

Separation and Biological Activities of the Main Compounds from the Bark of Myrica rubra Siebold & Zucc
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Reagents, and Chemicals
2.2. Preparation of Crude Extract
2.3. Partition Coefficients and HSSCC Separation
2.4. HPLC Analysis and Identification of Compounds
2.5. Assay for α-Glucosidase Inhibitory Activity
2.6. Evaluation of LPS-Induced Reactive Oxygen Species Scavenging Activity in Zebrafish Embryos
2.7. Apoptosis-Inducing Activities of Pure Myrica rubra Compounds
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
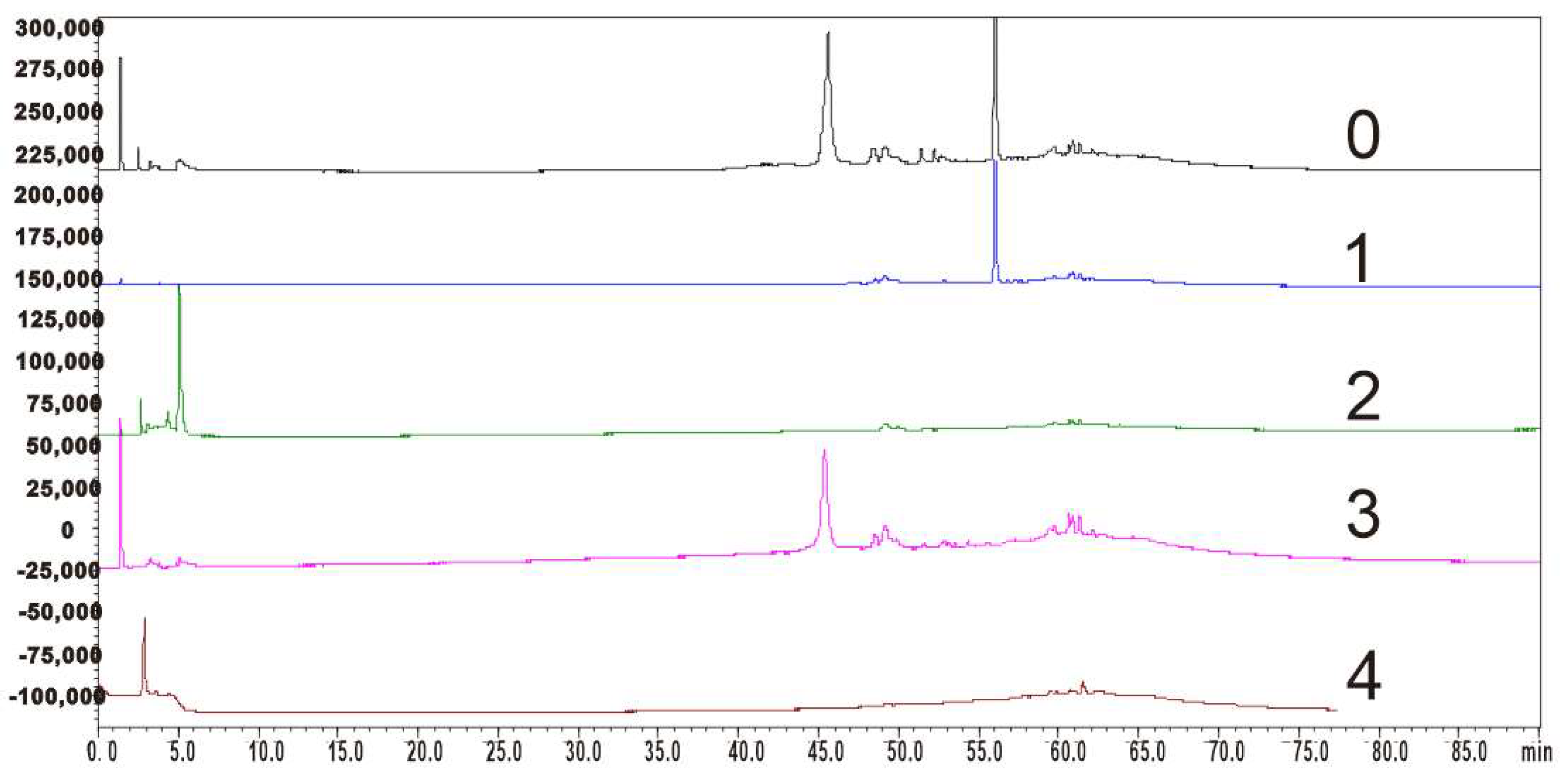
3.1. K Values in Solvent System and HSCCC Separation
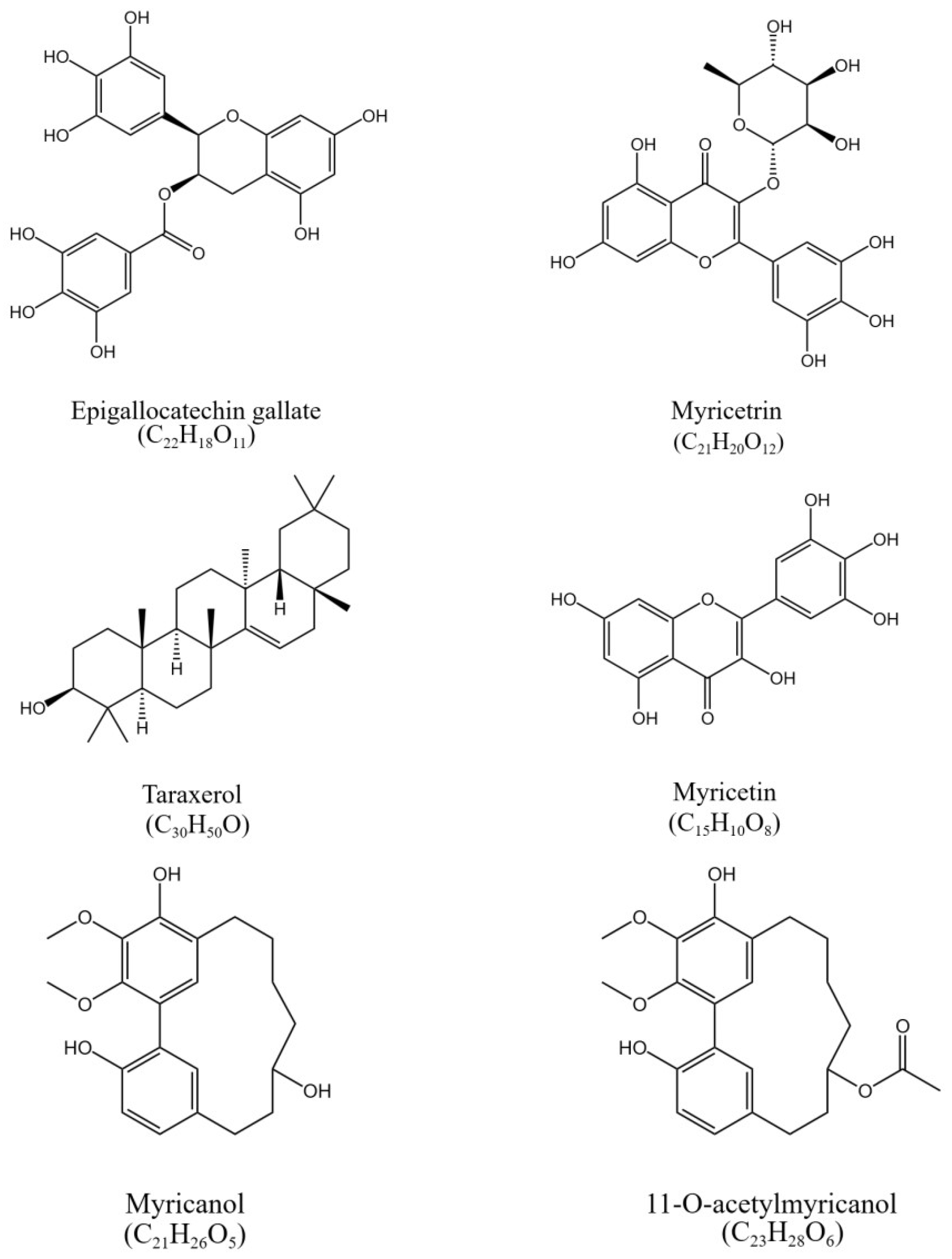
3.2. Purity and Structure Identification of HSCCC Peak Fractions
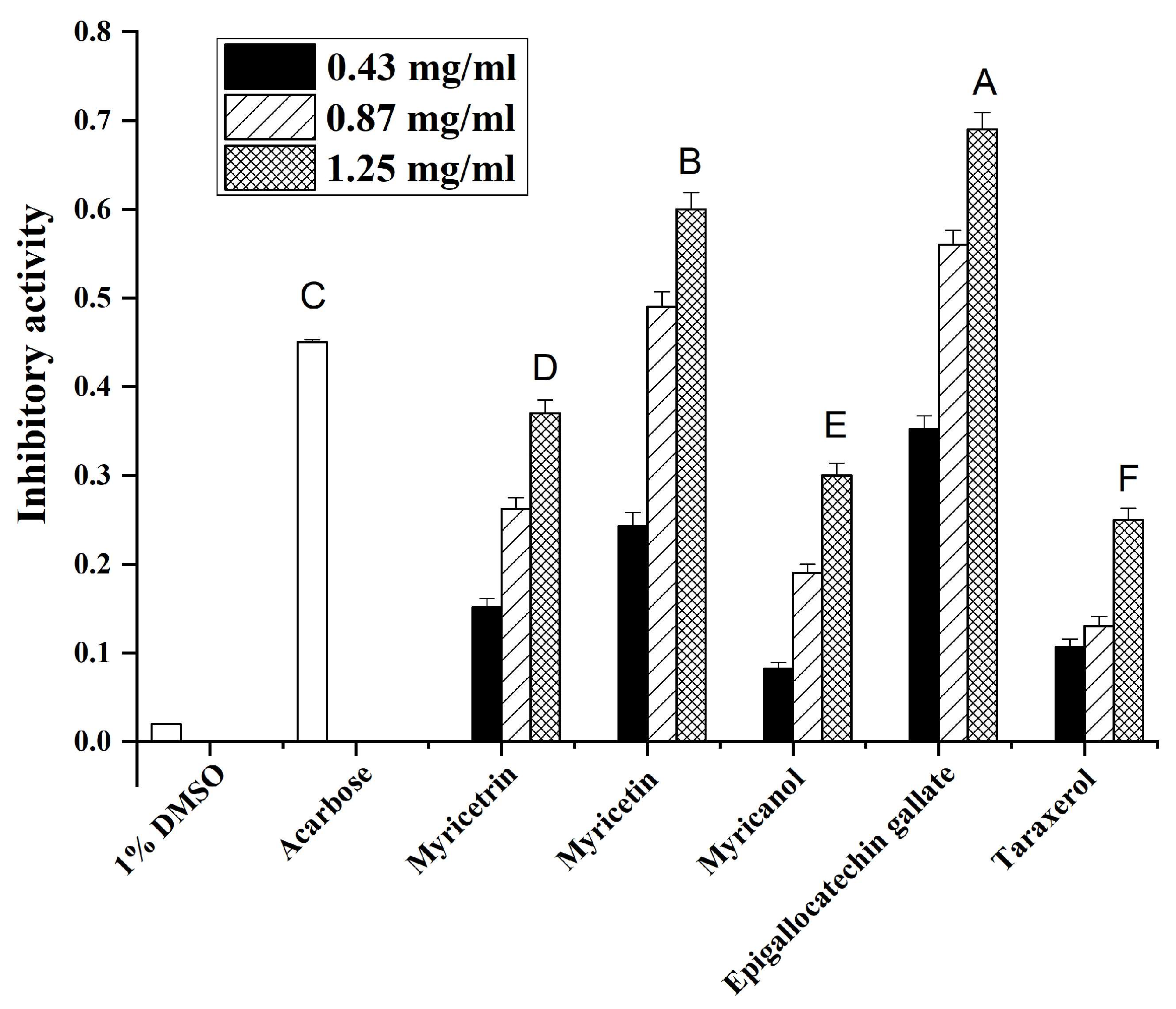
3.3. Effects of Main Compounds from Myrica rubra on α-Glucosidase Activity
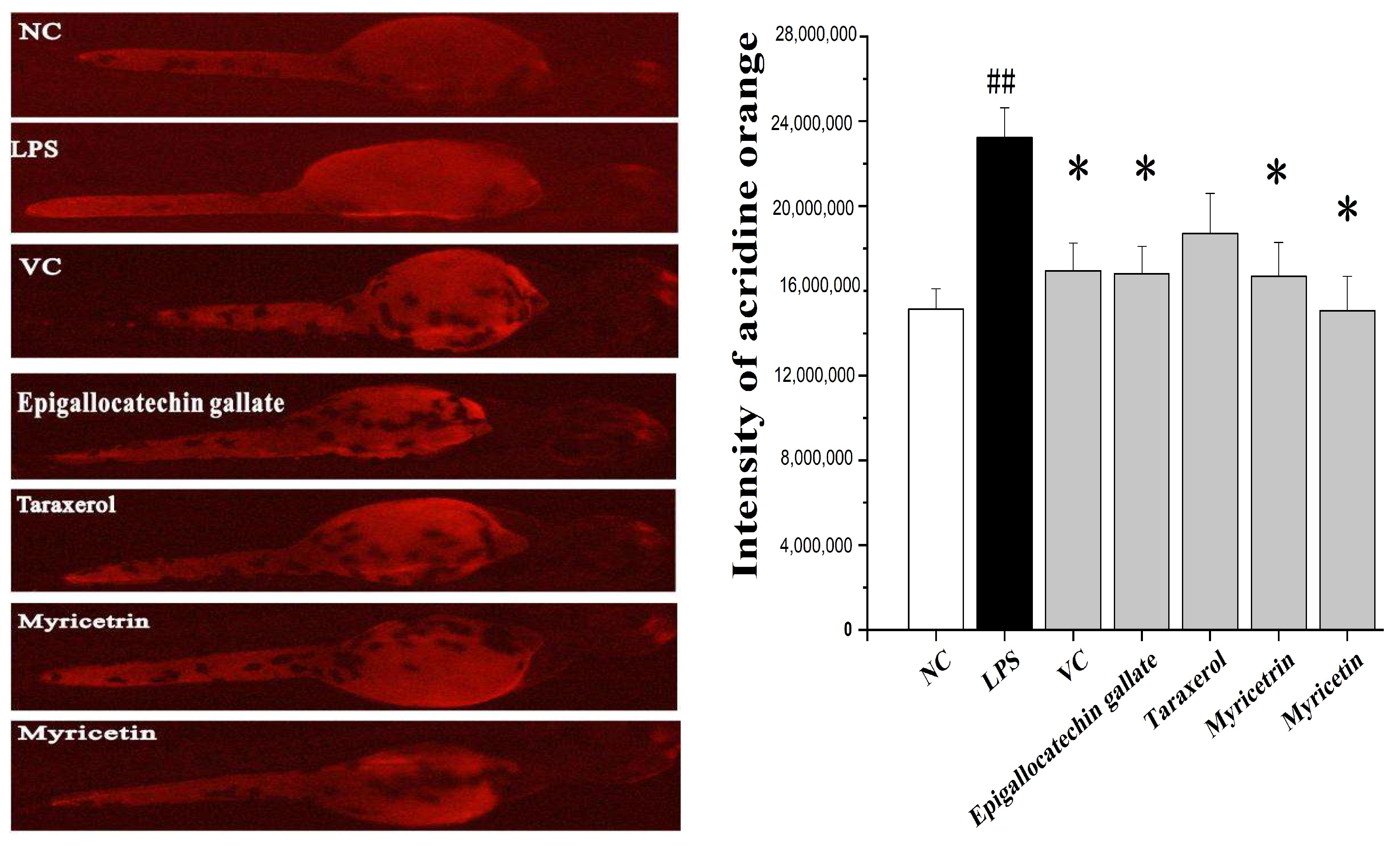
3.4. Reactive Oxygen Species Scavenging Activity in Zebrafish Embryos
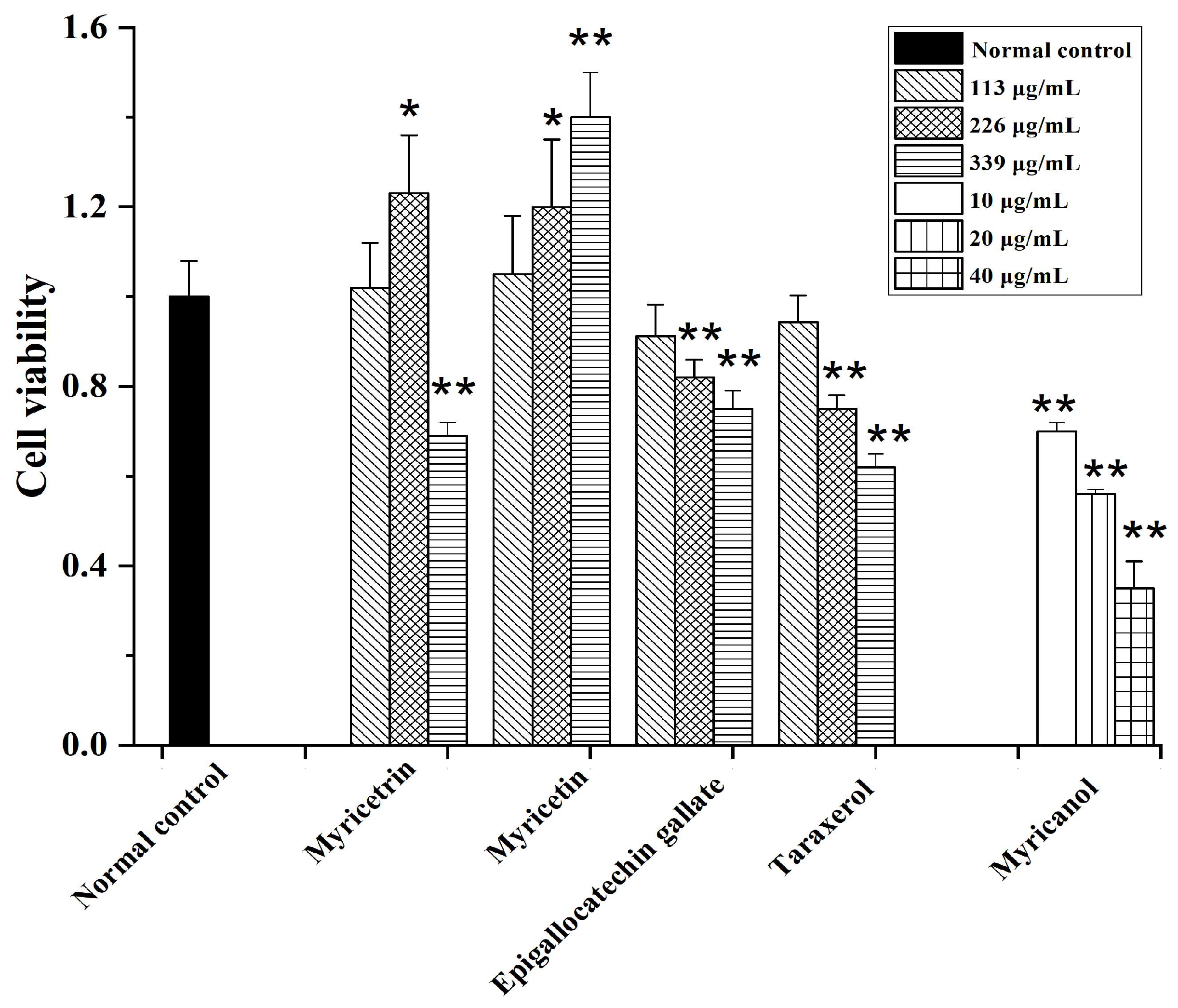
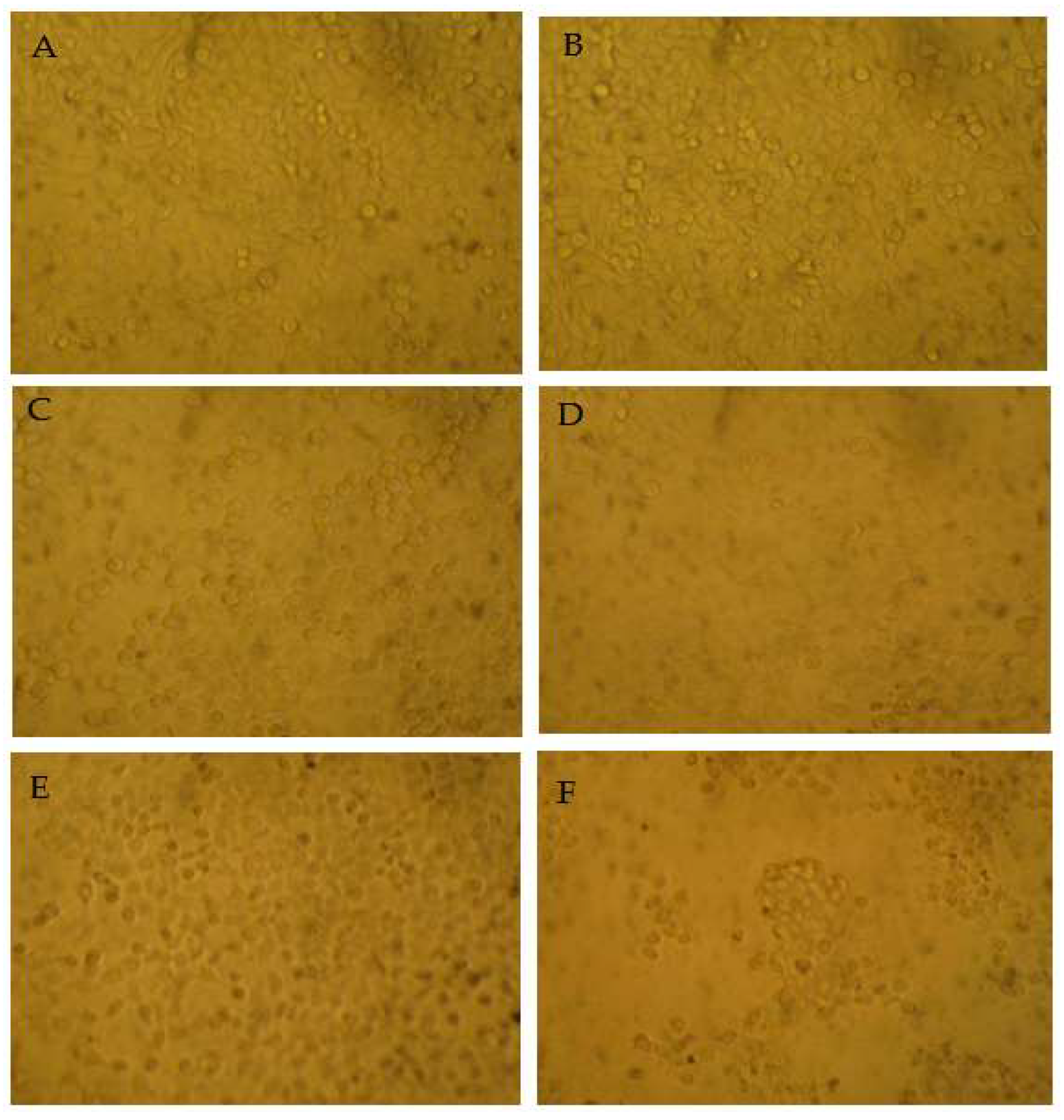
3.5. Apoptosis-Inducing Activities of Main Compounds from Myrica rubra on HepG2 Cells
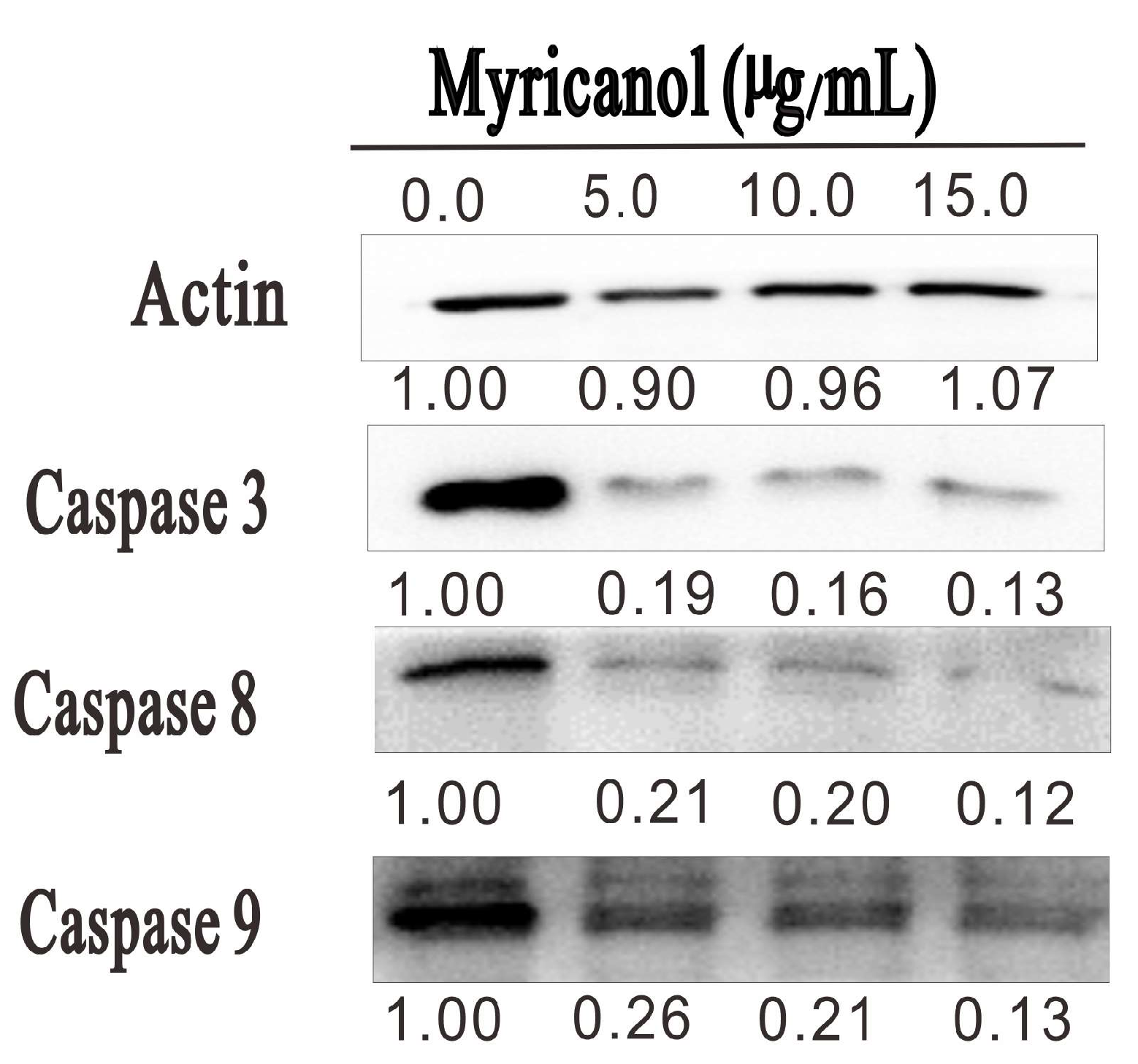
3.6. Myricanol Triggered Caspase-Dependent Apoptosis in HepG2 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ren, H.; He, Y.; Qi, X.; Zheng, X.; Zhang, S.; Yu, Z.; Hu, F. The bayberry database: A multiomic database for Myrica rubra, an important fruit tree with medicinal value. BMC Plant Biol. 2021, 21, 452. [Google Scholar] [CrossRef]
- Cheng, H.; Chen, Y.; Chen, Y.; Qin, D.; Ye, X.; Chen, J. Comparison and evaluation of aroma-active compounds for different squeezed Chinese bayberry (Myrica rubra) juices. J. Food Process. Preserv. 2021, 45, e15924. [Google Scholar] [CrossRef]
- Xia, W.; Gong, E.; Lin, Y.; Li, T.; Lian, F.; Zheng, B.; Liu, R.H. Comparison of phytochemical profiles, antioxidant and antiproliferative activities in Chinese bayberry (Myrica rubra Sieb. et Zucc.) fruits. J. Food Sci. 2021, 86, 4691–4703. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, J.; Lu, S.; Tang, W.; Zhou, Y.; Quek, S.Y. Effects of different drying methods on phenolic components and in vitro hypoglycemic activities of pulp extracts from two Chinese bayberry (Myrica rubra Sieb. et Zucc.) cultivars. Food Sci. Hum. Wellness 2022, 11, 366–373. [Google Scholar] [CrossRef]
- Tao, J.; Morikawa, T.; Toguchida, I.; Ando, S.; Matsuda, H.; Yoshikawa, M. Inhibitors of nitric oxide production from the bark of Myrica rubra: Structures of new biphenyl type diarylheptanoid glycosides and taraxerane type triterpene. Bioorgan. Med. Chem. 2002, 10, 4005–4012. [Google Scholar] [CrossRef]
- Wang, M.; Liu, Y.; Pan, R.L.; Wang, R.Y.; Ding, S.L.; Dong, W.R.; Sun, G.B.; Ye, J.X.; Sun, X.B. Protective effects of Myrica rubra flavonoids against hypoxia/reoxygenation-induced cardiomyocyte injury via the regulation of the PI3K/Akt/GSK3β pathway. Int. J. Mol. Med. 2019, 43, 2133–2143. [Google Scholar] [CrossRef]
- Yu, H.; Fan, Z.F.; Xu, J.N.; Lv, X.K. Anti-inflammatory and analgesic effect of Wujiapi and Yangmeishupi Compound Preparations on animals. J. North China Univ. Sci. Technol. 2021, 23, 444–449. [Google Scholar]
- Li, Z.; Shi, W.; Cheng, L.; Pan, S.; Wang, C. Screening of the phenolic profile and their antioxidative activities of methanol extracts of Myrica rubra fruits, leaves and bark. J. Food Meas. Charact. 2018, 12, 128–134. [Google Scholar] [CrossRef]
- Shen, S.; Zhao, M.; Li, C.; Chang, Q.; Liu, X.; Liao, Y.; Pan, R. Study on the material basis of neuroprotection of Myrica rubra bark. Molecules 2019, 24, 2993. [Google Scholar] [CrossRef]
- Wang, D.Y.; Liu, E.-G. A new diaryheptanoid from the bark of Myrica rubra. Nat. Prod. Res. 2008, 22, 292–295. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, J.; Chen, F.; Cheng, Y.; Guo, A. Separation, Identification, and Quantitation of Phenolic Acids in Chinese Waxberry (Myrica rubra) Juice by HPLC-PDA-ESI-MS. J. Food Sci. 2012, 77, C272–C277. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhao, J.; Yang, T.; Sun, B. High-speed countercurrent chromatography as an efficient technique for large separation of plant polyphenols: A review. Food. Res. Int. 2022, 153, 110956. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Ye, X.; Li, X.; Chen, H.; Yuan, L.; Deng, Y.; Chen, X.; Li, X. Separation of two constituents from purple sweet potato by combination of silica gel column and high-speed counter-current chromatography. J. Chromatogr. B 2012, 881, 49–54. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Li, X.; Chen, X.; Ye, X.; Huang, J.; Jin, Y.; Li, P.; Deng, Y.; Jin, Q.; Shi, Q. Evaluation of antidiabetic potential of selected traditional Chinese medicines in STZ-induced diabetic mice. J. Ethnopharmacol. 2011, 137, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Zhou, Y.; Yin, X.; Wei, X.; Zhou, Y. Two New Phenolic Glycosides with Lactone Structural Units from Leaves of Ardisia crenata Sims with Antibacterial and Anti-Inflammatory Activities. Molecules 2022, 27, 4903. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.; Ma, Y.; Lin, W.; Xu, K.; Chen, M. Study on the structure–activity of dihydromyricetin and its new production: Myricetin by using ESR and spectroscopic techniques. J. Therm. Anal. Calorim. 2014, 116, 241–248. [Google Scholar] [CrossRef]
- Kawai, S.; Nakata, K.; Ohashi, M.; Nishida, T. Myricanol and myricanone biosynthesis in Myrica rubra: Incorporation of two molecules of 4-coumaric acid. J. Wood Sci. 2008, 54, 256–260. [Google Scholar] [CrossRef]
- Peres, I.; Rocha, S.; do Carmo Pereira, M.; Coelho, M.; Rangel, M.; Ivanova, G. NMR structural analysis of epigallocatechin gallate loaded polysaccharide nanoparticles. Carbohyd. Polym. 2010, 82, 861–866. [Google Scholar] [CrossRef]
- Alamzeb, M.; Ali, S.; Khan, A.A.; Igoli, J.O.; Ferro, V.A.; Gray, A.I.; Khan, M.R. A new ceramide along with eight known compounds from the roots of Artemisia incisa Pamp. Rec. Nat. Prod. 2015, 9, 297–304. [Google Scholar]
- Xu, Y.; Rashwan, A.K.; Ge, Z.; Li, Y.; Ge, H.; Li, J.; Xie, J.; Liu, S.; Fang, J.; Cheng, K. Identification of a novel α-glucosidase inhibitor from Melastoma dodecandrum Lour. fruits and its effect on regulating postprandial blood glucose. Food Chem. 2023, 399, 133999. [Google Scholar] [CrossRef]
- Zhang, H.; Guo, Q.; Liang, Z.; Wang, M.; Wang, B.; Sun-Waterhouse, D.; Waterhouse, G.I.; Wang, J.; Ma, C.; Kang, W. Anti-inflammatory and antioxidant effects of Chaetoglobosin Vb in LPS-induced RAW264. 7 cells: Achieved via the MAPK and NF-κB signaling pathways. Food Chem. Toxicol. 2021, 147, 111915. [Google Scholar] [CrossRef] [PubMed]
- Riedl, S.J.; Shi, Y. Molecular mechanisms of caspase regulation during apoptosis. Nat. Rev. Mol. Cell Biol. 2004, 5, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R.; McCallum, J. Chemistry of flavonoids. Fruit Veg. Phytochem. 2010, 131, 357–367. [Google Scholar]
- Dludla, P.V.; Mabhida, S.E.; Ziqubu, K.; Nkambule, B.B.; Mazibuko-Mbeje, S.E.; Hanser, S.; Basson, A.K.; Pheiffer, C.; Kengne, A.P. Pancreatic β-cell dysfunction in type 2 diabetes: Implications of inflammation and oxidative stress. World J. Diabetes 2023, 14, 130. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, J.S.; Sehrawat, A.; Mishra, J.; Sidhu, I.S.; Navik, U.; Khullar, N.; Kumar, S.; Bhatti, G.K.; Reddy, P.H. Oxidative stress in the pathophysiology of type 2 diabetes and related complications: Current therapeutics strategies and future perspectives. Free Radic. Biol. Med. 2022, 184, 114–134. [Google Scholar] [CrossRef] [PubMed]
- Dai, T.; Li, T.; He, X.; Li, X.; Liu, C.; Chen, J.; McClements, D.J. Analysis of inhibitory interaction between epigallocatechin gallate and alpha-glucosidase: A spectroscopy and molecular simulation study. Spectrochim. Acta A 2020, 230, 118023. [Google Scholar] [CrossRef]
- Chang, G.; Cai, C.; Xiang, Y.; Fang, X.; Yang, H. Extraction and study of hypoglycemic constituents from Myrica rubra pomace. Molecules 2022, 27, 846. [Google Scholar] [CrossRef]
- Zhang, Q.; Huang, Z.; Wang, Y.; Wang, Y.; Fu, L.; Su, L. Chinese bayberry (Myrica rubra) phenolics mitigated protein glycoxidation and formation of advanced glycation end-products: A mechanistic investigation. Food Chem. 2021, 361, 130102. [Google Scholar] [CrossRef]
- Kandasamy, N.; Ashokkumar, N. Renoprotective effect of myricetin restrains dyslipidemia and renal mesangial cell proliferation by the suppression of sterol regulatory element binding proteins in an experimental model of diabetic nephropathy. Eur. J. Pharmacol. 2014, 743, 53–62. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, Y.; Xiang, W.; Wang, Q.; Cao, Y. Molecular mechanisms of the action of myricetin in cancer. Mini-Rev. Med. Chem. 2020, 20, 123–133. [Google Scholar] [CrossRef]
- Fan, S.; Wang, C.; Huang, K.; Liang, M. Myricanol Inhibits Platelet Derived Growth Factor-BB-Induced Vascular Smooth Muscle Cells Proliferation and Migration in vitro and Intimal Hyperplasia in vivo by Targeting the Platelet-Derived Growth Factor Receptor-β and NF-κB Signaling. Front. Physiol. 2022, 12, 2574. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Liao, Q.; Feng, Y.; Liu, J.; Pan, R.; Lee, S.M.-Y.; Lin, L. Myricanol mitigates lipid accumulation in 3T3-L1 adipocytes and high fat diet-fed zebrafish via activating AMP-activated protein kinase. Food Chem. 2019, 270, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Liao, Q.; Zhang, T.; Pan, R.; Lin, L. Myricanol modulates skeletal muscle–adipose tissue crosstalk to alleviate high-fat diet-induced obesity and insulin resistance. Br. J. Pharmacol. 2019, 176, 3983–4001. [Google Scholar] [CrossRef] [PubMed]
- Kesavardhana, S.; Malireddi, R.S.; Kanneganti, T.D. Caspases in cell death, inflammation, and pyroptosis. Annu. Rev. Immunol. 2020, 38, 567–595. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, T.; Fan, L.; Chang, Y.; Yang, H.; He, K. Separation and Biological Activities of the Main Compounds from the Bark of Myrica rubra Siebold & Zucc. Separations 2024, 11, 4. https://doi.org/10.3390/separations11010004
Hao T, Fan L, Chang Y, Yang H, He K. Separation and Biological Activities of the Main Compounds from the Bark of Myrica rubra Siebold & Zucc. Separations. 2024; 11(1):4. https://doi.org/10.3390/separations11010004
Chicago/Turabian StyleHao, Tianyang, Lingyang Fan, Yiyue Chang, Hui Yang, and Kai He. 2024. "Separation and Biological Activities of the Main Compounds from the Bark of Myrica rubra Siebold & Zucc" Separations 11, no. 1: 4. https://doi.org/10.3390/separations11010004
APA StyleHao, T., Fan, L., Chang, Y., Yang, H., & He, K. (2024). Separation and Biological Activities of the Main Compounds from the Bark of Myrica rubra Siebold & Zucc. Separations, 11(1), 4. https://doi.org/10.3390/separations11010004