Abstract
This is the first study aiming to enrich momilactones A (MA) and B (MB) and phenolic compounds in germinated brown rice (GBR) and non-GBR var. Koshihikari and Milky Queen through the cooking process. Extraction methods for these compounds were optimized by applying various conditions, including solvents (80% methanol and 80% ethanol), heat (80 °C), and sonication (2 h). Momilactone and phenolic quantities were determined by ultra-performance liquid chromatography–electrospray ionization mass spectrometry (UPLC–ESI-MS) and high-performance liquid chromatography (HPLC), respectively. Accordingly, cooked Koshihikari GBR extract using 80% methanol and sonication (GKB4) revealed the highest amounts of tricin, caffeic, ρ-hydroxybenzoic, ρ-coumaric, ferulic, salicylic, and cinnamic acids (1.71, 1.01, 0.62, 0.45, 0.94, 2.50, and 0.37 mg/g DW, respectively), consistent with the strongest antiradical activities in DPPH and ABTS assays (IC50 = 1.47 and 1.70 mg/mL, respectively). Non-cooked GBR Koshihikari extract using 80% ethanol and sonication (GKB9) exhibited the highest MA and MB contents (147.73 and 118.8 μg/g DW, respectively). Notably, GKB9 showed potent inhibition of α-amylase and α-glucosidase (IC50 = 0.48 and 0.15 mg/mL, respectively), compared with the anti-diabetic drug acarbose (IC50 = 0.26 and 2.48 mg/mL, respectively). The findings hold significant implications for developing phenolic- and momilactone-enriched brown rice with health-beneficial properties.
1. Introduction
Nowadays, consumers are increasingly drawn to pigmented rice varieties for their richness in bioactive constituents, such as antioxidants and anti-inflammatory elements [1]. Among them, brown rice (BR) not only serves as a nutritional powerhouse but also boasts a remarkable array of bioactive compounds that contribute to human health benefits [2,3,4]. BR contains a high level of phenolics and flavonoids, showcasing potential properties that combat oxidative stress and inflammation in the body [5,6]. Studies have also suggested that these compounds may play a role in reducing the risk of chronic diseases and promoting overall wellness [7]. These bioactive compounds are primarily found in the outer layers of the grain, which are retained in brown rice, unlike the processing that removes them in white rice [8]. Hence, the adoption of BR as a nutritious and versatile food option has become a recent trend. Nevertheless, due to the dense composition of its outer bran layer, BR typically possesses a firmer texture, making it more difficult to prepare and less readily digestible in contrast to white rice. Consequently, germinated brown rice (GBR) has emerged as a viable alternative, having added benefits while retaining its nutritional worth. The quality of BR is improved via heightened water absorption in the outer kernel, leading to a softer texture. Moreover, enzymatic processes in the germination stage result in the alteration of bioactive compounds due to interactions among carbohydrates and proteins in the endosperm [1,5]. As a result, GBR is known to contain higher levels of bio-functional components such as γ-aminobutyric acid (GABA) [2,6], vitamins, and amino acids [8], while also exhibiting reduced sugar content [7] in comparison with non-GBR. Earlier research has shown that GBR under abiotic stressors and diverse conditions can show enhanced nutritional characteristics and accumulation of bioactive substances and antioxidants. In our previous study, GBR treated with 75 mM salinity for 4 days presented the highest levels of momilactones and phenolic compounds, which were in line with the strongest antioxidant activity [9]. Hence, employing abiotic stressors and varied germination conditions offers a promising approach to promote the value of BR.
In rice, secondary compounds play significant functions concerning both nutritional value and physiological processes, encompassing metabolism, synthesis, and responses to the environment. Of these, tricin, a vital flavonoid, can be detected from different rice organs (e.g., leaves, husks, brans, and grains). Studies have demonstrated tricin’s potential for antioxidant [10], anti-skin aging [11], and anticancer activities [12,13,14]. On the other hand, phenolic acids, including caffeic, ρ-hydroxybenzoic, ρ-coumaric, ferulic, salicylic, and cinnamic acids are more abundant in rice, and are known for their bioactive attributes, comprising antioxidant, anti-inflammatory, and anticancer properties [15]. It is noteworthy that these phenolic compounds are primarily concentrated in the bran layer [4], making them generally more abundant in BR than in white rice [16,17]. Furthermore, the quantities of these phenolic compounds in GBR can be twice as high as those found in BR [4]. In addition, momilactones A (MA) and B (MB) have been acknowledged as valuable diterpene lactones from rice (Oryza lineage) [18], which exhibit various medicinal properties, including antioxidant [11], anticancer [19,20,21], anti-diabetic [10,12], anti-obesity [12], and anti-skin aging [11] potentials.
BR has demonstrated its potential to contain numerous valuable bioactive compounds such as phenolics and momilactones [2,9,22]. We previously found that these valuable compounds are enriched during the germination process of BR under saline conditions over a 4 day period [9]. However, the impact of cooking on phenolic and momilactone contents and their correlation has not been clearly elucidated. Reports have indicated that cooking regulated the levels of phenolics and flavonoids in rice, underscoring the necessity to include the evaluation of phytochemical contents and associated bioactivities in cooked rice [23]. On the other hand, optimal extraction methods for valuable bioactive compounds, especially momilactones from rice, are an essential approach to harness their biological benefits. However, optimized extraction of momilactones has experienced limited development since they were first discovered in rice. In the extraction process, the selection of solvents (e.g., water, methanol, or ethanol) is an important factor in efficiently obtaining target compounds. Ahmad et al. [24] indicated that the contents of MA and MB were greater in extracts using mixture of methanol and water compared to other solvents. In another consideration, sonication, a non-thermal processing technology that uses high-frequency sound waves to break down plant cell walls, provides favorable conditions for the release of phytochemicals. Studies have shown that sonication-assisted extraction can enhance the availability of antioxidants, MA, MB, phenolic acids, and flavonoids in rice [25,26,27]. Minh et al. [28] reported that extracting rice husk at 100 °C increased the yield of both MA and MB. Of these, MB was obtained with a higher content than MA, despite MA typically being more abundant in rice husk than MB.
The above-mentioned foundations motivated us to undertake a study aimed at enriching the levels of phenolics and momilactones in both GBR and non-GBR through a cooking process. The extraction procedure for these compounds was also optimized by applying various conditions, including solvents (80% methanol and 80% ethanol), temperatures (25 °C and 80 °C), and sonication (2 h). The antioxidant and antidiabetic potentials of rice samples enriched with phenolics and momilactones were also investigated.
2. Materials and Methods
2.1. Materials
Brown rice (BR, Oryza sativa) grains from two rice varieties, including Koshihikari and Milky Queen, were obtained from the Japan Agriculture (JA) shop in Hiroshima, Japan. Extraction solvents, comprising methanol and ethanol, were acquired from Junsei Chemical Co., Ltd. (Tokyo, Japan). Standards, including tricin, caffeic, ρ-hydroxybenzoic, ρ-coumaric, ferulic, salicylic, and cinnamic acids, along with sodium acetate (CH3COONa), sodium carbonate (Na2CO3), acetonitrile, aluminum chloride (AlCl3), methanol plus, Folin–Ciocalteu reagent, 2,2 diphenyl-1-picrylhydrazyl (DPPH), potassium persulfate (K2S2O8), 2,2′-azinobis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS), and sodium hypochlorite (NaClO) were obtained from Kanto Chemical Co., Inc. (Tokyo, Japan). Dimethyl sulfoxide (DMSO) was from Sigma-Aldrich (St. Louis, MO, USA). Trifluoroacetic acid, formic acid, and distilled water were purchased from Fisher Chemical (Hampton, VA, USA) and EMD Millipore Corporation (Billerica, MA, USA), and Nacalai Tesque (Kyoto, Japan), respectively.
2.2. Germination Process
Rice germination was performed according to the procedure outlined by Hasan et al. [9]. Initially, BR grains were sterilized with 0.1% NaOCl for 30 min before washing with clean tap water. Subsequently, the grains (150 g) in each pot were germinated at 30 °C with a 75 mM NaCl salt concentration for 4 days in darkness. Relative humidity (65%) was maintained during the process.
2.3. Cooking Process
After the germination process, both germinated brown rice (GBR) and non-GBR were washed with distilled water and drained for 5 min. After that, rice uniformly underwent cooking using an electric rice cooker (Tiger IH Rice Cooker, Osaka, Japan) with the same condition for all samples. Ultrapure distilled water was used in the cooking process, which eliminated impurities, ions, and microorganisms applying the Direct-Q® UV Water Purification System (Merck KGaA, Darmstadt, Germany). The samples are described in Table 1.

Table 1.
Description of treatments.
2.4. Extraction
Following the cooking process, both cooked and non-cooked samples were subjected to drying in an oven at 40 °C for 7 days. The dried samples were then finely ground into powder. Subsequently, 10 g of powder was extracted using two different solvents, including 80% methanol and 80% ethanol. The extraction process involved three different sonication techniques: (i) 2 h of sonication at 80 °C; (ii) 2 h of sonication at room temperature (RT); and (iii) 2 h of heat at 80 °C. After that, the obtained liquid phase underwent filtration before being evaporated at 45 °C to obtain crude extracts.
2.5. Identification and Quantification of Momilactones A (MA) and B (MB) by Ultra-Performance Liquid Chromatography–Electrospray Ionization-Mass Spectrometry (UPLC–ESI-MS)
The ultra-performance liquid chromatography–electrospray ionization-mass spectrometry (UPLC–ESI-MS) system comprised a mass spectrometer (LTQ Orbitrap XL, Thermo Fisher Scientific, Waltham, MA, USA) with an electrospray ionization (ESI) source. Methanolic sample (3.0 μL) was injected by an autosampler (Vanquish, Thermo Fisher Scientific, Waltham, MA, USA) into a column (1.7 μm, 50 × 2.1 mm i.d.) (Acquity UPLC® BEH C18, Waters Cooperation, Milford, MA, USA) maintained at 25 °C. A gradient mobile phase consisted of solvents A and B, which were trifluoroacetic acid in water (0.1:99.9, v/v) and trifluoroacetic acid in acetonitrile (0.1:99.9, v/v), respectively. The gradient program and MS analysis followed the procedure described in the report of Anh et al. [29]. Calibration curves for MA and MB were established using various standard concentrations (0.5, 1, 5, and 10 μg/mL). Quantities of MA and MB in each sample were determined by applying the peak areas detected in each sample to the respective standard curves.
2.6. Identification and Quantification of Phenolic and Flavonoid Compounds by High-Performance Liquid Chromatography (HPLC)
High-performance liquid chromatography (HPLC) was employed for the identification and quantification of tricin, caffeic acid, ρ-hydroxybenzoic acid, ρ-coumaric acid, ferulic acid, salicylic acid, and cinnamic acid. The HPLC system included a pump (PU-4180 RHPLC, Jasco, Tokyo, Japan), controller (LC-Net II/ADC, Jasco, Japan), and detector (UV-4075 UV/VIS, Jasco, Tokyo, Japan). A column (130 Å, 5 μm, 2.1 × 100 mm) (XBridge BEH Shield RP18, Waters Cooperation, Milford, MA, USA) was used as the stationary phase. The gradient mobile phases, solvent A (0.1% formic acid in water) and solvent B (acetonitrile), were conducted according to the procedure presented by Anh et al. [29]. The analysis of the compounds, lasting 35 min at room temperature, was operated with peak scanning at 350 nm for tricin and at 280 nm for caffeic acid, ρ-hydroxybenzoic acid, ρ-coumaric acid, ferulic acid, salicylic acid, and cinnamic acid. Quantification of these compounds was based on the corresponding peak areas.
2.7. Antioxidant Activities
The radical scavenging abilities of the samples were determined using 2,2-diphenyl-1-picrylhydrazyl (DPPH) and 2,2′-azino-bis (3-ethylbenzthiazoline-6-sulfonic acid) (ABTS) assays, following the procedures presented by Anh et al. [30]. In the DPPH assay, 80 µL of methanolic sample, 40 µL of DPPH working solution (0.5 mM), and 80 µL of acetate buffer (0.1 mM, pH 5.5) were mixed and underwent a 20 min incubation at 25 °C in darkness. For the ABTS assay, 20 µL of the methanolic sample and 180 µL of ABTS working solution underwent a 30 min incubation at 25 °C in darkness. The radical scavenging activities (%) were determined by assessing the reduced absorbance at 517 and 734 for the DPPH and ABTS assays, respectively, in comparison with the control (methanol).
where Ac is the absorbance of the control, As is the absorbance of the sample, and Ab is the absorbance of blank (without radical) solution.
Radical scavenging activity (%) = (Ac − (As − Ab)/Ac) × 100
2.8. α-Amylase Inhibition Assay
α-Amylase inhibitory activity was evaluated based on the starch–iodine method demonstrated by Quan et al. [12] with slight modifications. The extracts were liquefied in 0.2 M phosphate-buffered saline (pH 6.9). α-Amylase solution (5U) was generated by dissolving α-amylase from porcine pancreas (type VI-B, Sigma-Aldrich, St. Louis, MO, USA) in the buffer. Starch (0.5%) and iodine (0.25 mM) solutions were prepared in deionized distilled water. At first, 20 µL of α-amylase solution was diluted and incubated with 20 µL of sample at 37 °C for 9 min. Thereafter, 30 µL of starch (0.5%) solution was added to the mixture followed by incubation for 7 min at 37 °C. Subsequently, 20 µL of HCl (1 M) and 100 µL of iodine solution were pipetted. The mixture was evaluated at 565 nm using a microplate reader (MR, MultiskanTM Microplate Spectrophotometer, Thermo Fisher Scientific, Osaka, Japan). The inhibition percentage of the sample against α-amylase was calculated by the following formula:
where A is the absorbance of the reaction with the presence of the sample, B is the absorbance of the reaction without enzyme, and C is the absorbance of the reaction with absence of the sample. Acarbose was used as a standard inhibitor.
Inhibition (%) = (A − C)/(B − C) × 100
2.9. α-Glucosidase Inhibition Assay
α-Glucosidase inhibitory activity of the extract sample was evaluated using the method described by Quan et al. [12] with few modifications. At the beginning, 20 µL of sample in 40 µL of 0.1 M potassium phosphate buffer (pH 7) was pre-mixed with 20 µL of 0.5 U α-glucosidase enzyme (from Saccharomyces cerevisiae, Sigma Aldrich, St Louis, MO, USA). A 20 µL aliquot of 5 mM p-nitrophenyl-α-D-glucopyranoside (pNPG) substrate (in the buffer) was added after 5 min of incubation at 25 °C. Subsequently, the prepared mixture was incubated for 10 min at 25 °C. Finally, the reaction was suspended by adding 100 µL of 0.1 M Na2CO3. The resulting mixture was measured at 405 nm by a MR. The inhibition percentage of the sample against α-glucosidase was calculated by the following formula:
where As is the absorbance of the reaction with sample or standard inhibitor (acarbose) and Ac is the absorbance of the reaction with DMSO as a negative control.
Inhibition (%) = (1 − As/Ac) × 100
2.10. Statistical Analysis
All experiments were conducted with three replications. Statistical analyses, including one-way ANOVA, were performed using Minitab software (Minitab 16.2.3, Minitab Inc., State College, PA, USA). Results are expressed as mean ± standard deviation (SD). Additionally, the software was employed to calculate Pearson’s correlation coefficients among tested parameters.
A simple ordinary least-squares (OLS) regression was run using the following equation to examine the impacts of different factors on momilactone and phenolic contents extracted from BR and GBR:
where Y = outcome, a0 = constant, βi = impact (treatment effect), xi = treatment, and ui = error term.
Y = a0 + βixi + ui
3. Results and Discussion
3.1. Quantities of Momilactones A (MA) and B (MB) in Rice Samples
Momilactones A (MA) and B (MB) were soon introduced as allelochemicals derived from rice husk [31]. Correspondingly, preceding studies have mostly aimed at the identification and quantification of MA and MB in strong allelopathic rice plants [32,33]. Recently, they have been reported to have multiple human-health-beneficial properties, including antioxidant, anti-diabetes, anti-obesity, anti-skin-aging, and anti-cancer properties [14,34]. Previous screening of 99 rice varieties indicated that rice varieties with awns and later maturing times contained higher levels of MA and MB [35], but most of the studied cultivars were not commercial rice. Hence, our study focused on two popular commercial rice varieties, Koshihikari and Milky Queen, to enrich their momilactone contents and optimize the extraction of these compounds. Figure 1 presents results regarding the contents of MA and MB in germinated brown rice (GBR) and non-GBR of the varieties Koshihikari and Milky Queen through the cooking process and different extraction techniques.
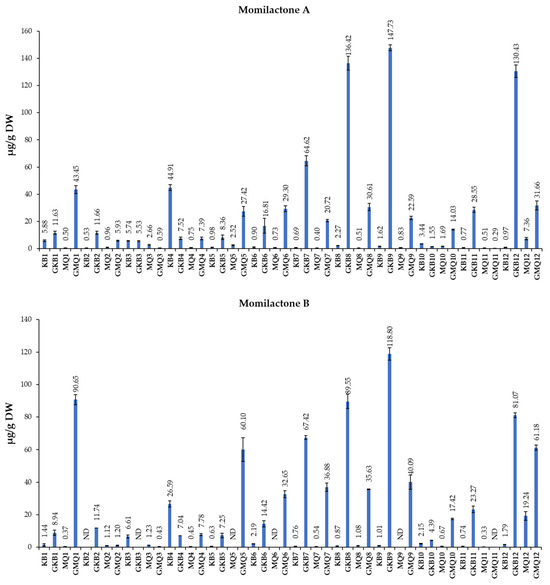
Figure 1.
Quantities of momilactones A (MA) and B (MB) (μg/g DW) in rice samples. Whiskers enclosed in a column express the standard deviation (SD). DW: dry weight; ND: not detected. The description of sample codes is presented in Table 1.
Our study revealed a notably higher concentration of MA (147.73 μg/g DW) and MB (118.8 μg/g DW) in Koshihikari compared to Milky Queen where MA (22.59 μg/g DW) and MB (40.09 μg/g DW) were observed. This could be attributed to the genetic diversity among rice varieties. Previous studies have also indicated variations in momilactone contents in different rice varieties [35,36]. It has been observed that the expression of momilactone biosynthetic genes, including syn-copalyl diphosphate synthase-like (OsCPS4), syn-pimara-7,15-diene synthase-like (OsKSL4), 9-beta-pimara-7,15-diene oxidase-like (CYP99A3), and momilactone A synthase-like (OsMAS and OsMAS2), may vary among rice cultivars, origins, and subtypes Indica and Japonica [36]. The activation of these genes may determine different the MA and MB quantities in studied rice cultivars with different characteristics [36].
The current investigation, for the first time, reveals a significant increase in the quantities of MA and MB in GBR compared to non-GBR, as depicted in Figure 1. This aspect underscores the importance of considering germination as a factor contributing to the enrichment of MA and MB in BR. This phenomenon could be attributed to the elevated expression of genes associated with momilactone biosynthesis, such as OsCPS4, OsKSL4, CYP99A3, OsMAS, and OsMAS2 [29]. Germination may trigger the activation of these genes, leading to a higher production of momilactones in GBR, which needs further elaboration. Upon examining the outcomes presented in Figure 1, it became evident that the cooking process resulted in a slight reduction in MA and MB contents (130.43 and 81.07 μg/g DW, respectively) compared to the non-cooking procedure (147.73 and 118.8 μg/g DW, respectively).
In the research of Minh et al. [28], rice husks subjected to 100 °C, along with a combination of EtOAc and MeOH (v/v), had higher concentrations of both MA and MB compared to untreated rice husks, with MB showing a more significant increase than MA; the most substantial yields of MA (58.76 μg/g DW) and MB (104.43 μg/g DW) were achieved with the EtOAc extract obtained from samples dried at 100 °C for 1 h and subsequently immersed in MeOH at 100% for 1 week [28]. Our research revealed that utilizing 80% ethanol for an 80 °C heat extraction significantly increased the concentrations of MA (130.43 μg/g DW) and MB (81.07 μg/g DW) in cooked Koshihikari GBR compared to other samples. The use of 80% ethanol, coupled with heat at 80 °C, proved to be an effective approach for extracting MA and MB, emphasizing the importance of solvent and temperature conditions in optimizing the extraction process from BR. On the other hand, our research indicated that 80% ethanolic extraction and 2 h of sonication at room temperature are more efficient for quantifying MA (147.73 μg/g DW) and MB (118.80 μg/g DW) in brown rice.
As previously mentioned, the potential roles of MA and MB in preventing chronic diseases and cancer have been being explored. Several potential approaches, including induced gene expression, metabolic engineering techniques, and genetic modifications, have been proposed to enhance the biosynthesis of MA and MB within rice sources to increase their exploitable value [37]. For instance, a study highlighted that N-methyl-N-nitrosourea mutations could lead to increased accumulation of MA and MB in mutated rice lines [38]. In this study, we propose, for the first time, several methods to enrich and extract MA and MB from BR. Among the 48 samples, non-cooked Koshihikari GBR extracted using 80% ethanol and 2 h sonication at room temperature (RT) demonstrated the highest concentrations of MA and MB (147.73 and 118.8 μg/g DW, respectively), and non-cooked Milky Queen BR extracted using 80% ethanol and 2 h sonication at 80 °C demonstrated the lowest concentrations of MA and MB (0.4 and 0.54 μg/g DW, respectively), as detailed in Figure 1.
3.2. Contents of Tricin, Caffeic Acid, ρ-Hydroxybenzoic Acid, ρ-Coumaric Acid, Ferulic Acid, Salicylic Acid, and Cinnamic Acid in Rice Samples
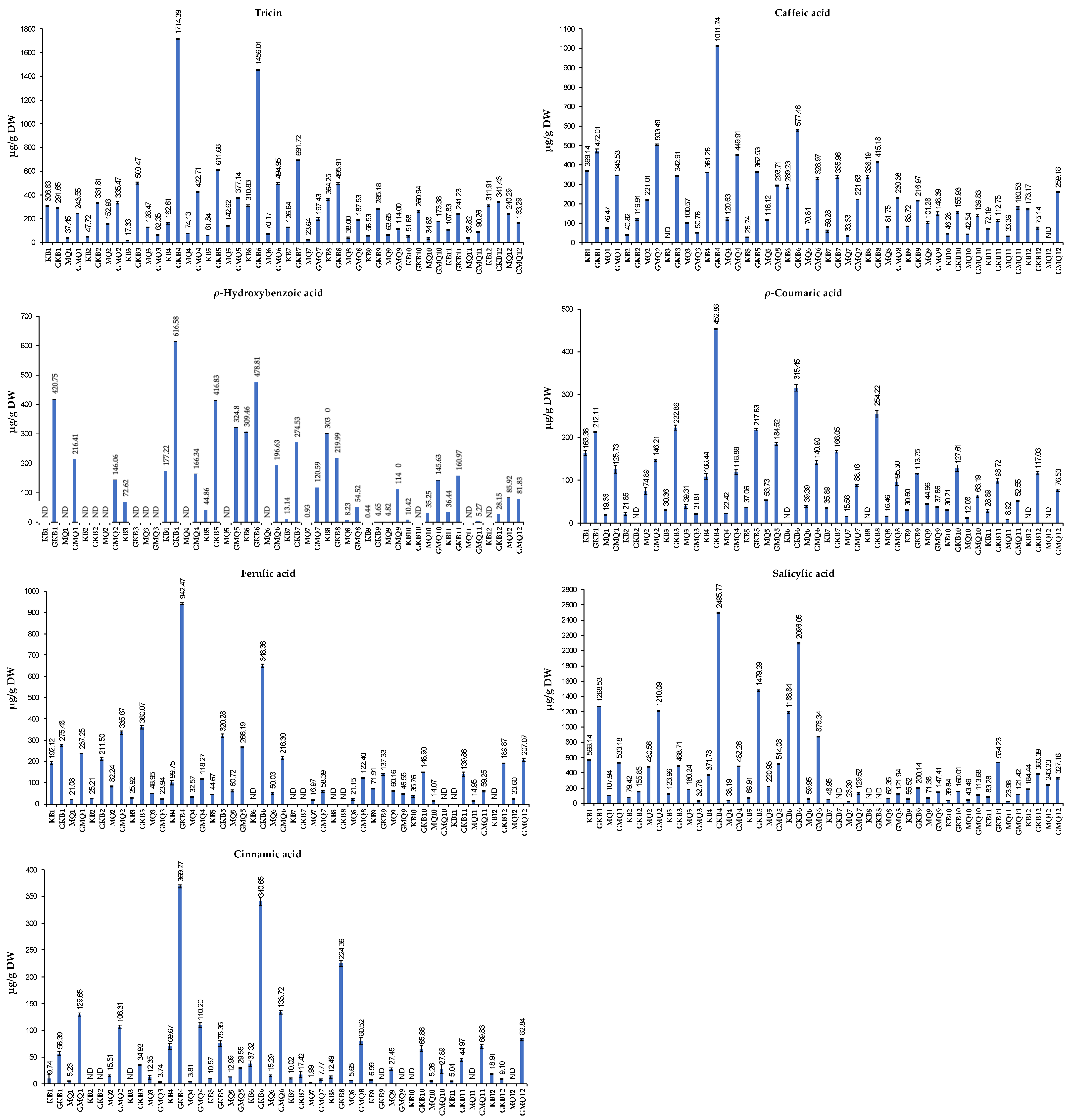
The results in Figure 2 show that cooked Koshihikari GBR extracted using 80% MeOH and sonication at RT (GKB4) exhibited the highest concentrations of tricin, caffeic acid, ρ-hydroxybenzoic acid, ρ-coumaric acid, ferulic acid, salicylic acid, and cinnamic acid (1714.39, 1011.24, 616.58, 452.88, 942.47, 2495.77, and 369.27 μg/g DW, respectively). Conversely, non-cooked Milky Queen BR extracted using 80% EtOH and sonication at 80 °C showed the lowest contents of these phenolic compounds (23.64, 33.33, 0.93, 15.56, 16.97, 23.39, and 1.99 μg/g DW, respectively) (Figure 2). According to Tian et al. [39], the swift rise in free phenolic acid during seed germination is primarily attributed to the activation of endogenous esterase. The elevation of bound phenolic acids in GBR is likely a result of polymerization originating from free phenolics, as suggested by Ti et al. [40]. The increased concentration of free phenolic acid in GBR may be attributed to the protective role of rice, preventing the loss and oxidation of free phenolic acid.
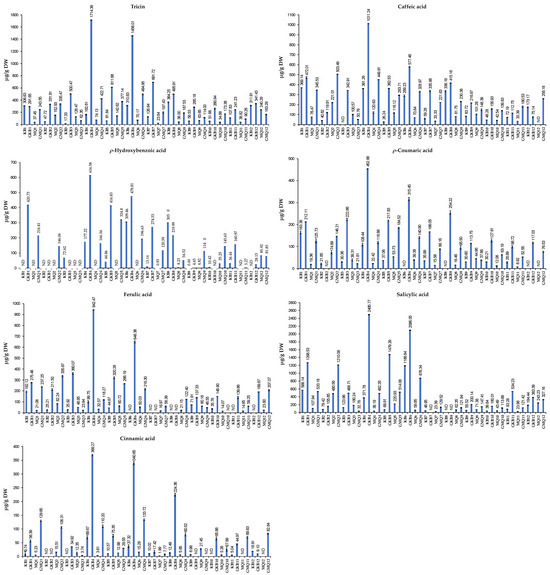
Figure 2.
Quantities of tricin, caffeic acid, ρ-hydroxybenzoic acid, ρ-coumaric acid, ferulic acid, salicylic acid, and cinnamic acid (μg/g DW) in rice samples. Whiskers enclosed in a column express the standard deviation (SD). DW: dry weight; ND: not detected. The description of sample codes is presented in Table 1.
In the previous study, tricin, ρ-coumaric acid, ferulic acid, salicylic acid, and cinnamic acid (107.63, 93.77, 139.03, 46.05, and 596.26 μg/g DW, respectively) exhibited ascending concentrations in GBR [9]. In the current study, all these compounds significantly increased after cooking, with the highest amounts found in sample GKB4 (cooked Koshihikari GBR, extracted using 80% methanol with 2 h of sonication at RT). For extraction, water, methanol, 80% acetone, and 80% ethanol were identified as more effective solvents for obtaining phenolic and flavonoid compounds and antioxidant capacities [41,42]. However, the current study indicated that 80% methanolic extract was relatively more effective in acquiring phenolics and flavonoids. It is possible that the inclusion of other controls in the study, such as germination, the cooking process, and sonication, has contributed to the higher levels of phenolics and flavonoids in BR.
3.3. Average Values of Momilactones A (MA) and B (MB), Phenolic Acids, and Flavonoids in Rice Varieties under Germination Conditions, Cooking Processes, and Extraction Methods
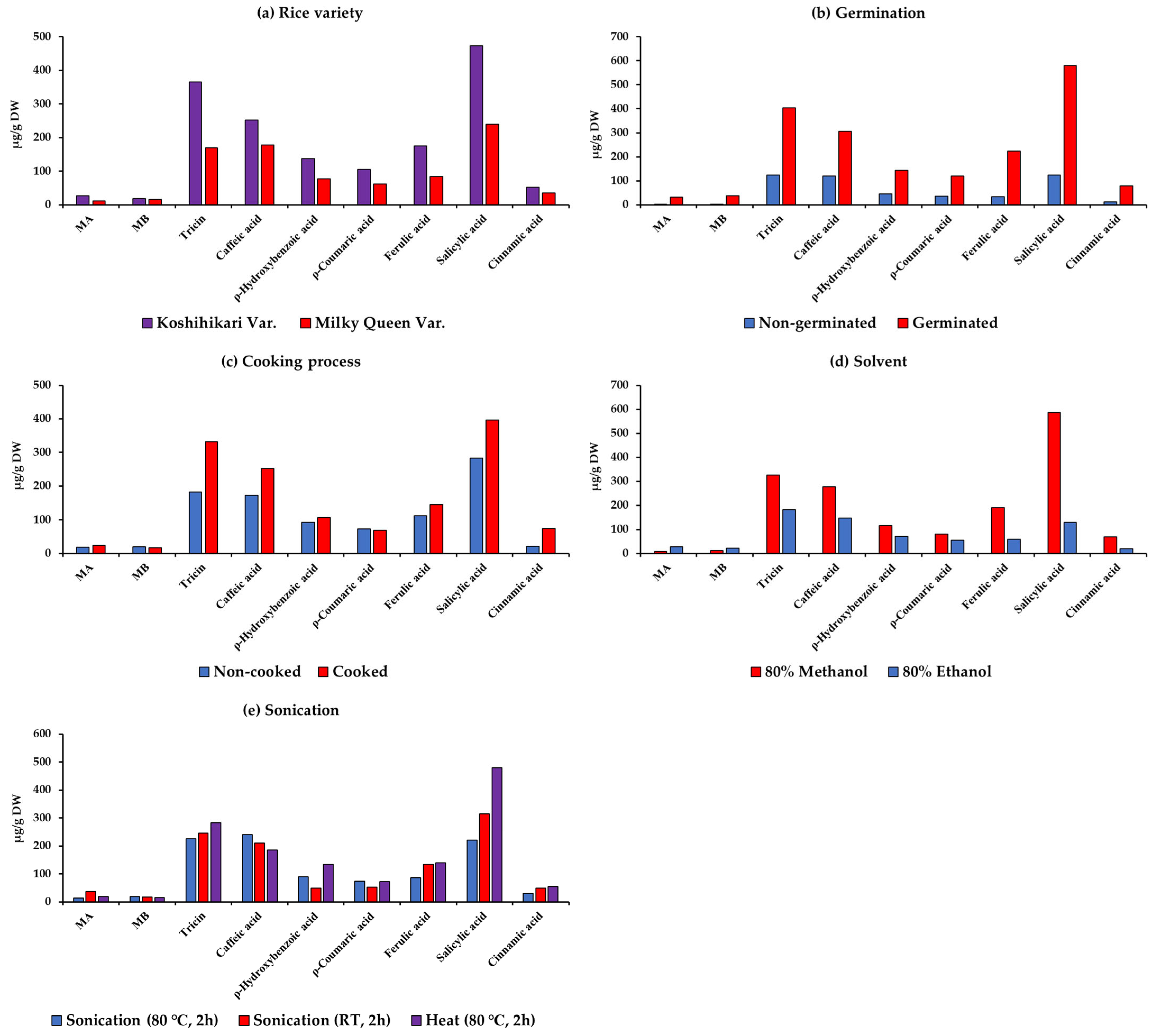
Figure 3 presents the average values of MA, MB, tricin, caffeic acid, ρ-hydroxybenzoic acid, ρ-coumaric acid, ferulic acid, salicylic acid, and cinnamic acid in Koshihikari and Milky Queen under different germination conditions, cooking processes, and extraction procedures.
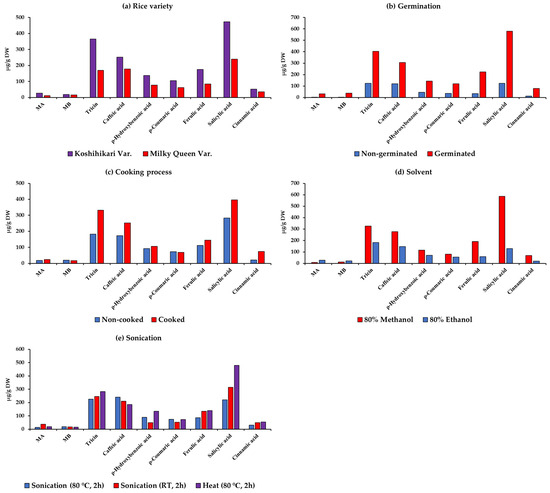
Figure 3.
Average contents of momilactones A (MA) and B (MB), phenolic acids, and flavonoids among rice varieties under different germination conditions, cooking processes, and extraction methods.
The outcomes showed that Koshihikari had higher contents of MA, MB, tricin, caffeic acid, ρ-hydroxybenzoic acid, ρ-coumaric acid, ferulic acid, salicylic acid, and cinnamic acid than Milky Queen (Figure 3a). In Figure 3b, it is evident that GBR contained higher amounts of all compounds compared to non-GBR. Our findings could contribute to enhancing the value of GBR, which can be directly consumed as a component in dishes, harnessing its health-beneficial properties. Although less common than white rice, GBR might have been consumed since ancient times [43]. For example, the total consumption of soaked GBR in Japan is approximately 9 tons annually [43]. GBR is also served in restaurants and frequently featured in health and fashion magazines. Both older and younger generations, particularly those interested in health, highly appreciate GBR [43]. It is utilized in various products, such as rice balls, soups, bread, pastries, rice burgers, etc., often combined with other ingredients. GBR has been incorporated into numerous cuisines worldwide, such as Italian risotto, Spanish paella, Brazilian feijoada, etc. [43].
During the use of rice as a food source, the cooking process induces changes in its physical, chemical, and biological composition, consequently altering its structural characteristics and nutritional contents, as reported by Zhanqian et al. [44]. Beneficial effects of cooking include enhanced digestibility and food detoxification, contributing to safer consumption for humans [45]. Conversely, cooking can modify the levels of bioactive compounds, thereby altering the biological value and nutritional profile of the food [46]. Therefore, the impact of the cooking process on momilactones in rice is a topic of our current interest, which has not been investigated in other studies. After cooking, the availability of MA, tricin, caffeic acid, ρ-hydroxybenzoic acid, ferulic acid, salicylic acid, and cinnamic acid in rice samples significantly increased, while MB and ρ-coumaric acid decreased (Figure 3c).
In the extraction process of bioactive phytocompounds, various solvents (e.g., water, methanol, ethanol, hexane, ethyl acetate, etc.) have been used [47]. Among them, aqueous organic solvents have been demonstrated to be more effective than individual organic solvents. Particularly, plant extraction using aqueous methanol yields the highest phenolic content compared to other solvents [48,49]. On the other hand, the most effective extraction of MA and MB was achieved through 100% methanolic extraction of rice husks treated at 100 °C [28]. In another study, higher enrichment of MA and MB was observed with Soxhlet extraction employing a combination of MeOH and MeOH:water, surpassing other methods [24]. While aqueous methanol proves to be an effective solvent in extracting phenolics and momilactones from rice materials, its toxicity renders it less preferable in applications intended for human consumption of the final product [50]. Compared to methanol, ethanol exhibits almost similar polarity and also is extensively used to extract a wide range of natural substances [51]. Despite also posing potential toxicity risks to humans, ethanol is considered less toxic than methanol and might be preferred in the food and pharmaceutical industries [51]. Notably, we observed that extraction with 80% ethanol yielded the greatest quantities of momilactones, while extraction with 80% methanol revealed the highest amount of phenolic and flavonoid compounds, as depicted in Figure 3d. Our results may contribute to standardizing an extraction method for screening rice samples with dominant quantities of phenolics and momilactones. Additionally, our findings might contribute to developing extraction processes to obtain high levels of target compounds from rice on an industrial scale, aimed at producing functional products with health-beneficial properties. However, further extensive investigations are needed to confirm their possibility and efficacy in industrial implications. On the other hand, strict adherence to safety regulations is paramount when handling ethanol in industrial settings and applications that directly involve consumers.
Sonication plays a crucial role in extracting bioactive compounds from plant samples, enhancing the efficiency of the extraction process, and improving the yield of these compounds. The application of sonication is instrumental in breaking down cell structures and facilitating the release of compounds from the rice matrix [24]. In Figure 3e, 2 h of sonication at 80 °C resulted in the highest amounts of MB and caffeic acid, while 2 h of sonication at RT led to the highest amount of MA. Additionally, 2 h of heat at 80 °C determined a significantly higher quantity of tricin, ρ-hydroxybenzoic acid, ρ-coumaric acid, ferulic acid, salicylic acid, and cinnamic acid compared to 2 h of sonication at RT and 80 °C.
Another key consideration is the impact of the drying process on phenolic and momilactone contents in rice samples. Studies have highlighted variations in the levels of bioactive compounds during the drying process, indicating increases or losses depending on the compound types [52]. Under the drying process, bioactive compounds react with oxygen at high temperatures, leading to their degradation owing to high reducibility. The diverse molecular structures of various bioactive components cause varying susceptibilities to temperature [52]. Specifically, phenolic compounds, being reducing agents, are susceptible to oxidation under the drying process. Moreover, higher drying temperatures escalate the reaction rates in phenolics. Therefore, lower drying temperatures are favored to achieve a higher preservation rate for these compounds [52]. However, concerning momilactones, no studies have explored the impact of the drying process on these compounds; thus, investigating this aspect is highly recommended in subsequent studies. In general, the decomposition of bioactive compounds during convective drying primarily occurs due to prolonged exposure of the sample to high temperatures [52]. In this study, we employed the convective drying method for rice samples at 40 °C for 7 days. However, the actual impact of these conditions on variations in the levels of phenolics and momilactones in the samples has not been evaluated. Thus, the optimization of drying methods for these targeted compounds necessitates thorough exploration in future investigations. Other drying methods, such as microwave freeze drying, infrared, and vacuum drying may also hold potential advantages over convective drying in terms of product quality that should be also considered [52]. Overall, research and development to achieve optimal drying methods with high product quality, low cost, energy efficiency, and environmental friendliness are necessary to advance commercial products based on momilactone- and phenolic-enriched rice grains.
3.4. Ordinary Least-Square Regression (OLS) Estimation for the Effects of Different Treatments on Brown Rice
Table S1 illustrates the average treatment effects of rice variety on momilactones, phenolics, and flavonoids in BR, highlighting a significant boost in Koshihikari at a 5% significance level according to the OLS estimate. Conversely, compared to in Milky Queen, tricin, ρ-hydroxybenzoic acid, and ρ-coumaric acid decreased by −218.4, −3815, −78.38, and −53.59, respectively. Table S2 demonstrates that germination had positive and statistically significant effects on momilactones, phenolics, and flavonoids, with significance observed at a 1% level for all compounds. Germination enhanced the availability of MA, MB, tricin, caffeic acid, ρ-hydroxybenzoic acid, ρ-coumaric acid, ferulic acid, salicylic acid, and cinnamic acid in BR. Table S3 reveals that cooking had a positive and statistically significant impact only on cinnamic acid at a 5% significance level, with an increase of 48.24. Cooking showed no statistically significant impact on the other nine components. In Table S4, the average treatment effects of 80% methanol and 80% ethanol on nine compounds indicate that 80% ethanol significantly elevated MA and MB at a 1% significance level while 80% methanol had positive and statistically significant impacts on tricin, caffeic acid, ρ-hydroxybenzoic acid, ρ-coumaric acid, ferulic acid, salicylic acid, and cinnamic acid at a 5% significance level. Table S5 demonstrated that 2 h of sonication at 80 °C and room temperature had no statistically significant impacts on any tested compounds, but 2 h of heat at 80 °C had positive and statistically significant impacts on tricin, caffeic acid, ρ-hydroxybenzoic acid, ρ-coumaric acid, ferulic acid, salicylic acid, and cinnamic acid.
3.5. Antioxidant Activity by the DPPH and ABTS Radical Scavenging Assay
Following the screening of momilactone and phenolic contents, we selected three samples including GKB4 (cooked Koshihikari GBR extracted by 80% methanol with 2 h of sonication at RT), GKB9 (non-cooked Koshihikari GBR extract by 80% ethanol with 2 h of sonication at RT), and MQ7 (non-cooked Milky Queen non-GBR extracted by 80% ethanol with 2 h of sonication at 80 °C). Of these, GKB4 was highly enriched in phenolic and flavonoid contents, GKB9 exhibited the greatest contents of MA and MB, while MQ7 contained the lowest quantities of momilactones and phenolics. The DPPH radical scavenging activity of the selected samples is presented in Table 2. Accordingly, GKB4 exhibited the strongest inhibition against DPPH cations (IC50 = 1.47 mg/mL), followed by GKB9 and MQ7 (IC50 = 1.56 and 4.98 mg/mL, respectively). In the ABTS assay, MQ7 showed the weakest activity, with IC50 values of 3.49 mg/mL, while GKB4 and GKB9 displayed the strongest activity, with IC50 values of 1.70 and 1.80 mg/mL, respectively. Sample GKB4 showed the strongest antioxidant activity as well as being enriched in phenolics and flavonoid contents. A previous study pointed out that the antioxidant activity of whole rice grain is closely correlated with its free and bound phenolic contents [53]. MQ7 displayed weaker antioxidant effects, correlated with its lower phenolic and flavonoid contents. These findings contribute to the understanding of the relationship between specific compounds and antioxidant potential in rice, emphasizing the importance of phenolics and flavonoids in conferring antioxidant activity to whole rice grains.
Table 2.
Antioxidant, α-amylase, and α-glucosidase inhibitory activity in selected rice samples.
3.6. α-Amylase and α-Glucosidase Inhibition Assay
The inhibition of α-amylase and α-glucosidase was assessed for three selected samples, GKB4 (cooked Koshihikari GBR extract using 80% methanol with 2 h of sonication at RT), GKB9 (non-cooked Koshihikari GBR extract uisng 80% ethanol with 2 h of sonication at RT), and MQ7 (non-cooked Milky Queen non-GBR extract using 80% ethanol with 2 h of sonication at 80 °C), as shown in Table 2. GKB9 exhibited the highest inhibitory activity against α-amylase (IC50 = 0.48 mg/mL) and α-glucosidase (IC50 = 0.15 mg/mL). In contrast, MQ7 displayed the lowest inhibitory activity against α-amylase (IC50 = 4.89 mg/mL) and minor effects on α-glucosidase. In this study, GKB9 inhibited the most potent inhibition against α-amylase and α-glucosidase, which was consistent with it having the highest MA and MB contents. Remarkably, Quan et al. [12] reported that MA and MB, detected and isolated from rice, were effective inhibitors of α-amylase and α-glucosidase, suggesting their potential exploration as novel and potent candidates for antidiabetic therapy.
3.7. Comparison of Extraction Techniques for Momilactones A (MA) and B (MB) from Different Rice Types
Table 3 presents a comparison between previous and current studies, encompassing various parts, varieties, and extraction methods for rice. For example, MA and MB were found in straw with concentrations of 3.8 and 2.01 μg/g DW, respectively [54]. The levels of these compounds were determined in the aerial parts of 30 rice varieties (ranging from 69.9 to 99.3 μg/g DW for MA and 64.4 to 114.1 μg/g DW for MB) [36]. MA and MB were also quantified in various plant parts (e.g., husks and grains) [10,28] (Table 3). In our present study, GBR exhibited elevated levels of MA and MB (147.73 and 118 μg/g DW, respectively). Furthermore, the levels of MA and MB in rice grain (1.56 and 1.61 μg/g DW, respectively) [11] and GBR (18.94 and 41.00 μg/g DW, respectively) [9] in previous findings were significantly lower than those observed in the present study. Our current study demonstrated that 80% ethanol was more effective for the extraction of MA and MB than 80% methanol. In contrast, this study indicates that utilizing 80% ethanolic extraction with sonication at room temperature can result in higher yields of MA and MB in GBR.
Table 3.
Comparison of extraction techniques for MA and MB from different rice samples.
4. Conclusions
This study, for the first time, identified an optimized method for enriching and extracting momilactones A (MA) and B (MB) and phenolic compounds from germinated brown rice (GBR) and non-GBR of the Koshihikari and Milky Queen varieties through the cooking process. Specifically, cooked Koshihikari GBR extracts using 80% methanol and 2 h sonication (GKB4) exhibited the highest contents of phenolic compounds, closely correlating with them having the strongest antioxidant activity. On the other hand, non-cooked Koshihikari GBR extract using 80% ethanol and 2 h sonication (GKB9) showed the greatest quantities of MA and MB, consistent with its ability to inhibit α-amylase and α-glucosidase. Notably, the activity of GKB9 was comparable to the diabetes drug acarbose. In the context of the underappreciation of brown rice (BR), leading to its improper utilization or wastage, our research results hold promise in enhancing the nutritional value of BR and fostering the development of rice-derived products that contribute to improving human health. Therefore, this study aims to encourage BR consumption by emphasizing its intrinsic value and potential benefits. Moreover, these current findings are expected to contribute to achieving the Sustainable Development Goals (SDGs) by promoting the collective welfare of individuals, eradicating hunger and poverty, and ensuring global food security, especially in rice-dependent nations.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/separations11010006/s1, Table S1: OLS estimation for different rice varieties; Table S2: OLS estimation for non-germinated and germinated samples; Table S3: OLS estimation for non-cooked and cooked samples; Table S4: OLS estimation for 80% ethanol and 80% methanol samples; and Table S5: OLS estimation for sonicated samples.
Author Contributions
Conceptualization, T.D.X.; methodology, L.H.A. and M.H.; investigation, M.H. and L.H.A.; data curation, M.H. and L.H.A.; writing—original draft preparation, M.H. and L.H.A.; writing—review and editing, M.H., L.H.A. and T.D.X.; supervision, T.D.X. and L.H.A.; project administration, T.D.X. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The corresponding author can provide data supporting the findings of this study upon request.
Acknowledgments
We express our gratitude to Nguyen Van Quan, Nguyen Phuong Mai, and Nguyen Thi Doan for their valuable support in sample preparation and analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Alves, G.H.; Ferreira, C.D.; Vivian, P.G.; Monks, J.L.F.; Elias, M.C.; Vanier, N.L.; De Oliveira, M. The revisited levels of free and bound phenolics in rice: Effects of the extraction procedure. Food Chem. 2016, 208, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Ravichanthiran, K.; Ma, Z.F.; Zhang, H.; Cao, Y.; Wang, C.W.; Muhammad, S.; Aglago, E.K.; Zhang, Y.; Jin, Y.; Pan, B. Phytochemical profile of brown rice and its nutrigenomic implications. Antioxidants 2018, 7, 71. [Google Scholar] [CrossRef] [PubMed]
- Sukegawa, H.; Kokawa, M.; Kitamura, Y. Establishment of a cultivation method for sprouted brown rice and elucidation of its functional food characteristics. Food Sci. Technol. Res. 2021, 27, 341–349. [Google Scholar] [CrossRef]
- Wu, X.; Guo, T.; Luo, F.; Lin, Q. Brown Rice: A missing nutrient-rich health food. Food Sci. Hum. Wellness. 2023, 12, 1458–1470. [Google Scholar] [CrossRef]
- Gong, E.S.; Luo, S.J.; Li, T.; Liu, C.M.; Zhang, G.W.; Chen, J.; Zeng, Z.C.; Liu, R.H. Phytochemical profiles and antioxidant activity of brown rice varieties. Food Chem. 2017, 227, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, A.; Shabbir, U.; Chen, X.; Chelliah, R.; Elahi, F.; Ham, H.J.; Oh, D.H. Phytochemical profiling and cellular antioxidant efficacy of different rice varieties in colorectal adenocarcinoma cells exposed to oxidative stress. PLoS ONE 2022, 17, e0269403. [Google Scholar] [CrossRef] [PubMed]
- Barber, T.M.; Kabisch, S.; Pfeiffer, A.F.H.; Weickert, M.O. The health benefits of dietary fibre. Nutrients 2020, 12, 3209. [Google Scholar] [CrossRef]
- Verardo, V.; Gómez-Caravaca, A.M.; Marconi, E.; Segura-Carretero, A.; Garrido-Frenich, A.; Fernández-Gutiérrez, A. Determination of lipophilic and hydrophilic bioactive compounds in raw and parboiled rice bran. RSC Adv. 2016, 6, 50786–50796. [Google Scholar] [CrossRef]
- Hasan, M.; Quan, N.V.; Anh, L.H.; Khanh, T.D.; Xuan, T.D. Salinity treatments promote the accumulations of momilactones and phenolic compounds in germinated brown rice. Foods 2023, 12, 2501. [Google Scholar] [CrossRef]
- Quan, N.V.; Xuan, T.D.; Tran, H.D.; Ahmad, A.; Khanh, T.D.; Dat, T.D. Contribution of momilactones A and B to diabetes inhibitory potential of rice bran: Evidence from in vitro assays. Saudi Pharm. J. 2019, 27, 643–649. [Google Scholar] [CrossRef]
- Quan, N.V.; Thien, D.D.; Khanh, T.D.; Tran, H.D.; Xuan, T.D. Momilactones A, B, and tricin in rice grain and by-products are potential skin aging inhibitors. Foods 2019, 8, 602. [Google Scholar] [CrossRef]
- Quan, N.V.; Tran, H.D.; Xuan, T.D.; Ahmad, A.; Dat, T.D.; Khanh, T.D.; Teschke, R. Momilactones A and B are α-amylase and α-glucosidase inhibitors. Molecules 2019, 24, 482. [Google Scholar] [CrossRef] [PubMed]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compos. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Wongsa, P. Phenolic compounds and potential health benefits of pigmented rice. J. Rice Res. Dev. 2020, 4, 19–21. [Google Scholar]
- Delbaere, K.; Close, J.C.T.; Brodaty, H.; Sachdev, P.; Lord, S.R. Determinants of disparities between perceived and physiological risk of falling among elderly people: Cohort study. BMJ 2010, 341, 4165. [Google Scholar] [CrossRef]
- Do Nascimento, L.Á.; Abhilasha, A.; Singh, J.; Elias, M.C.; Colussi, R. Rice germination and its impact on technological and nutritional properties: A review. Rice Sci. 2022, 29, 201–215. [Google Scholar] [CrossRef]
- Gujral, H.S.; Sharma, P.; Bajaj, R.; Solah, V. Effects of incorporating germinated brown rice on the antioxidant properties of wheat flour chapatti. Food Sci. Technol. Int. 2012, 18, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Anh, L.H.; Khanh, T.D.; Xuan, T.D. Biological roles of momilactones: Achievements, challenges, and promising approaches to exploit their beneficial properties. Front. Nat. Prod. 2023, 2, 1245869. [Google Scholar] [CrossRef]
- Park, C.; Jeong, N.Y.; Kim, G.Y.; Han, M.H.; Chung, I.M.; Kim, W.J.; Yoo, Y.H.; Choi, Y.H. Momilactone B induces apoptosis and G1 arrest of the cell cycle in human monocytic leukemia U937 cells through downregulation of PRB phosphorylation and induction of the cyclin-dependent kinase inhibitor P21Waf1/Cip1. Oncol. Rep. 2014, 31, 1653–1660. [Google Scholar] [CrossRef]
- Lee, S.C.; Chung, I.-M.; Jin, Y.J.; Song, Y.S.; Seo, S.Y.; Park, B.S.; Cho, K.H.; Yoo, K.S.; Kim, T.H.; Yee, S.B. Momilactone B, an allelochemical of rice hulls, induces apoptosis on human lymphoma cells (Jurkat) in a micromolar concentration. Nutr. Cancer 2008, 60, 542–551. [Google Scholar] [CrossRef]
- Kim, S.J.; Park, H.R.; Park, E.; Lee, S.C. Cytotoxic and antitumor activity of momilactone B from rice hulls. J. Agric. Food Chem. 2007, 55, 1702–1706. [Google Scholar] [CrossRef] [PubMed]
- Cho, D.H.; Lim, S.T. Germinated brown rice and its bio-functional compounds. Food Chem. 2016, 196, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Ti, H.; Zhang, R.; Li, Q.; Wei, Z.; Zhang, M. Effects of cooking and in vitro digestion of rice on phenolic profiles and antioxidant activity. Food Res. Int. 2015, 76, 813–820. [Google Scholar] [CrossRef]
- Ahmad, A.; Xuan, T.D.; Minh, T.N.; Siddiqui, N.A.; Quan, N.V. Comparative extraction and simple isolation improvement techniques of active constituents’ momilactone A and B from rice husks of Oryza Sativa by HPLC analysis and column chromatography. Saudi Pharm. J. 2019, 27, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Zubair, M.; Anwar, F.; Zafar, M.N.; Nazar, M.F. Effect of ultrasonic extraction regimes on phenolics and antioxidant attributes of rice (Oryza sativa L.) cultivars. IJCCE 2018, 37, 109–119. [Google Scholar]
- Bonto, A.P.; Tiozon, R.N.; Sreenivasulu, N.; Camacho, D.H. Impact of ultrasonic treatment on rice starch and grain functional properties: A review. Ultrason. Sonochem. 2021, 71, 105383. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Pan, Z.; Yue, T.; Atungulu, G.G.; Berrios, J. Effect of ultrasonic treatment of brown rice at different temperatures on cooking properties and quality. Cereal Chem. 2010, 87, 403–408. [Google Scholar] [CrossRef]
- Minh, T.N.; Xuan, T.D.; Ahmad, A.; Elzaawely, A.A.; Teschke, R.; Van, T.M. Momilactones A and B: Optimization of yields from isolation and purification. Separations 2018, 5, 28. [Google Scholar] [CrossRef]
- Anh, L.H.; Quan, N.V.; Quang, V.L.; Takami, A.; Khanh, T.D.; Xuan, T.D. Rice momilactones and phenolics: Expression of relevant biosynthetic genes in response to UV and chilling stresses. Agronomy 2022, 12, 1731. [Google Scholar]
- Anh, L.H.; Quan, N.V.; Lam, V.Q.; Iuchi, Y.; Takami, A.; Teschke, R.; Xuan, T.D. Antioxidant, anti-tyrosinase, anti-α-amylase, and cytotoxic potentials of the invasive weed Andropogon virginicus. Plants 2021, 10, 69. [Google Scholar] [CrossRef]
- Kato, T.; Kabuto, C.; Sasaki, N.; Tsunagawa, M.; Aizawa, H.; Fujita, K. Momilactones, growth inhibitors from rice, Oryza sativa L. Tetrahedron. Lett. 1973, 39, 3861–3864. [Google Scholar] [CrossRef]
- Kong, C.; Liang, W.; Xu, X.; Hu, F.; Wang, P.; Jiang, Y. Release and activity of allelochemicals from allelopathic rice seedlings. J. Agric. Food Chem. 2004, 52, 2861–2865. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.H.; Li, H.B.; Hu, F.; Xu, X.H.; Wang, P. Allelochemicals released by rice roots and residues in soil. Plant Soil. 2006, 288, 47–56. [Google Scholar] [CrossRef]
- Goufo, P.; Trindade, H. Rice Antioxidants: Phenolic acids, flavonoids, anthocyanins, proanthocyanidins, tocopherols, tocotrienols, γ-oryzanol, and phytic acid. Food Sci. Nutr. 2014, 2, 75–104. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.M.; Jung, T.K.; Kim, S.H. Evaluation of allelopathic potential and quantification of momilactone A, B from rice hull extracts and assessment of inhibitory bioactivity on paddy field weeds. J. Agric. Food Chem. 2006, 54, 2527–2536. [Google Scholar] [CrossRef] [PubMed]
- Xuan, T.D.; Minh, T.N.; Anh, L.H.; Khanh, T.D. Allelopathic momilactones A and B are implied in rice drought and salinity tolerance, not weed resistance. Agron. Sustain. Dev. 2016, 36, 52. [Google Scholar] [CrossRef]
- Bourgaud, F.; Gravot, A.; Milesi, S.; Gontier, E. Production of plant secondary metabolites: A historical perspective. Plant Sci. 2001, 161, 839–851. [Google Scholar] [CrossRef]
- Kakar, K.; Xuan, T.D.; Quan, N.V.; Wafa, I.K.; Tran, H.D.; Khanh, T.D.; Dat, T.D. Effcacy of N-Methyl-N-Nitrosourea mutation on physicochemical properties, phytochemicals, and momilactones A and B in rice. Sustainability 2019, 11, 6862. [Google Scholar] [CrossRef]
- Tian, S.; Nakamura, K.; Kayahara, H. Analysis of phenolic compounds in white rice, brown rice, and germinated brown rice. J. Agric. Food Chem. 2004, 52, 4808–4813. [Google Scholar] [CrossRef]
- Ti, H.; Zhang, R.; Zhang, M.; Li, Q.; Wei, Z.; Zhang, Y.; Tang, X.; Deng, Y.; Liu, L.; Ma, Y. Dynamic changes in the free and bound phenolic compounds and antioxidant activity of brown rice at different germination stages. Food Chem. 2014, 161, 337–344. [Google Scholar] [CrossRef]
- Lien, D.T.P.; Tram, P.T.B.; Toan, H.T. Effects of extraction process on phenolic content and antioxidant activity of soybean. J. Food Nutr. Sci. 2015, 3, 33–38. [Google Scholar]
- Ly, D.; Lee, S.G. Comparisons of phenolic compounds, flavonoids contents and antioxidant activity in eco-friendly cultivated and conventional cultivated rice. Free Radic. Antioxid. 2014, 141, 378–385. [Google Scholar]
- Patil, S.B.; Khan, M.K. Germinated brown rice as a value-added rice product: A review. J. Food Sci. Technol. 2011, 48, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Zhai, X.; Zhang, N.; Tan, B. Effects of germination, fermentation and extrusion on the nutritional, cooking and sensory properties of brown rice products: A comparative study. Foods 2023, 12, 1542. [Google Scholar] [CrossRef] [PubMed]
- Palermo, M.; Pellegrini, N.; Fogliano, V. The effect of cooking on the phytochemical content of vegetables. J. Sci. Food Agric. 2014, 94, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Lindemann, I.; Dittgen, C.L.; de Souza Batista, C.; Dos Santos, J.P.; Bruni, G.P.; Elias, M.C.; Vanier, N.L. Rice and common bean blends: Effect of cooking on in vitro starch digestibility and phenolics profile. Food Chem. 2021, 340, 127908. [Google Scholar] [CrossRef] [PubMed]
- Aires-de-Sousa, M. Methicillin-resistant staphylococcus aureus among animals: Current overview. Clin. Microbiol. Infect. 2017, 23, 373–380. [Google Scholar] [CrossRef]
- Iloki-Assanga, S.B.; Lewis-Luján, L.M.; Lara-Espinoza, C.L.; Gil-Salido, A.A.; Fernandez-Angulo, D.; Rubio-Pino, J.L.; Haines, D.D. Solvent effects on phytochemical constituent profiles and antioxidant activities, using four different extraction formulations for analysis of Bucida buceras L. and Phoradendron californicum. BMC Res. Notes 2015, 8, 396. [Google Scholar] [CrossRef]
- Metrouh-Amir, H.; Duarte, C.M.M.; Maiza, F. Solvent effect on total phenolic contents, antioxidant, and antibacterial activities of Matricaria pubescens. Ind. Crops Prod. 2015, 67, 249–256. [Google Scholar] [CrossRef]
- Chan, A.P.L.; Chan, T.Y.K. Methanol as an unlisted ingredient in supposedly alcohol-based hand rub can pose serious health risk. Int. J. Environ. Res. Public Health 2018, 15, 1440. [Google Scholar] [CrossRef]
- Alzeer, J.; Abou Hadeed, K. Ethanol and its halal status in food industries. Trends Food Sci. Technol. 2016, 58, 14–20. [Google Scholar] [CrossRef]
- ElGamal, R.; Song, C.; Rayan, A.M.; Liu, C.; Al-Rejaie, S.; ElMasry, G. Thermal degradation of bioactive compounds during drying process of horticultural and agronomic products: A comprehensive overview. Agronomy 2023, 13, 1580. [Google Scholar] [CrossRef]
- Min, B.; Gu, L.; McClung, A.M.; Bergman, C.J.; Chen, M.H. Free and bound total phenolic concentrations, antioxidant capacities, and profiles of proanthocyanidins and anthocyanins in whole grain rice (Oryza sativa L.) of different bran colours. Food Chem. 2012, 133, 715–722. [Google Scholar] [CrossRef]
- Lee, C.W.; Yoneyama, K.; Takeuchi, Y.; Konnai, M.; Tamogami, S.; Kodama, O. Momilactones A and B in rice straw harvested at different growth stages. Biosci. Biotechnol. Biochem. 1999, 63, 1318–1320. [Google Scholar] [CrossRef]
- Quan, N.V.; Xuan, T.D.; Tran, H.D.; Thuy, N.T.D. Inhibitory activities of momilactones A, B, E, and 7-ketostigmasterol isolated from rice husk on paddy and invasiveweeds. Plants 2019, 8, 159. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).