.; Koyama, M.; Betzig, E.; Ji, N. Dynamic super-resolution structured illumination imaging in the living brain. Proc. Natl. Acad. Sci. USA 2019, 116, 9586–9591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Taki, M.; Sato, Y.; Tamura, Y.; Yaginuma, H.; Okada, Y.; Yamaguchi, S. A photostable fluorescent marker for the superresolution live imaging of the dynamic structure of the mitochondrial cristae. Proc. Natl. Acad. Sci. USA 2019, 116, 15817–15822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chlanda, P.; Zimmerberg, J. Protein–lipid interactions critical to replication of the influenza A virus. FEBS Lett. 2016, 590, 1940. [Google Scholar] [CrossRef] [Green Version]
- Hess, S.T.; Gould, T.J.; Gudheti, M.V.; Maas, S.A.; Mills, K.D.; Zimmerberg, J. Dynamic clustered distribution of hemagglutinin resolved at 40 nm in living cell membranes discriminates between raft theories. Proc. Natl. Acad. Sci. USA 2007, 104, 17370–17375. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, R.; Blanco, M.R.; Kahlscheuer, M.L.; Abelson, J.; Guthrie, C.; Walter, N.G. Biased Brownian ratcheting leads to pre-mRNA remodeling and capture prior to first-step splicing. Nat. Struct. Mol. Biol. 2013, 20, 1450–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voicu, A.; Duteanu, N.; Voicu, M.; Vlad, D.; Dumitrascu, V. The rcdk and cluster R packages applied to drug candidate selection. J. Cheminform. 2020, 12, 3. [Google Scholar] [CrossRef] [Green Version]
- Lemmer, M.; Inkpen, M.S.; Kornysheva, K.; Long, N.J.; Albrecht, T. Unsupervised vector-based classification of single-molecule charge transport data. Nat. Commun. 2016, 7, 12922. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Park, S.; Challapalli, S.D.; Fei, J.; Wang, Y. Robust nonparametric quantification of clustering density of molecules in single-molecule localization microscopy. PLoS ONE 2017, 12, e0179975. [Google Scholar] [CrossRef] [Green Version]
- Ripley, B.D. Modelling spatial patterns. J. R. Stat. Soc. Ser. B Methodol. 1977, 39, 172–192. [Google Scholar] [CrossRef]
- Kiskowski, M.A.; Hancock, J.F.; Kenworthy, A.K. On the use of Ripley’s K-function and its derivatives to analyze domain size. Biophys. J. 2009, 97, 1095–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ester, M.; Kriegel, H.P.; Sander, J.; Xu, X. A density-based algorithm for discovering clusters in large spatial databases with noise. In Proceedings of the Second International Conference on Knowledge Discovery and Data Mining (KDD-96), Portland, OR, USA, 2–4 August 1996; pp. 226–231. [Google Scholar]
- Parent, M.; Hess, S.T. Quantification of Mitochondrial Membrane Curvature by Three-Dimensional Localization Microscopy. iSci. Notes 2019, 4, 3. [Google Scholar] [CrossRef]
- Mondal, P.P. Spatio-Temporal Super-resolution Optical Fluorescence Microscopy. iSci. Notes 2016, 1, 4. [Google Scholar] [CrossRef]
- Joshi, P.; Mondal, P.P. Adaptable single molecule localization microscopy (aSMLM) for superresolution optical fluorescence imaging. Appl. Phys. Lett. 2021, 119, 173703. [Google Scholar] [CrossRef]
- Thompson, R.E.; Larson, D.R.; Webb, W.W. Precise nanometer localization analysis for individual fluorescent probes. Biophys. J. 2002, 82, 2775–2783. [Google Scholar] [CrossRef] [Green Version]
- Veit, M.; Serebryakova, M.V.; Kordyukova, L.V. Palmitoylation of influenza virus proteins. Biochem. Soc. Trans. 2013, 41, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Veit, M.; Kretzschmar, E.; Kuroda, K.; Garten, W.; Schmidt, M.F.; Klenk, H.D.; Rott, R. Site-specific mutagenesis identifies three cysteine residues in the cytoplasmic tail as acylation sites of influenza virus hemagglutinin. J. Virol. 1991, 65, 2491–2500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ankerst, M.; Breunig, M.M.; Kriegel, H.P.; Sander, J. OPTICS: Ordering points to identify the clustering structure. In Proceedings of the 1999 ACM SIGMOD International Conference on Management of Data, Philadelphia, PA, USA, 31 May–3 June 1999. [Google Scholar]
- Marcon, Y. Distance-Based Clustering of a Set of XY Coordinates. MATLAB Central File Exchange. 2021. Available online: https://www.mathworks.com/matlabcentral/fileexchange/56150-distance-based-clustering-of-a-set-of-xy-coordinates (accessed on 22 October 2021).
- McLachlan, G.; Peel, D. Finite Mixture Models; John Wiley and Sons, Inc.: Hoboken, NJ, USA, 2000. [Google Scholar]
- Richardson, W.H. Bayesian-Based Iterative Method of Image Restoration. J. Opt. Soc. Am. 1972, 62, 55–59. [Google Scholar] [CrossRef]
- Lucy, L.B. An iterative technique for the rectification of observed distributions. Astron. J. 1974, 79, 745. [Google Scholar] [CrossRef] [Green Version]
- Mondal, P.P.; Vicidomini, G.; Diaspro, A. Image reconstruction for multiphoton fluorescence microscopy. Appl. Phys. Lett. 2008, 92, 103902. [Google Scholar] [CrossRef]
- Mondal, P.P. Markov random field aided Bayesian approach for image reconstruction in confocal microscopy. J. Appl. Phys. 2007, 102, 044701. [Google Scholar] [CrossRef]
- Arthur, D.; Vassilvitskii, S. k-means++: The advantages of careful seeding. In Proceedings of the Eighteenth Annual ACM-SIAM Symposium on Discrete Algorithms, New Orleans, LA, USA, 7–9 January 2007; p. 1027. [Google Scholar]
- Lloyd, S.P. Least squares quantization in PCM. IEEE Trans. Inf. Theory 1982, 28, 129. [Google Scholar] [CrossRef]
- Fraix-Burnet, D.; Thuillard, M.; Chattopadhyay, A.K. Multivariate approaches to classification in extragalactic astronomy. Front. Astron. Space Sci. 2015, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Perez-Ortega, J.; Almanza-Ortega, N.N.; Romero, D. Balancing effort and benefit of K-means clustering algorithms in Big Data realms. PLoS ONE 2018, 13, e0201874. [Google Scholar] [CrossRef] [PubMed]
- Okawa, S.; Yano, A.; Uchida, K.; Mitsui, Y.; Yoshida, M.; Takekoshi, M.; Marjono, A.; Gao, F.; Hoshi, Y.; Kida, I.; et al. Phantom and mouse experiments of time-domain fluorescence tomography using total light approach. Biomed. Opt. Exp. 2013, 4, 635–651. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Gaoa, M.; Papadakis, G.Z.; Luna, B.; Jain, S.; Mollura, D.J.; Bagci, U. Joint solution for PET image segmentation, denoising, and partial volume correction. Med. Image Anal. 2018, 46, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Dou, D.; Revol, R.; Östbye, H.; Wang, H.; Daniels, R. Influenza A Virus Cell Entry, Replication, Virion Assembly and Movement. Front Immunol. 2018, 9, 1581. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joshi, P.; Mondal, P.P. Single-Molecule Clustering for Super-Resolution Optical Fluorescence Microscopy. Photonics 2022, 9, 7. https://doi.org/10.3390/photonics9010007
Joshi P, Mondal PP. Single-Molecule Clustering for Super-Resolution Optical Fluorescence Microscopy. Photonics. 2022; 9(1):7. https://doi.org/10.3390/photonics9010007
Chicago/Turabian StyleJoshi, Prakash, and Partha Pratim Mondal. 2022. "Single-Molecule Clustering for Super-Resolution Optical Fluorescence Microscopy" Photonics 9, no. 1: 7. https://doi.org/10.3390/photonics9010007
APA StyleJoshi, P., & Mondal, P. P. (2022). Single-Molecule Clustering for Super-Resolution Optical Fluorescence Microscopy. Photonics, 9(1), 7. https://doi.org/10.3390/photonics9010007