
Decrease in Ca2+ Concentration in Quail Cardiomyocytes Is Faster than That in Rat Cardiomyocytes


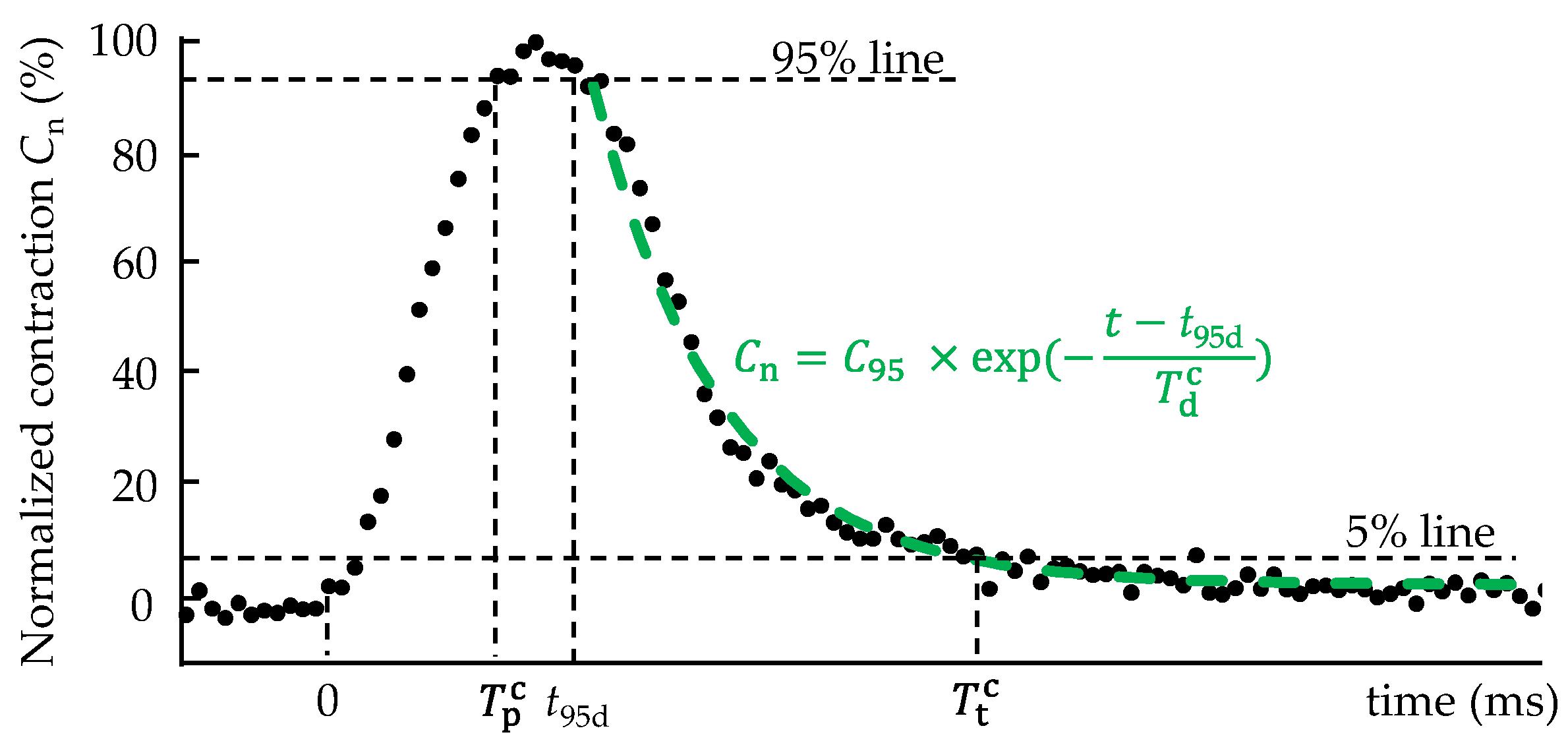{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Isolation of Cardiomyocytes
2.3. Experimental Setup
2.4. Analysis of Cell Morphology and Contraction–Relaxation Behavior
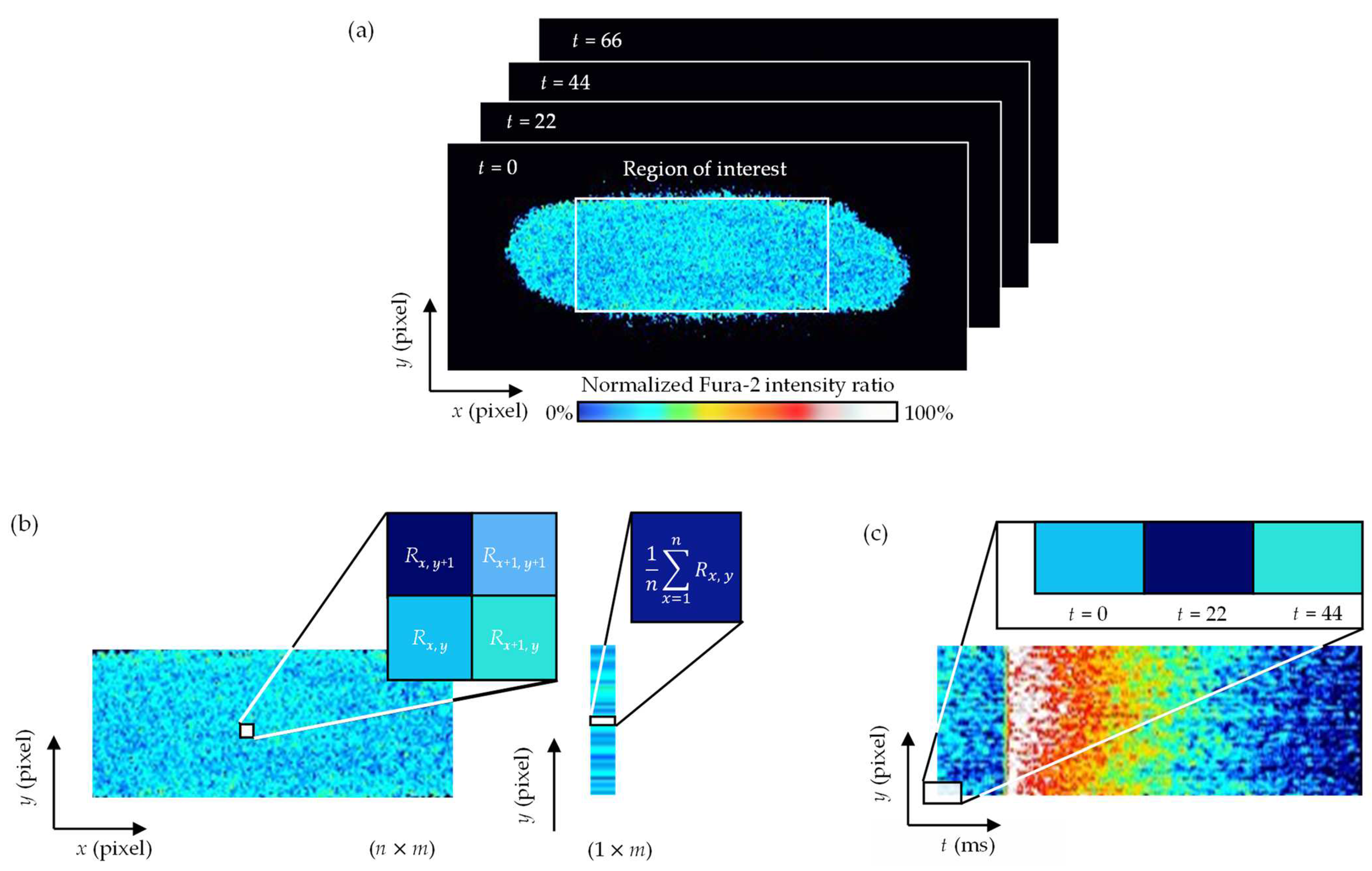
2.5. Analysis of Ca2+ Concentration Changes at the Whole-Cell Level
2.6. Analysis of the Spatial Distributions of Changes in Ca2+ Concentration
2.7. Statistical Method
3. Results
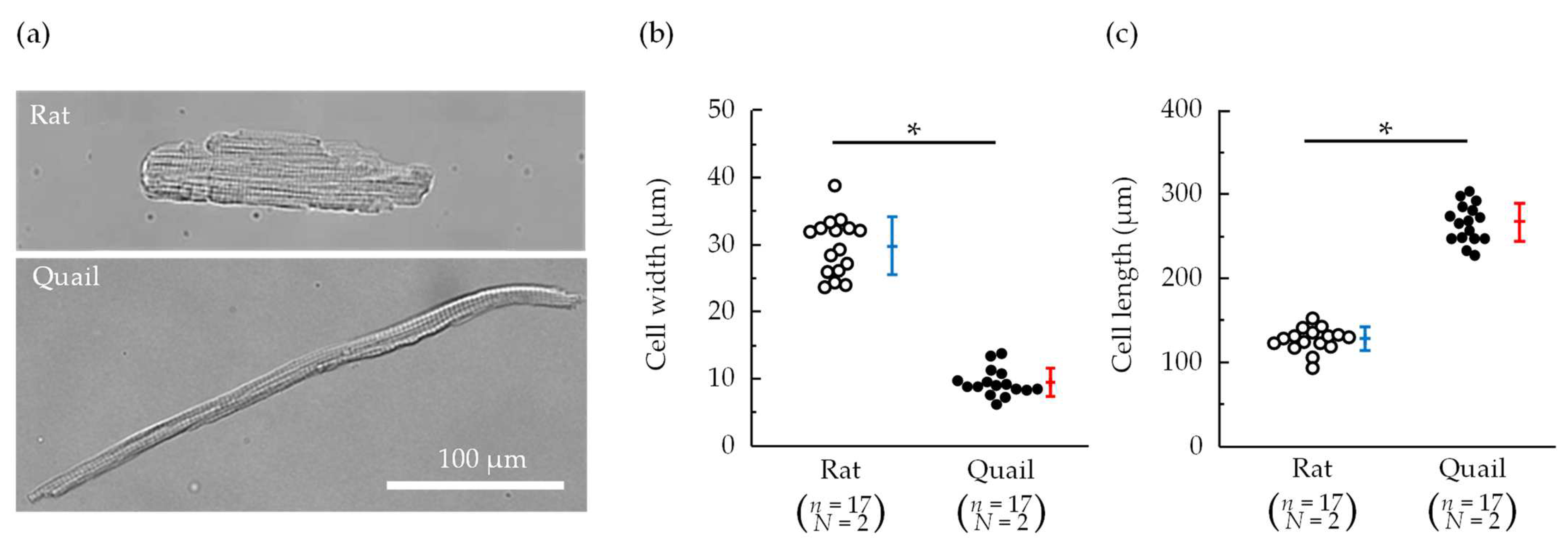
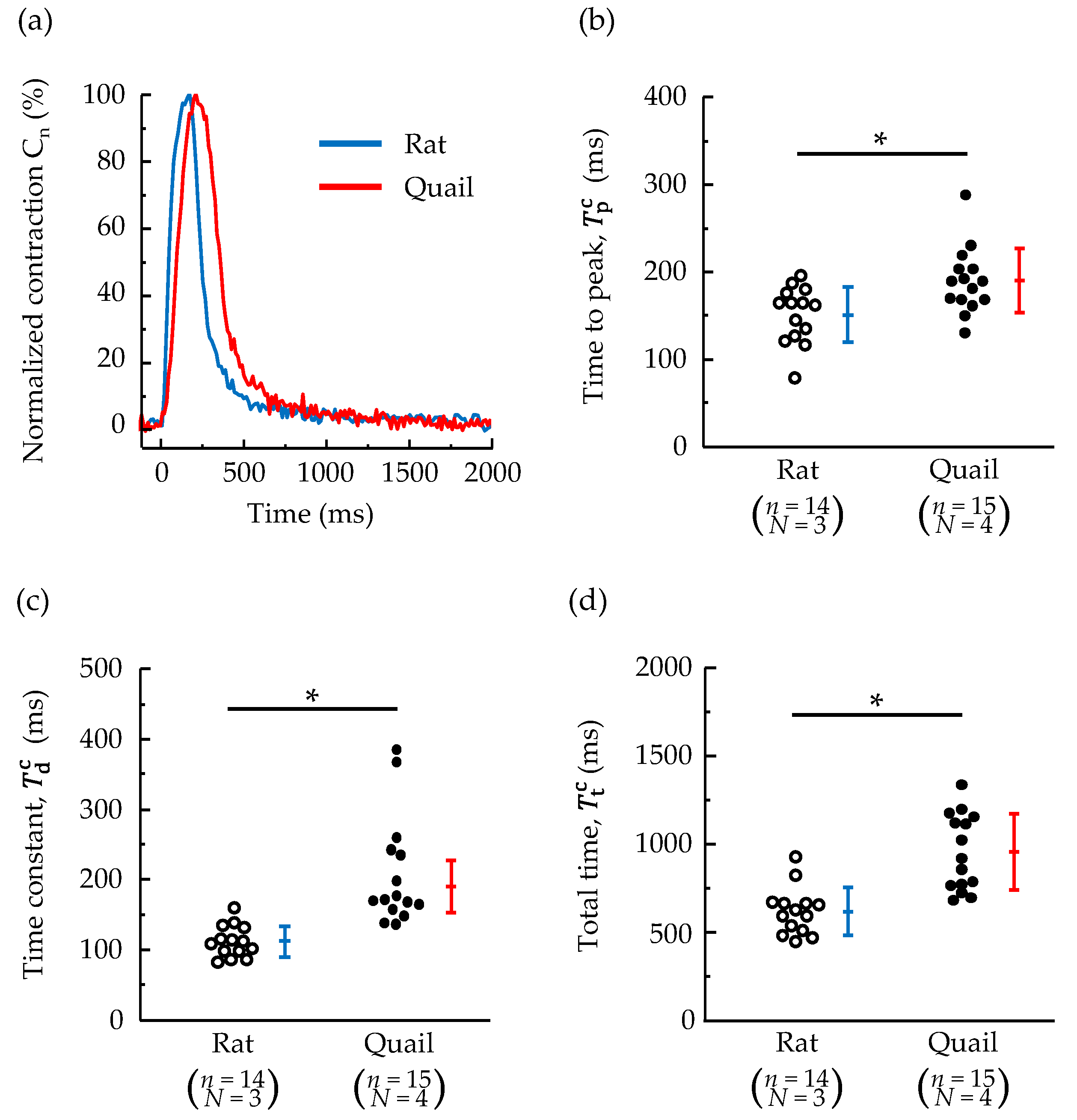
3.1. Morphology and Contraction—Relaxation Behavior of Isolated Cardiomyocytes
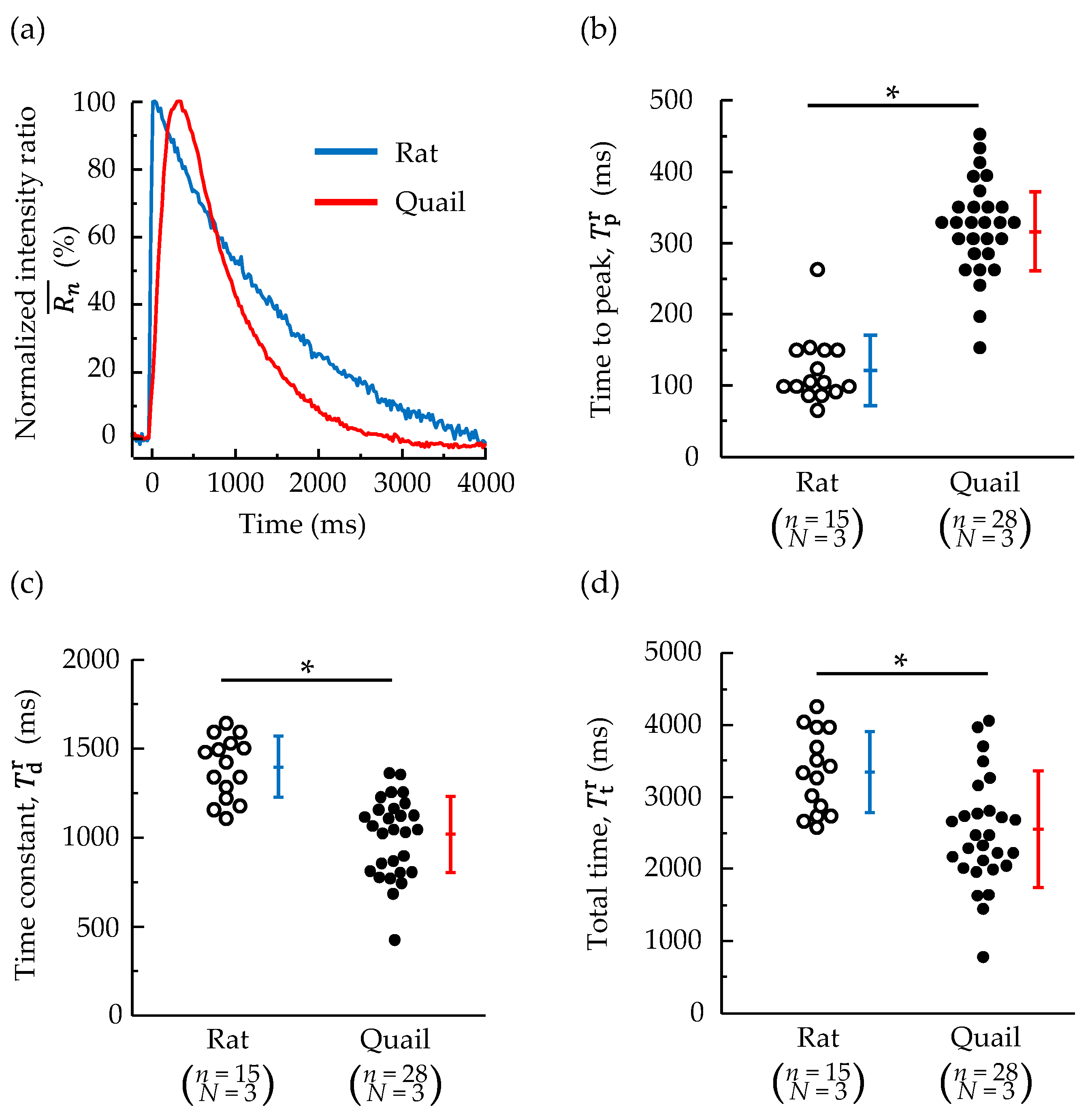
3.2. Changes in Whole-Cell Ca2+ Concentration
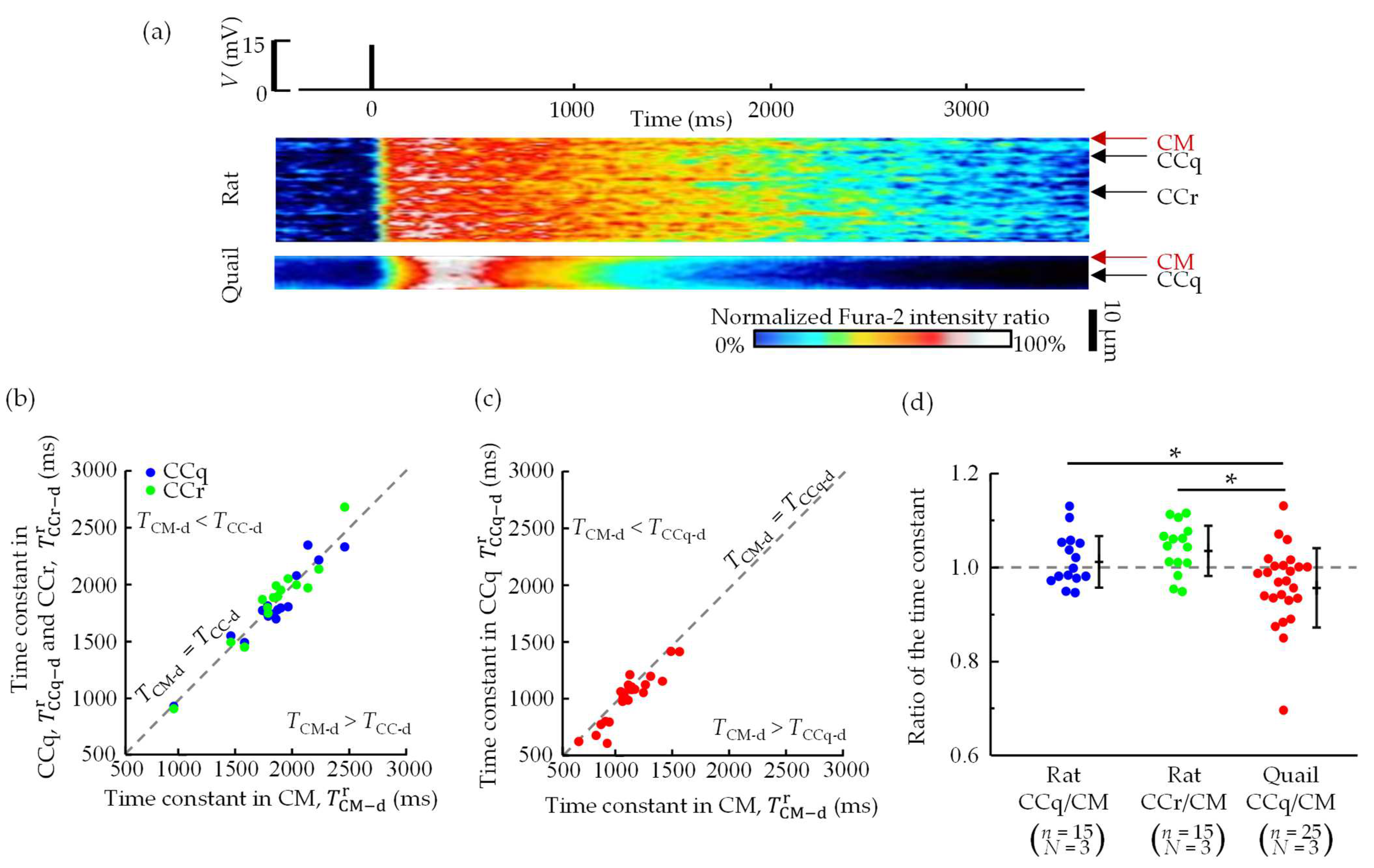
3.3. Spatial Distributions of Ca2+ Concentration
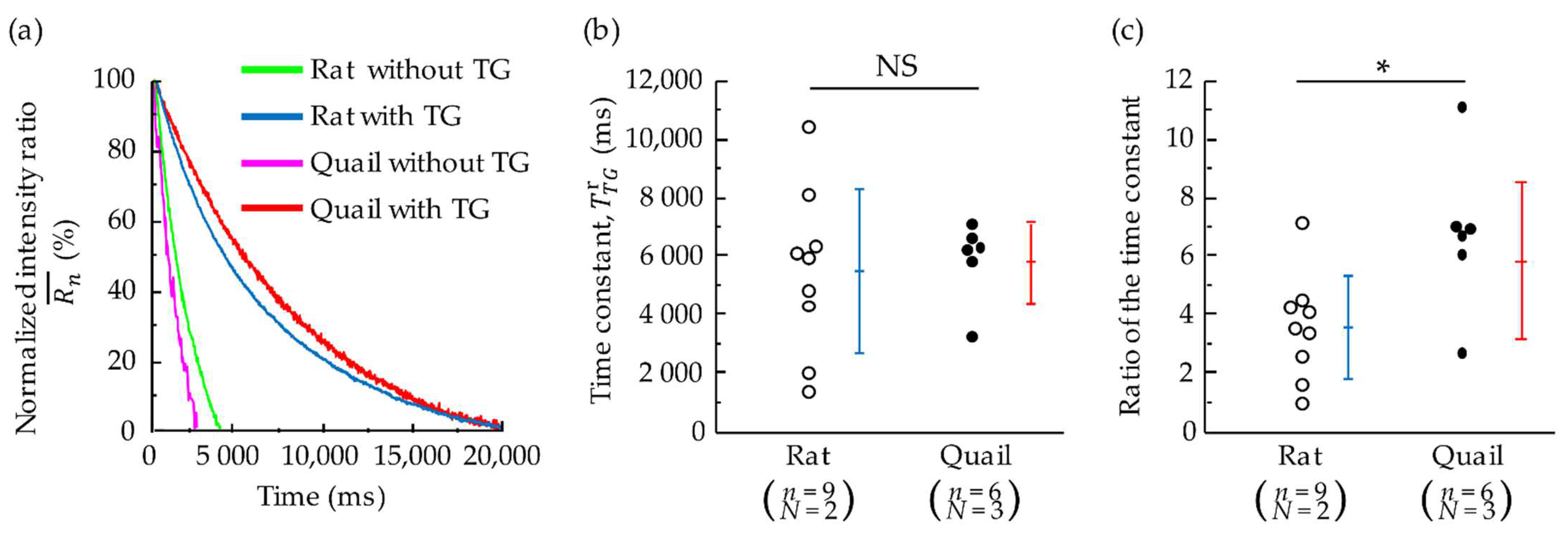
3.4. Effects of SERCA Inhibition on the Decay Speed after Peak of Whole-Cell Ca2+ Concentration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bers, D.M. Cardiac excitation-contraction coupling. Nature 2002, 415, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Bassani, J.W.; Bassani, R.A.; Bers, D.M. Relaxation in rabbit and rat cardiac cells: Species-dependent differences in cellular mechanisms. J. Physiol. 1994, 476, 279–293. [Google Scholar] [CrossRef]
- Johnson, E.A.; Sommer, J.R. A strand of cardiac muscle. Its ultrastructure and the electrophysiological implications of its geometry. J. Cell Biol. 1967, 33, 103–129. [Google Scholar] [CrossRef] [PubMed]
- Takeshima, H.; Komazaki, S.; Nishi, M.; Iino, M.; Kangawa, K. Junctophilins: A novel family of junctional membrane complex proteins. Mol. Cell 2000, 6, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Louch, W.E.; Hake, J.; Jølle, G.F.; Mørk, H.K.; Sjaastad, I.; Lines, G.T.; Sejersted, O.M. Control of Ca2+ release by action potential configuration in normal and failing murine cardiomyocytes. Biophys. J. 2010, 99, 1377–1386. [Google Scholar] [CrossRef]
- Dibb, K.M.; Louch, W.E.; Trafford, A.W. Cardiac Transverse Tubules in Physiology and Heart Failure. Annu. Rev. Physiol. 2021, 84, 229–255. [Google Scholar] [CrossRef]
- Shiels, H.A.; Galli, G.L. The sarcoplasmic reticulum and the evolution of the vertebrate heart. Physiology 2014, 29, 456–469. [Google Scholar] [CrossRef]
- Franzini-Armstrong, C.; Protasi, F.; Tijskens, P. The assembly of calcium release units in cardiac muscle. Ann. N. Y. Acad. Sci. 2005, 1047, 76–85. [Google Scholar] [CrossRef]
- Honda, T.; Ujihara, Y.; Hanashima, A.; Hashimoto, K.; Tanemoto, K.; Mohri, S. Turtle spongious ventricles exhibit more compliant diastolic property and possess larger elastic regions of connectin in comparison to rat compact left ventricles. Kawasaki Med. J. 2018, 44, 1–17. [Google Scholar] [CrossRef]
- Grubb, B.R. Allometric relations of cardiovascular function in birds. Am. J. Physiol. 1983, 245, H567–H572. [Google Scholar] [CrossRef]
- Sheard, T.M.D.; Kharche, S.R.; Pinali, C.; Shiels, H.A. 3D ultrastructural organisation of calcium release units in the avian sarcoplasmic reticulum. J. Exp. Biol. 2019, 222, jeb197640. [Google Scholar] [CrossRef] [PubMed]
- Filatova, T.S.; Abramochkin, D.V.; Shiels, H.A. Warmer, faster, stronger: Ca2+ cycling in avian myocardium. J. Exp. Biol. 2020, 223, jeb228205. [Google Scholar] [CrossRef] [PubMed]
- Valance, D.; Desprès, G.; Boissy, A.; Mignon-Grasteau, S.; Constantin, P.; Leterrier, C. Genetic selection on a behavioural fear trait is associated with changes in heart rate variability in quail. Genes Brain Behav. 2007, 6, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Möller, C.; van Dijk, R.M.; Wolf, F.; Keck, M.; Schönhoff, K.; Bierling, V.; Potschka, H. Impact of repeated kindled seizures on heart rate rhythms, heart rate variability, and locomotor activity in rats. Epilepsy Behav. 2019, 92, 36–44. [Google Scholar] [CrossRef]
- Shioya, T. A simple technique for isolating healthy heart cells from mouse models. J. Physiol. Sci. 2007, 57, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Ujihara, Y.; Kanagawa, M.; Mohri, S.; Takatsu, S.; Kobayashi, K.; Toda, T.; Naruse, K.; Katanosaka, Y. Elimination of fukutin reveals cellular and molecular pathomechanisms in muscular dystrophy-associated heart failure. Nat. Commun. 2019, 10, 5754. [Google Scholar] [CrossRef]
- Sato, M.; Miyata, K.; Tian, Z.; Kadomatsu, T.; Ujihara, Y.; Morinaga, J.; Horiguchi, H.; Endo, M.; Zhao, J.; Zhu, S.; et al. Loss of Endogenous HMGB2 Promotes Cardiac Dysfunction and Pressure Overload-Induced Heart Failure in Mice. Circ. J. 2019, 83, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Kodama, A.; Ohira, M.; Kimoto, M.; Nakagawa, M.; Usui, Y.; Ujihara, Y.; Hanashima, A.; Mohri, S. Transient induction of cell cycle promoter Fam64a improves cardiac function through regulating Klf15-dependent cardiomyocyte differentiation in mice. BioRxiv 2021. [Google Scholar] [CrossRef]
- Illaste, A.; Wullschleger, M.; Fernandez-Tenorio, M.; Niggli, E.; Egger, M. Automatic Detection and Classification of Ca2+ Release Events in Line- and Frame-Scan Images. Biophys. J. 2019, 116, 383–394. [Google Scholar] [CrossRef]
- Katanosaka, Y.; Iwasaki, K.; Ujihara, Y.; Takatsu, S.; Nishitsuji, K.; Kanagawa, M.; Sudo, A.; Toda, T.; Katanosaka, K.; Mohri, S.; et al. TRPV2 is critical for the maintenance of cardiac structure and function in mice. Nat. Commun. 2014, 5, 3932. [Google Scholar] [CrossRef]
- Ujihara, Y.; Iwasaki, K.; Takatsu, S.; Hashimoto, K.; Naruse, K.; Mohri, S.; Katanosaka, Y. Induced NCX1 overexpression attenuates pressure overload-induced pathological cardiac remodelling. Cardiovasc. Res. 2016, 111, 348–361. [Google Scholar] [CrossRef] [PubMed]
- Scriven, D.R.; Moore, E.D. Ca²⁺ channel and Na⁺/Ca²⁺ exchange localization in cardiac myocytes. J. Mol. Cell. Cardiol. 2013, 58, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Ujihara, Y.; Mohri, S.; Katanosaka, Y. Effects of induced Na+/Ca2+ exchanger overexpression on the spatial distribution of L-type Ca2+ channels and junctophilin-2 in pressure-overloaded hearts. Biochem. Biophys. Res. Commun. 2016, 480, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Louch, W.E.; Mørk, H.K.; Sexton, J.; Strømme, T.A.; Laake, P.; Sjaastad, I.; Sejersted, O.M. T-tubule disorganization and reduced synchrony of Ca2+ release in murine cardiomyocytes following myocardial infarction. J. Physiol. 2006, 574 Pt 2, 519–533. [Google Scholar] [CrossRef]
- Øyehaug, L.; Loose, K.Ø.; Jølle, G.F.; Røe, Å.T.; Sjaastad, I.; Christensen, G.; Sejersted, O.M.; Louch, W.E. Synchrony of cardiomyocyte Ca2+ release is controlled by T-tubule organization, SR Ca2+ content, and ryanodine receptor Ca2+ sensitivity. Biophys. J. 2013, 104, 1685–1697. [Google Scholar] [CrossRef]
- Galli, G.L.; Taylor, E.W.; Shiels, H.A. Calcium flux in turtle ventricular myocytes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R1781–R1789. [Google Scholar] [CrossRef]
- Ito, M.; Ujihara, Y.; Sugita, S.; Nakamura, M. Comparison of the histology and stiffness of ventricles in Anura of different habitats. J. Biol. Phys. 2021, 47, 287–300. [Google Scholar] [CrossRef]
- Ito, M.; Sugita, S.; Nakamura, M.; Ujihara, Y. Differences in Pressure Resistance and Tissue Structure of Ventricles between Aquatic, Semiaquatic, and Terrestrial Anura. Trans. Jpn. Soc. Med. Biol. Eng. 2021, 59, 162–168. [Google Scholar]
- Akester, A.R. Intercalated discs, nexuses, sarcoplasmic reticulum and transitional cells in the heart of the adult domestic fowl (Gallus gallus domesticus). J. Anat. 1981, 133 Pt 2, 161–179. [Google Scholar]
- Kim, C.S.; Davidoff, A.J.; Maki, T.M.; Doye, A.A.; Gwathmey, J.K. Intracellular calcium and the relationship to contractility in an avian model of heart failure. J. Comp. Physiol. B 2000, 170, 295–306. [Google Scholar] [CrossRef][Green Version]
- Baylor, S.M.; Hollingworth, S. Calcium indicators and calcium signalling in skeletal muscle fibres during excitation-contraction coupling. Prog. Biophys. Mol. Biol. 2011, 105, 162–179. [Google Scholar] [CrossRef] [PubMed]
- Baylor, S.M.; Hollingworth, S. Fura-2 calcium transients in frog skeletal muscle fibres. J. Physiol. 1988, 403, 151–192. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogura, Y.; Ito, H.; Sugita, S.; Nakamura, M.; Ujihara, Y. Decrease in Ca2+ Concentration in Quail Cardiomyocytes Is Faster than That in Rat Cardiomyocytes. Processes 2022, 10, 508. https://doi.org/10.3390/pr10030508
Ogura Y, Ito H, Sugita S, Nakamura M, Ujihara Y. Decrease in Ca2+ Concentration in Quail Cardiomyocytes Is Faster than That in Rat Cardiomyocytes. Processes. 2022; 10(3):508. https://doi.org/10.3390/pr10030508
Chicago/Turabian StyleOgura, Yuhei, Hiroaki Ito, Shukei Sugita, Masanori Nakamura, and Yoshihiro Ujihara. 2022. "Decrease in Ca2+ Concentration in Quail Cardiomyocytes Is Faster than That in Rat Cardiomyocytes" Processes 10, no. 3: 508. https://doi.org/10.3390/pr10030508
APA StyleOgura, Y., Ito, H., Sugita, S., Nakamura, M., & Ujihara, Y. (2022). Decrease in Ca2+ Concentration in Quail Cardiomyocytes Is Faster than That in Rat Cardiomyocytes. Processes, 10(3), 508. https://doi.org/10.3390/pr10030508