
Analysis of Unusual Sulfated Constituents and Anti-infective Properties of Two Indonesian Mangroves, Lumnitzera littorea and Lumnitzera racemosa (Combretaceae)

 , ,
, ,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Root Sample Extraction
2.3. General Experimental Procedures
2.4. UHPLC-ESI-QqTOF-MS and MS/MS
2.5. RP-UHPLC-ESI-LIT-Orbitrap-MS
2.6. Extraction and Isolation
2.7. DNA Extraction, Polymerase Chain Reaction, and Sequencing
2.8. Phylogenetic Analyses
2.9. Anti-infective Bioassays
3. Results and Discussion
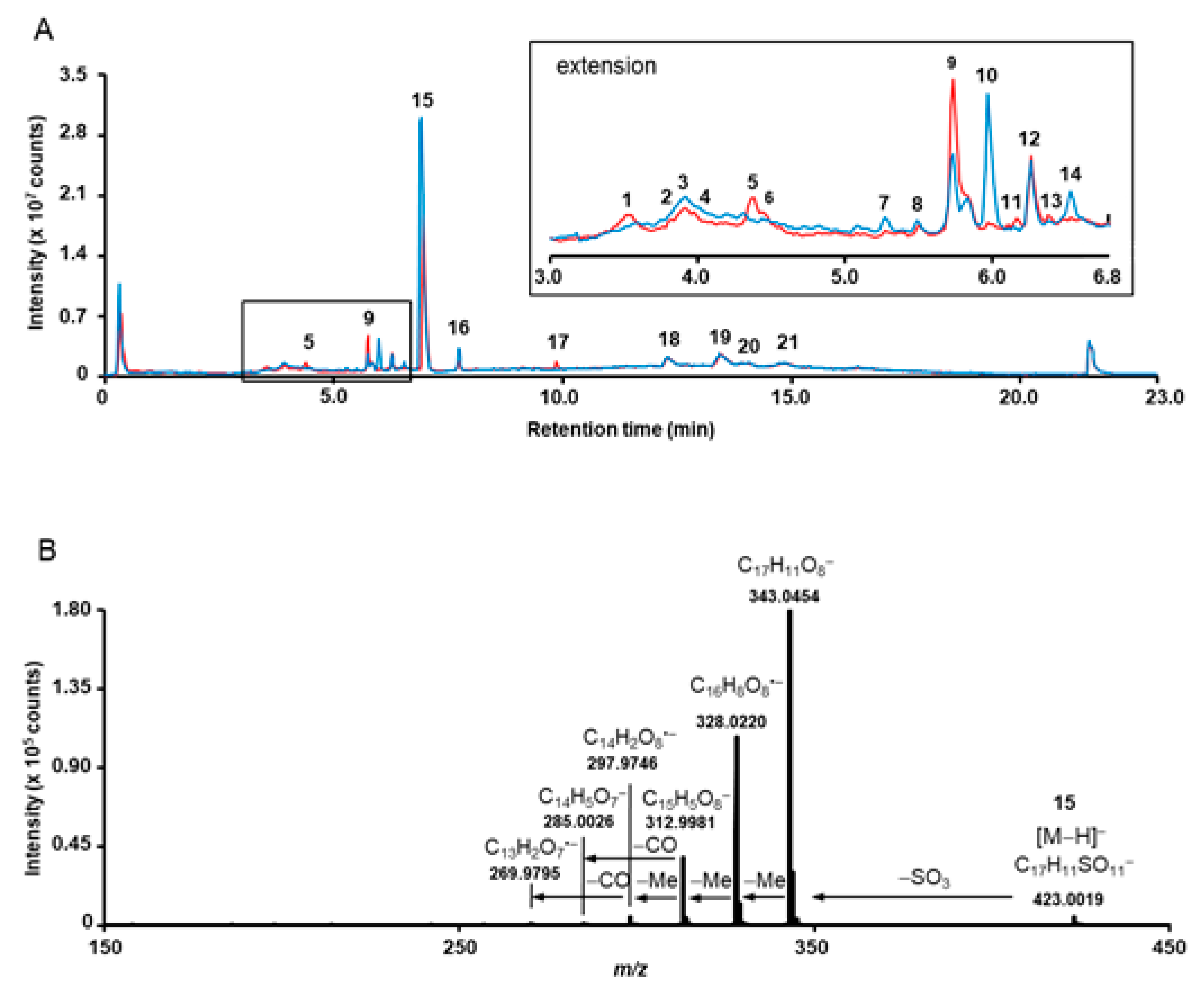
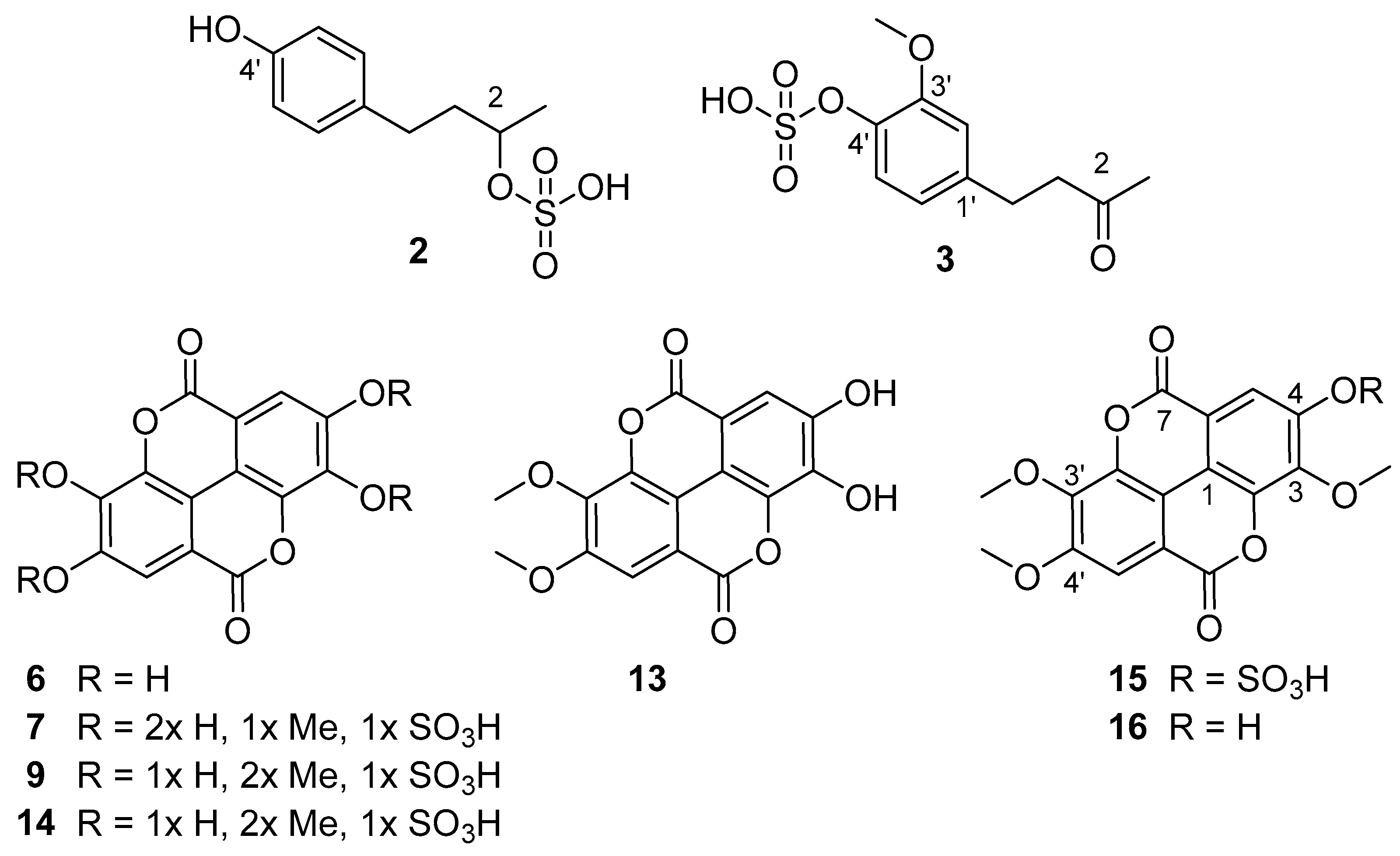
3.1. Phytochemcial Analyses
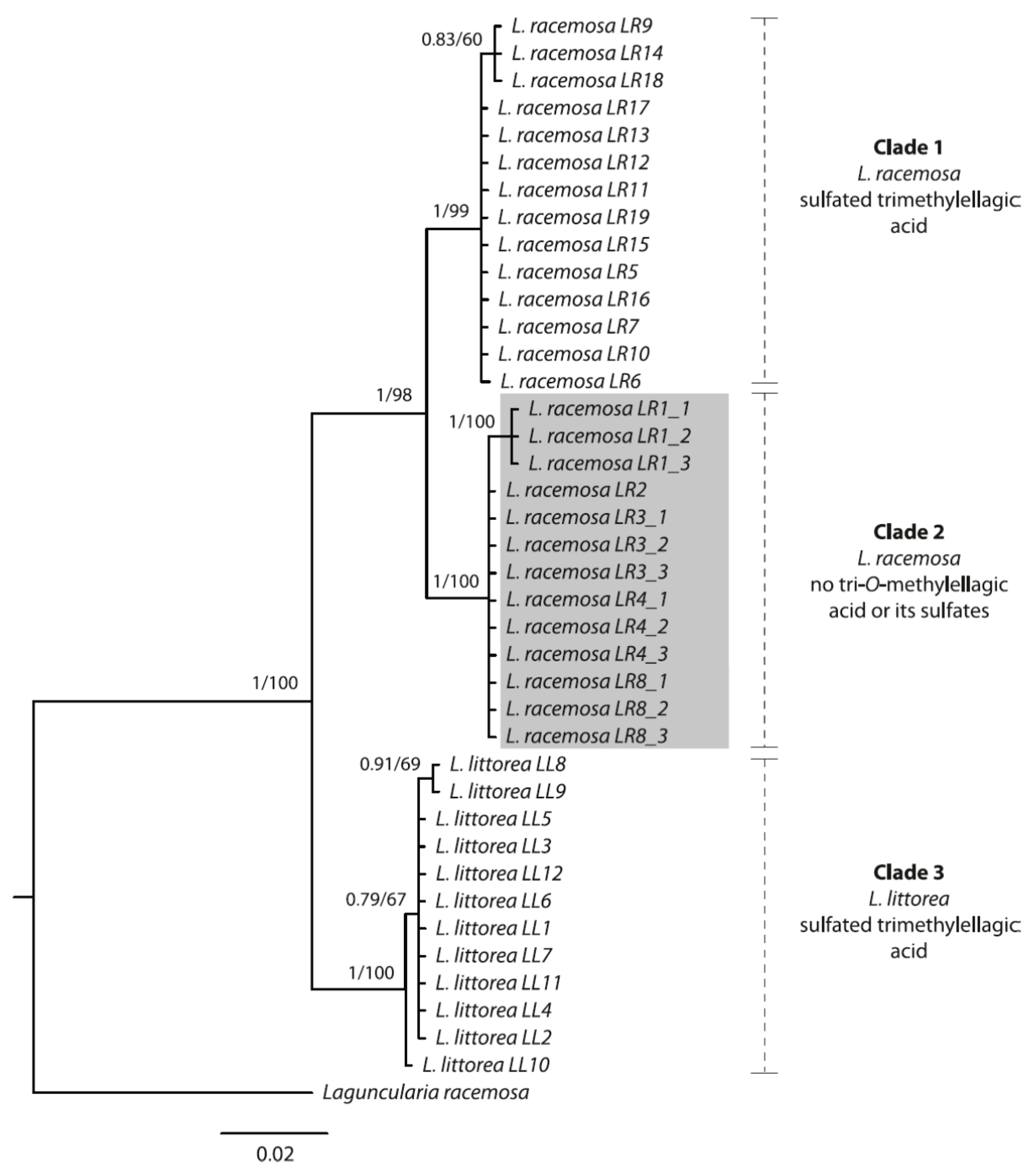
3.2. Phylogenetic Analyses
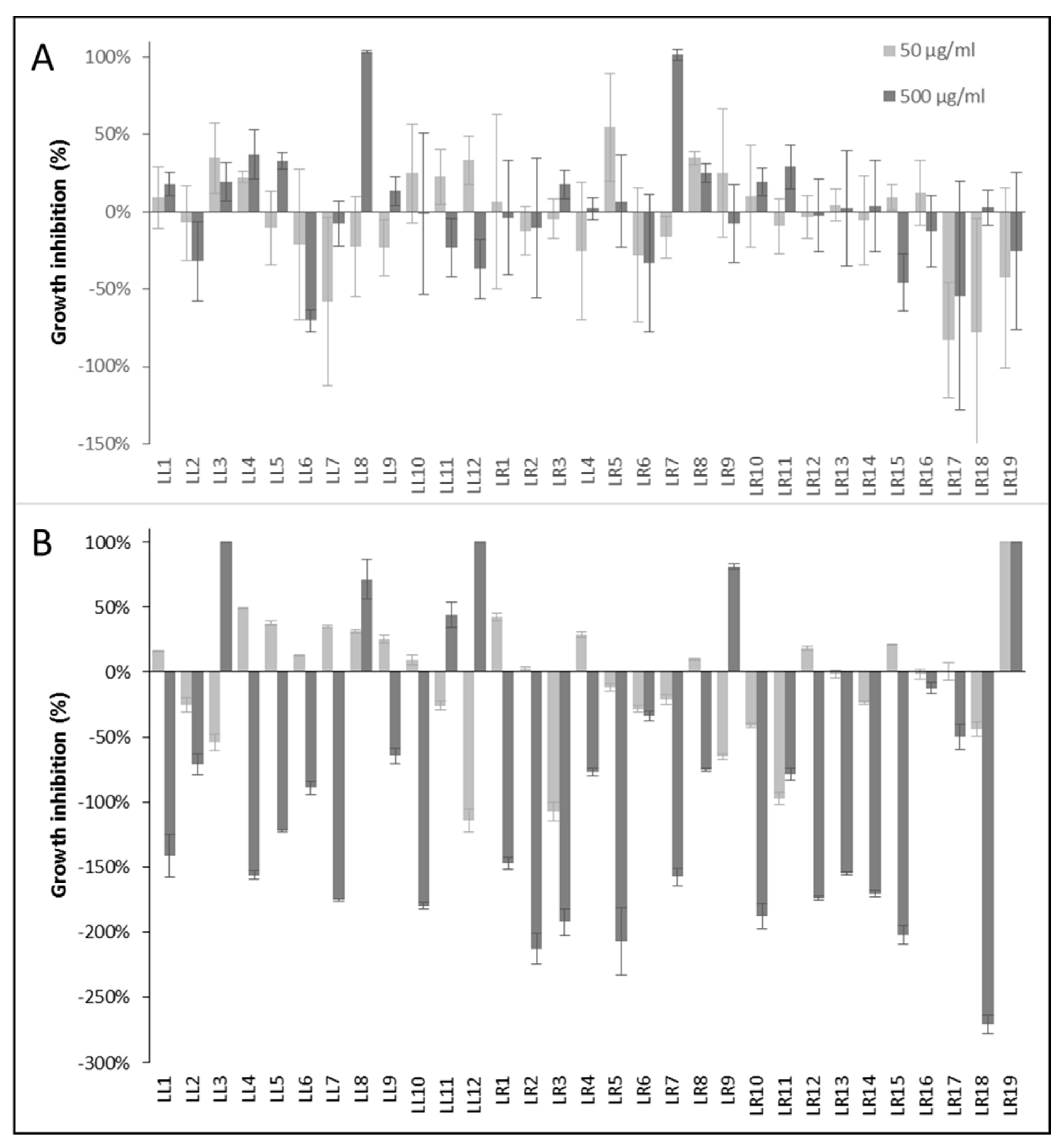
3.3. Evaluation of Anti-Infective Properties
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kathiresan, K.; Bingham, B.L. Biology of mangroves and mangrove ecosystems. Adv. Mar. Biol. 2001, 40, 81–251. [Google Scholar]
- Noor, Y.R.; Khazali, M.; Suryadiputra, I.N.N. Panduan Pengenalan Mangrove di Indonesia; Ditjen PHKA; Wetlands International Indonesia Programme: Bogor, India, 2006; ISBN 9799589908. [Google Scholar]
- Giesen, W.; Wulffraat, S.; Zieren, M.; Scholten, L. Mangrove Guidebook for Southeast Asia; FAO Regional Office for Asia and the Pacific, Wetlands International: Bangkok, Thailand, 2007; ISBN 974-7946-85-8. [Google Scholar]
- Bandaranayake, W.M. Traditional and medicinal uses of mangroves. Mangroves Salt Marshes 1998, 2, 133–148. [Google Scholar] [CrossRef]
- Kovacs, J.M. Assessing mangrove use at the local scale. Landsc. Urban Plan. 1999, 43, 201–208. [Google Scholar] [CrossRef]
- Pattanaik, C.; Reddy, C.S.; Dhal, N.K.; Das, R. Utilisation of mangrove forests in Bhitarkanika wildlife sanctuary, Orissa. Indian J. Tradit. Knowl. 2008, 7, 598–603. [Google Scholar]
- Govindasamy, C.; Kannan, R. Pharmacognosy of mangrove plants in the system of unani medicine. Asian Pac. J. Trop. Dis. 2012, 2, S38–S41. [Google Scholar] [CrossRef]
- Balk, M.; Keuskamp, J.A.; Laanbroek, H.J. Potential for sulfate reduction in mangrove forest soils: Comparison between two dominant species of the Americas. Front. Microbiol. 2016, 7, 1855. [Google Scholar] [CrossRef]
- Spalding, M.; Blasco, F.; Field, C.D. World Mangrove Atlas; International Society for Mangrove Ecosystems: Okinawa, Japan; World Conservation Monitoring Centre: Cambridge, UK; International Tropical Timber Organization: Yokohama, Japan, 1997; ISBN 4906584039. [Google Scholar]
- Giri, C.; Ochieng, E.; Tieszen, L.L.; Zhu, Z.; Singh, A.; Loveland, T.; Masek, J.; Duke, N. Status and distribution of mangrove forests of the world using earth observation satellite data. Glob. Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- Fyhrquist, P.; Mwasumbi, L.; Hæggström, C.-A.; Vuorela, H.; Hiltunen, R.; Vuorela, P. Ethnobotanical and antimicrobial investigation on some species of Terminalia and Combretum (Combretaceae) growing in Tanzania. J. Ethnopharmacol. 2002, 79, 169–177. [Google Scholar] [CrossRef]
- Martini, N.D.; Katerere, D.R.P.; Eloff, J.N. Biological activity of five antibacterial flavonoids from Combretum erythrophyllum (Combretaceae). J. Ethnopharmacol. 2004, 93, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Eloff, J.N.; Famakin, J.O.; Katerere, D.R.P. Isolation of an antibacterial stilbene from Combretum woodii (Combretaceae) leaves. Afr. J. Biotechnol. 2005, 4, 1167–1171. [Google Scholar]
- Eloff, J.N.; Famakin, J.O.; Katerere, D.R.P. Combretum woodii (Combretaceae) leaf extracts have high activity against Gram-negative and Gram-positive bacteria. Afr. J. Biotechnol. 2005, 4, 1161–1166. [Google Scholar]
- Eloff, J.N.; Katerere, D.R.; McGaw, L.J. The biological activity and chemistry of the southern African Combretaceae. J. Ethnopharmacol. 2008, 119, 686–699. [Google Scholar] [CrossRef]
- Aderogba, M.A.; Kgatle, D.T.; McGaw, L.J.; Eloff, J.N. Isolation of antioxidant constituents from Combretum apiculatum subsp. apiculatum. S. Afr. J. Bot. 2012, 79, 125–131. [Google Scholar] [CrossRef]
- Masoko, P.; Eloff, J.N. The diversity of antifungal compounds of six South African Terminalia species (Combretaceae) determined by bioautography. Afr. J. Biotechnol. 2005, 4, 1425–1431. [Google Scholar]
- Masoko, P.; Picard, J.; Eloff, J.N. The antifungal activity of twenty-four southern African Combretum species (Combretaceae). S. Afr. J. Bot. 2007, 73, 173–183. [Google Scholar] [CrossRef]
- Lin, T.-C.; Hsu, F.-L.; Cheng, J.-T. Antihypertensive Activity of Corilagin and Chebulinic Acid, Tannins from Lumnitzera racemosa. J. Nat. Prod. 1993, 56, 629–632. [Google Scholar] [CrossRef]
- Ravikumar, S.; Gnanadesigan, M. Hepatoprotective and antioxidant activity of a mangrove plant Lumnitzera racemosa. Asian Pac. J. Trop. Biomed. 2011, 1, 348–352. [Google Scholar] [CrossRef]
- Thao, N.P.; Bui, T.T.L.; Chau, N.D.; Bui, H.T.; Kim, E.J.; Kang, H.K.; Lee, S.H.; Jang, H.D.; Nguyen, T.C.; van Nguyen, T.; et al. In vitro evaluation of the antioxidant and cytotoxic activities of constituents of the mangrove Lumnitzera racemosa Willd. Arch. Pharm. Res. 2015, 38, 446–455. [Google Scholar] [CrossRef]
- Darwish, A.G.G.; Samy, M.N.; Sugimoto, S.; Abdel-Salam, H.; Matsunami, K. Effects of Hepatoprotective compounds from the leaves of Lumnitzera racemosa on acetaminophen-induced liver damage in vitro. Chem. Pharm. Bull. 2016, 64, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Paul, T.; Ramasubbu, S. The antioxidant, anticancer and anticoagulant activities of Acanthus ilicifolius L. roots and Lumnitzera racemosa Willd. leaves, from southeast coast of India. J. Appl. Pharm. Sci. 2017, 7, 081–087. [Google Scholar] [CrossRef]
- D’Souza, L.; Wahidulla, S.; Devi, P. Antibacterial phenolics from the mangrove Lumnitzera racemosa. Indian J. Mar. Sci. 2010, 39, 294–298. [Google Scholar]
- Abeysinghe, P.D. Antibacterial activity of aqueous and ethanol extracts of mangrove species collected from Southern Sri Lanka. Asian J. Pharm. Biol. Res. 2012, 2, 79–83. [Google Scholar]
- Yu, S.-Y.; Wang, S.-W.; Hwang, T.-L.; Wei, B.-L.; Su, C.-J.; Chang, F.-R.; Cheng, Y.-B. Components from the leaves and twigs of mangrove Lumnitzera racemosa with anti-angiogenic and anti-inflammatory effects. Mar. Drugs 2018, 16, 404. [Google Scholar] [CrossRef]
- Eswaraiah, G.; Peele, K.A.; Krupanidhi, S.; Indira, M.; Kumar, R.B.; Venkateswarulu, T.C. GC–MS analysis for compound identification in leaf extract of Lumnitzera racemosa and evaluation of its in vitro anticancer effect against MCF7 and HeLa cell lines. J. King Saud Univ. Sci. 2020, 32, 780–783. [Google Scholar] [CrossRef]
- Gosh, M.S.; Gouri, P. Chemical investigation of some mangrove species: Part VIII. Lumnitzera racemosa. J. Indian Chem. Soc. 1980, 57, 568–570. [Google Scholar]
- Saad, S.; Taher, M.; Susanti, D.; Qaralleh, H.; Rahim, N.A.B.A. Antimicrobial activity of mangrove plant (Lumnitzera littorea). Asian Pac. J. Trop. Biomed. 2011, 4, 523–525. [Google Scholar] [CrossRef]
- Le Nguyen, T.T.; Pham, T.T.; Hansen, P.E.; Nguyen, P.K.P. In vitro α-glucosidase inhibitory activity of compounds isolated from mangrove Lumnitzera littorea leaves. Sci. Technol. Dev. J. 2019, 22. [Google Scholar] [CrossRef]
- Wongsomboon, P.; Maneerat, W.; Pyne, S.G.; Vittaya, L.; Limtharakul, T.R. 12-Hydroxycorniculatolide a from the mangrove tree, Lumnitzera littorea. Nat. Prod. Commun. 2018, 13, 1327–1328. [Google Scholar] [CrossRef]
- Debbab, A.; Aly, A.H.; Proksch, P. Mangrove derived fungal endophytes—A chemical and biological perception. Fungal Divers. 2013, 61, 1–27. [Google Scholar] [CrossRef]
- Ancheeva, E.; Daletos, G.; Proksch, P. Lead compounds from mangrove-associated microorganisms. Mar. Drugs 2018, 16, 319. [Google Scholar] [CrossRef]
- Deng, Q.; Li, G.; Sun, M.; Yang, X.; Xu, J. A new antimicrobial sesquiterpene isolated from endophytic fungus Cytospora sp. from the Chinese mangrove plant Ceriops tagal. Nat. Prod. Res. 2018, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Chaeprasert, S.; Piapukiew, J.; Whalley, A.J.; Sihanonth, P. Endophytic fungi from mangrove plant species of Thailand: Their antimicrobial and anticancer potentials. Bot. Mar. 2010, 53, 9. [Google Scholar] [CrossRef]
- Raab, A.; Feldmann, J. Biological sulphur-containing compounds—Analytical challenges. Anal. Chim. Acta 2019, 1079, 20–29. [Google Scholar] [CrossRef]
- Saslis-Lagoudakis, C.H.; Savolainen, V.; Williamson, E.M.; Forest, F.; Wagstaff, S.J.; Baral, S.R.; Watson, M.F.; Pendry, C.A.; Hawkins, J.A. Phylogenies reveal predictive power of traditional medicine in bioprospecting. Proc. Natl. Acad. Sci. USA 2012, 109, 15835–15840. [Google Scholar] [CrossRef]
- Holzmeyer, L.; Hartig, A.-K.; Franke, K.; Brandt, W.; Muellner-Riehl, A.N.; Wessjohann, L.A.; Schnitzler, J. Evaluation of plant sources for antiinfective lead compound discovery by correlating phylogenetic, spatial, and bioactivity data. Proc. Natl. Acad. Sci. USA 2020, 117, 12444–12451. [Google Scholar] [CrossRef]
- Leonhardt, S.D.; Rasmussen, C.; Schmitt, T. Genes versus environment: Geography and phylogenetic relationships shape the chemical profiles of stingless bees on a global scale. Proc. Biol. Sci. 2013, 280, 20130680. [Google Scholar] [CrossRef]
- Schmitt, I.; Barker, F.K. Phylogenetic methods in natural product research. Nat. Prod. Rep. 2009, 26, 1585–1602. [Google Scholar] [CrossRef]
- Adamek, M.; Alanjary, M.; Ziemert, N. Applied evolution: Phylogeny-based approaches in natural products research. Nat. Prod. Rep. 2019, 36, 1295–1312. [Google Scholar] [CrossRef]
- Mawalagedera, S.M.U.P.; Callahan, D.L.; Gaskett, A.C.; Rønsted, N.; Symonds, M.R.E. Combining evolutionary enference and metabolomics to identify plants with medicinal potential. Front. Ecol. Evol. 2019, 7, 471. [Google Scholar] [CrossRef]
- Abdelmohsen, U.R.; Pimentel-Elardo, S.M.; Hanora, A.; Radwan, M.; Abou-El-Ela, S.H.; Ahmed, S.; Hentschel, U. Isolation, phylogenetic analysis and anti-infective activity screening of marine sponge-associated actinomycetes. Mar. Drugs 2010, 8, 399–412. [Google Scholar] [CrossRef]
- Prasad, M.A.; Zolnik, C.P.; Molina, J. Leveraging phytochemicals: The plant phylogeny predicts sources of novel antibacterial compounds. Future Sci. OA 2019, 5, FSO407. [Google Scholar] [CrossRef]
- Grudinski, M.; Pannell, C.M.; Chase, M.W.; Ahmad, J.A.; Muellner-Riehl, A.N. An evaluation of taxonomic concepts of the widespread plant genus Aglaia and its allies across Wallace’s Line (tribe Aglaieae, Meliaceae). Mol. Phylogenet. Evol. 2014, 73, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Dos Santos, C.H.C.; de Carvalho, M.G.; Franke, K.; Wessjohann, L. Dammarane-type triterpenoids from the stem of Ziziphus glaziovii Warm. (Rhamnaceae). Phytochemistry 2019, 162, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, H.; Reider, K.; Franke, K.; Wessjohann, L.A.; Keiser, J.; Dagne, E.; Arnold, N. Characterization of constituents and anthelmintic properties of Hagenia abyssinica. Sci. Pharm. 2012, 80, 433–446. [Google Scholar] [CrossRef]
- Singh, A.; Bajpai, V.; Kumar, K.; Sharma, K.; Kumar, B. Profiling of gallic and ellagic acid derivatives in different plant parts of Terminalia Arjuna by HPLC-ESI-QTOF-MS/MS. Nat. Prod. Comm. 2016, 11, 239–244. [Google Scholar] [CrossRef]
- Atta-Ur-Rahman; Ngounou, F.N.; Choudhary, M.I.; Malik, S.; Makhmoor, T.; Nur-E-Alam, M.; Zareen, S.; Lontsi, D.; Ayafor, J.F.; Sondengam, B.L. New antioxidant and antimicrobial ellagic acid derivatives from Pteleopsis hylodendron. Planta Med. 2001, 67, 335–339. [Google Scholar] [CrossRef]
- Bai, M.; Wu, L.-J.; Cai, Y.; Wu, S.-Y.; Song, X.-P.; Chen, G.-Y.; Zheng, C.-J.; Han, C.-R. One new lignan derivative from the Combretum alfredii Hance. Nat. Prod. Res. 2017, 31, 1022–1027. [Google Scholar] [CrossRef] [PubMed]
- Bathe, U.; Frolov, A.; Porzel, A.; Tissier, A. CYP76 oxidation network of abietane diterpenes in Lamiaceae reconstituted in yeast. J. Agric. Food Chem. 2019, 67, 13437–13450. [Google Scholar] [CrossRef]
- Song, R.; Lin, H.; Zhang, Z.; Li, Z.; Xu, L.; Dong, H.; Tian, Y. Profiling the metabolic differences of anthraquinone derivatives using liquid chromatography/tandem mass spectrometry with data-dependent acquisition. Rapid Commun. Mass Spectrom. 2009, 23, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Frolov, A.; Henning, A.; Böttcher, C.; Tissier, A.; Strack, D. An UPLC-MS/MS method for the simultaneous identification and quantitation of cell wall phenolics in Brassica napus seeds. J. Agric. Food Chem. 2013, 61, 1219–1227. [Google Scholar] [CrossRef]
- Geng, C.-A.; Chen, H.; Chen, X.-L.; Zhang, X.-M.; Lei, L.-G.; Chen, J.-J. Rapid characterization of chemical constituents in Saniculiphyllum guangxiense by ultra fast liquid chromatography with diode array detection and electrospray ionization tandem mass spectrometry. Int. J. Mass Spectrom. 2014, 361, 9–22. [Google Scholar] [CrossRef]
- Correia-da-Silva, M.; Sousa, E.; Pinto, M.M.M. Emerging sulfated flavonoids and other polyphenols as drugs: Nature as an inspiration. Med. Res. Rev. 2014, 34, 223–279. [Google Scholar] [CrossRef]
- Subhashini, P.; Dilipan, E.; Thangaradjou, T.; Papenbrock, J. Bioactive natural products from marine angiosperms: Abundance and functions. Nat. Prod. Bioprospect. 2013, 3, 129–136. [Google Scholar] [CrossRef]
- Kurth, C.; Welling, M.; Pohnert, G. Sulfated phenolic acids from Dasycladales siphonous green algae. Phytochemistry 2015, 117, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Mullen, W.; Borges, G.; Lean, M.E.J.; Roberts, S.A.; Crozier, A. Identification of metabolites in human plasma and urine after consumption of a polyphenol-rich juice drink. J. Agric. Food Chem. 2010, 58, 2586–2595. [Google Scholar] [CrossRef]
- Owczarek, A.; Różalski, M.; Krajewska, U.; Olszewska, M. Rare ellagic acid sulphate derivatives from the rhizome of Geum rivale L.—Structure, cytotoxicity, and validated HPLC-PDA Assay. Appl. Sci. 2017, 7, 400. [Google Scholar] [CrossRef]
- Khac, D.D.; Tran-Van, S.; Campos, A.M.; Jean-Yves Lallemand, J.Y.; Fetizon, M. Ellagic compounds from Diplopanax stachyathus. Phytochemistry 1990, 29, 251–256. [Google Scholar] [CrossRef]
- Shikishima, Y.; Takaishi, Y.; Honda, G.; Ito, M. Phenylbutanoids and stilbene derivatives of Rheum maximowiczii. Phytochemistry 2001, 56, 377–381. [Google Scholar] [CrossRef]
- Pabst, A.; Barron, D.; Adda, J.; Schreier, P. Phenylbutan-2-one β-d-glucosides from raspberry fruit. Phytochemistry 1990, 29, 3853–3858. [Google Scholar] [CrossRef]
- Zhao, D.-K.; Shi, Y.-N.; Petrova, V.; Yue, G.G.L.; Negrin, A.; Wu, S.-B.; D’Armiento, J.M.; Lau, C.B.S.; Kennelly, E.J. Jaboticabin and related polyphenols from Jaboticaba (Myrciaria cauliflora) with anti-inflammatory activity for Chronic Obstructive Pulmonary Disease. J. Agric. Food Chem. 2019, 67, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Terashima, S.; Shimizu, M.; Nakayama, H.; Ishikura, M.; Ueda, Y.; Imai, K.; Suzui, A.; Morita, N. Terashima et al 1990: Studies on aldose reductase Inhibitors from Medicinal plant of "Sinfito" Potentilla candicans, and further synthesis of their related compounds. Chem. Pharm. Bull. 1990, 38, 2733–2736. [Google Scholar] [CrossRef]
- Tomlinson, P.B.; Bunt, J.S.; Primack, R.B.; Duke, N.C. Lumnitzera rosea (Combretaceae)—Its status and floral morphology. J. Arnold Arbor. 1978, 59, 342–351. [Google Scholar]
- Harborne, J.B. Flavonoid sulphates: A new class of sulphur compounds in higher plants. Phytochemistry 1975, 14, 1147–1155. [Google Scholar] [CrossRef]
- Almeida, J.R.; Correia-da-Silva, M.; Sousa, E.; Antunes, J.; Pinto, M.; Vasconcelos, V.; Cunha, I. Antifouling potential of nature-inspired sulfated compounds. Sci. Rep. 2017, 7, 42424. [Google Scholar] [CrossRef]
- Filho, L.C.K.; Picão, B.W.; Silva, M.L.A.; Cunha, W.R.; Pauletti, P.M.; Dias, G.M.; Copp, B.R.; Bertanha, C.S.; Januario, A.H. Bioactive aliphatic sulfates from marine invertebrates. Mar. Drugs 2019, 17, 527. [Google Scholar] [CrossRef]
- Sherman, R.E.; Fahey, T.J.; Howarth, R.W. Soil-plant interactions in a neotropical mangrove forest: Iron, phosphorus and sulfur dynamics. Oecologia 1998, 115, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, E.; Holmer, M.; Bussarawit, N. Benthic metabolism and sulfate reduction in a Southeast Asian mangrove swamp. Mar. Ecol. Prog. Ser. 1991, 73, 93–103. [Google Scholar] [CrossRef]
- Glasenapp, Y.; Korth, I.; Nguyen, X.-V.; Papenbrock, J. Sustainable use of mangroves as sources of valuable medicinal compounds: Species identification, propagation and secondary metabolite composition. S. Afr. J. Bot. 2019, 121, 317–328. [Google Scholar] [CrossRef]
- Husori, D.I.; Sumardi, H.T.; Gemasih, S.; Ningsih, S.R. In vitro anthelmintic activity of Acanthus ilicifolius leaves extracts on Ascardia galli and Pheretima posthuma. J. Appl. Pharm. Sci. 2018, 8, 164–167. [Google Scholar] [CrossRef]
- Sardar, P.K.; Dev, S.; Al Bari, M.A.; Paul, S.; Yeasmin, M.S.; Das, A.K.; Biswas, N.N. Antiallergic, anthelmintic and cytotoxic potentials of dried aerial parts of Acanthus ilicifolius L. Clin. Phytosci. 2018, 4, 894. [Google Scholar] [CrossRef]
- Vattem, D.A.; Shetty, K. Biological functionality of ellagic acid: A review. J. Food Biochem. 2005, 29, 234–266. [Google Scholar] [CrossRef]
- García-Niño, W.R.; Zazueta, C. Ellagic acid: Pharmacological activities and molecular mechanisms involved in liver protection. Pharmacol. Res. 2015, 97, 84–103. [Google Scholar] [CrossRef]
- Zhu, M.; Phillipson, J.D.; Greengrass, P.M.; Bowery, N.E.; Cai, Y. Plant Polyphenols: Biologically active compounds or non-selective binder protein? Phytochemistry 1997, 44, 441–447. [Google Scholar] [CrossRef]
- Michels, K.; Heinke, R.; Schöne, P.; Kuipers, O.P.; Arnold, N.; Wessjohann, L.A. A fluorescence-based bioassay for antibacterials and its application in screening natural product extracts. J. Antibiot. 2015, 68, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-K.; Xu, J.-K.; Zhang, X.-Q.; Yao, X.-S.; Ye, W.-C. Chemical constituents with antibacterial activity from Euphorbia sororia. Nat. Prod. Res. 2008, 22, 353–359. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Code | Collector´s No. | Voucher No. | Collection Date | GenBankAccession (ITS) | Location | Coordinates Lat. (S)/ Long. (E) | Tissue | Extract Amount(mg) | Growth Form |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | LL1 | J.M. 02L-02 | BO1959909 | 20-04-2018 | MT251443 | Ladong Village, Aceh | 5.61/95.49 | Rb | 11 | tree |
| 2 | LL2 | J. M. 03L-7 | BO1959584 | 25-04-2018 | MT251447 | Northern Nias Island | 1.51/ 97.37 | Fr | 6 | tree |
| 3 | LL3 | J.M. 03L-10 | BO1959583 | 26-04-2018 | MT251438 | Southern Nias Island | 0.56/97.78 | Fr | 8 | tree |
| 4 | LL4 | J.M. 04L-3 | BO1959578 | 30-04-2018 | MT251446 | Batam Island | 0.91/104.15 | Fr | 7 | tree |
| 5 | LL5 | J.M. 05L-10 | BO1959420 | 09-05-2018 | MT251437 | Balikpapan, EK | −1.20/116.84 | Fr | 2 | tree |
| 6 | LL6 | J.M. 07L-8 | BO1959417 | 16-05-2018 | MT251442 | Kendari, SS | −4.48/122.13 | Rb | 13 | tree |
| 7 | LL7 | J.M. 08L-10 | BO1959415 | 19-05-2018 | MT251444 | Manado, NS | 1.60/124.85 | Rb | 10 | tree |
| 8 | LL8 | J.M. 09L-15 | BO1959410 | 23-05-2018 | MT251439 | Halmahera Island, Maluku | 1.04/127.50 | Rb | 4 | tree |
| 9 | LL9 | J.M. 13L-12 | BO1959404 | 31-05-2018 | MT251440 | Peling Island, CS | −1.23/123.40 | Fr | 7 | tree |
| 10 | LL10 | J.M. 14L-11 | 05-06-2018 | MT251436 | Luwuk, CS | −0.74/122.96 | Fr | 6 | tree | |
| 11 | LL11 | J.M. 16L-3 | BO1959651 | 29-06-2018 | MT251445 | Banten, West Java | −6.83/105.45 | Rb | 21 | tree |
| 12 | LL12 | J.M. 24L-4 | BO1959653 | 27-07-2018 | MT251441 | Banyuwangi, East Java | −8.59/114.27 | Rb | 14 | tree |
| 13 | LR1_1 | J.M. 01-13 | BO1959913 | 18-04-2018 | MT251462 | Batu Bara, North Sumatra | 3.22/99.57 | Fr | 7 | tree |
| LR1_2 | MT251463 | |||||||||
| LR1_3 | MT251464 | |||||||||
| 14 | LR2 | J.M. 02R-01a | BO1959908 | 21-04-2018 | MT251467 | Ladong Village, Aceh | 5.65/95.45 | Fr | 8 | shrub |
| 15 | LR3_1 | J.M. 02R-02L | BO1959580 | 21-04-2018 | MT251461 | Durung Village, Aceh | 5.61/95.49 | Rb | 22 | shrub |
| LR3_2 | MT251470 | |||||||||
| LR3_3 | MT251469 | |||||||||
| 16 | LR4_1 | J.M. 5R-11 | BO1959421 | 08-05-2018 | MT251473 | Kartanegara, EK | −1.05/117.10 | Fr | 4 | tree |
| LR4_2 | MT251471 | |||||||||
| LR4_3 | MT251468 | |||||||||
| 17 | LR5 | J.M. 06R-3 | BO1959416 | 11-05-2018 | MT251454 | Makassar, South Sulawesi | −5.49/119.32 | Fr | 10 | shrub |
| 18 | LR6 | J.M. 07R-3 | BO1959413 | 16-05-2018 | MT250380 | Kendari, SS | −4.51/122.10 | Fr | 6 | shrub |
| 19 | LR7 | J.M. 10R-2 | BO1959402 | 24-05-2018 | MT251456 | Ternate Island, Maluku | 0.84/127.31 | Fr | 3 | shrub |
| 20 | LR8_1 | J.M. 11R-8 | BO1959776 | 29-05-2018 | MT251472 | Seram Island, Maluku | −3.35/128.36 | Fr | 2 | shrub |
| LR8_2 | MT251466 | |||||||||
| LR8_3 | MT251465 | |||||||||
| 21 | LR9 | J.M. 13R-12 | BO1959649 | 04-06-2018 | MT251458 | Peling Island, Central Sulawesi | −1.23/123.40 | Fr | 9 | shrub |
| 22 | LR10 | J.M. 16R-1 | BO1959655 | 29-06-2018 | MT251457 | Banten, West Java | −6.83/105.45 | Rb | 10 | tree |
| 23 | LR11 | J.M. 18R-14 | BO1959641 | 05-07-2018 | MT251451 | East Sumba, ENT | −9.67/120.33 | Fr | 14 | shrub |
| 24 | LR12 | J.M. 18R-15 | BO1959646 | 05-07-2018 | MT251450 | East Sumba, ENT | −9.64/120.24 | Fr | 19 | shrub |
| 25 | LR13 | J.M. 17R-13 | BO1959642 | 04-07-2018 | MT251449 | Kupang, ENT | −10.15/123.64 | Fr | 5 | shrub |
| 26 | LR14 | J.M. 19R-15 | BO1959644 | 09-07-2018 | MT251459 | Labuan Bajo, ENT | −8.46/119.88 | Fr | 37 | tree |
| 27 | LR15 | J.M. 20R-3 | BO1959423 | 11-07-2018 | MT251453 | Komodo Island, ENT | −8.54/119.55 | Rb | 32 | tree |
| 28 | LR16 | J.M. 21R-9 | BO1959591 | 13-07-2018 | MT251455 | Padar Island, ENT | −8.64/119.58 | Rb | 19 | tree |
| 29 | LR17 | J.M. 22R-15 | BO1959588 | 13-07-2018 | MT251448 | Rinca Island, ENT | −8.65/119.72 | Fr | 21 | tree |
| 30 | LR18 | J.M. 23R-3 | BO1959586 | 20-07-2018 | MT251460 | Bali Island | −8.73/115.20 | Fr | 9 | tree |
| 31 | LR19 | J.M. 24R-15 | BO1959640 | 27-07-2018 | MT251452 | Banyuwangi, East Java | −8.59/114.27 | Fr | 17 | tree |
| No | tR (min) | m/z [M−H]− Observed | m/z [M−H]− Calculated | Elemental Composition | Fragmentation Patterns | RDB | Error (ppm) | Assignment |
|---|---|---|---|---|---|---|---|---|
| 1 | 3.5 | 305.0702 | 305.0700 | C12H18SO7 | 96.9596 (54) | 4 | 0.7 | unknown |
| 2 | 3.8 | 245.0488 | 245.0489 | C10H14SO5 | 79.9566 (22), 96.9592 (53), 130.9649 (9), 165.0506 (100), 168.9878 (77), 201.8320 (8), 219.8438 (8), 243.0313 (7), 245.0478 (53) | 4 | −0.4 | 4-(4-hydroxyphenyl)-2-butanol 2-sulfate |
| 3 | 3.9 | 273.0436 | 273.0438 | C11H14SO6 | 193.0867 (100), 258.0201 (4), 273.0434 (6) c | 5 | −0.7 | 4-(4-sulfoxy-3-methoxy phenyl)-2-butanone (zingeron sulfate) |
| 4 | 3.9 | 275.0591 | 275.0595 | C11H16SO6 | 79.9567 (30), 121.0274 (16), 135.0441 (6), 178.0625 (15), 180.0786 (100), 193.0859 (35), 195.1019 (63), 273.0418 (19), 275.0583 (47).180.0791 (6), 195.1025 (100), 260.0357 (5), 275.0592 (6) c | 4 | −1.5 | unknown(e.g. zingerol sulfate) |
| 5 | 4.4 | 499.1267 | 499.1280 | C22H28SO11 | 96.9594 (5), 499.1267 | 9 | −2.6 | unknown |
| 6 a | 4.4 | 300.9989 | 300.9990 | C14H6O8 | 173.0237 (4), 201.0188 (5), 229.0121 (5), 283.9959 (6), 299.9890 (6), 300.9992 (100) | 12 | −0.3 | ellagic acid |
| 7b | 5.3 | 394.9707 | 394.9715 | C15H8SO11 | 299.9906 (41), 315.0141 (100), 394.9662 (4) | 12 | −2.0 | methylellagic acid sulfate |
| 8 | 5.5 | 487.0179 | 487.0188 | C18H16SO14 | 300.9981 (15), 316.0218 (17), 331.0445 (37), 375.0351 (100) | 11 | −1.8 | unknown ellagic acid derivative |
| 9 | 5.7 | 408.9898 | 408.9871 | C16H10SO11 | 298.9820 (11), 314.0063 (40), 329.0256 (100) | 12 | −6.6 | dimethylellagic acid sulfate, isomer I |
| 10 b | 6.0 | 551.1027 | 551.1026 | C24H24O15 | 312.9971 (4), 328.0211 (14), 343.0452 (100), 491.0806 (2) | 13 | 0.2 | unknown trimethyl ellagic acid derivative |
| 11 | 6.1 | 369.1221 | 369.1225 | C14H26SO9 | 96.9593 (17), 177.0397 (18), 256.9953 (14), 369.1211 (100) | 2 | −1.1 | unknown |
| 12 | 6.3 | 449.2027 | 449.2028 | C20H34O11 | 81.0338 (8), 83.0500 (57), 127.0395 (23), 233.1026 (100), 343.1388 (9), 361.1494 (6), 449.2016 (64) | 4 | −0.2 | unknown |
| 13 | 6.4 | 329.0301 | 329.0303 | C16H10O8 | 242.9944 (5), 270.9875 (35), 298.9824 (60), 314.0052 (100), 329.0290 (29) | 12 | −0.6 | 3,4-O-dimethylellagic acid |
| 14 | 6.5 | 408.9867 | 408.9871 | C16H10SO11 | 298.9813 (6), 314.0049 (15), 329.0292 (100) | 12 | −1.0 | dimethylellagic acid sulfate, isomer II |
| 15 | 6.9 | 423.0035 | 423.0028 | C17H12SO11 | 297.9752 (5), 312.9987 (42), 328.0223 (100), 343.0480 (100), 423.0026 (5) | 12 | 1.7 | 3,3’,4’-trimethyl ellagic acid 4-sulfate |
| 16 | 7.7 | 343.0455 | 343.0459 | C17H12O8 | 269.9798 (9), 285.0031 (6), 297.9744 (28), 312.9981 (69), 328.0217 (100), 343.0443 (18) | 12 | −1.2 | 3,3’,4’-trimethyl ellagic acid |
| 17 a | 9.8 | 487.3425 | 487.3429 | C30H48O5 | 379.3010 (12), 391.3011 (11), 393.3163 (8), 409.3113 (100), 421.3114 (26), 441.3372 (8) | 7 | −0.8 | unknown triterpene acid |
| 18 | 12.2 | 265.1476 | 265.1479 | C12H26SO4 | 79.9562 (12), 96.9596 (100), 98.9556 (9),134.8930 (5), 166.8646 (4), 185.8829 (5), 201.8339 (6), 203.8311 (4) 265.1468 (95) | 0 | −1.1 | unknown aliphatic sulfate |
| 19 | 13.4 | 309.1733 | 309.1741 | C14H30SO5 | 96.9604 (45), 122.9761 (5), 309.1744 (100) c | 0 | −2.6 | unknown aliphatic sulfate |
| 20 | 13.4 | 311.1685 | 311.1686 | C17H28SO3 | 183.0113 (28), 311.1677 (100) | 4 | −0.3 | unknown |
| 21 | 14.6 | 325.1833 | 325.1843 | C18H30SO3 | 183.0111 (23), 325.1833 (100) | 4 | −3.1 | unknown |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manurung, J.; Kappen, J.; Schnitzler, J.; Frolov, A.; Wessjohann, L.A.; Agusta, A.; Muellner-Riehl, A.N.; Franke, K. Analysis of Unusual Sulfated Constituents and Anti-infective Properties of Two Indonesian Mangroves, Lumnitzera littorea and Lumnitzera racemosa (Combretaceae). Separations 2021, 8, 82. https://doi.org/10.3390/separations8060082
Manurung J, Kappen J, Schnitzler J, Frolov A, Wessjohann LA, Agusta A, Muellner-Riehl AN, Franke K. Analysis of Unusual Sulfated Constituents and Anti-infective Properties of Two Indonesian Mangroves, Lumnitzera littorea and Lumnitzera racemosa (Combretaceae). Separations. 2021; 8(6):82. https://doi.org/10.3390/separations8060082
Chicago/Turabian StyleManurung, Jeprianto, Jonas Kappen, Jan Schnitzler, Andrej Frolov, Ludger A. Wessjohann, Andria Agusta, Alexandra N. Muellner-Riehl, and Katrin Franke. 2021. "Analysis of Unusual Sulfated Constituents and Anti-infective Properties of Two Indonesian Mangroves, Lumnitzera littorea and Lumnitzera racemosa (Combretaceae)" Separations 8, no. 6: 82. https://doi.org/10.3390/separations8060082
APA StyleManurung, J., Kappen, J., Schnitzler, J., Frolov, A., Wessjohann, L. A., Agusta, A., Muellner-Riehl, A. N., & Franke, K. (2021). Analysis of Unusual Sulfated Constituents and Anti-infective Properties of Two Indonesian Mangroves, Lumnitzera littorea and Lumnitzera racemosa (Combretaceae). Separations, 8(6), 82. https://doi.org/10.3390/separations8060082