Stochastic and Temporal Models of Olfactory Perception

Abstract
1. Introduction
1.1. Preamble
1.2. Odor Identification of Single Compounds
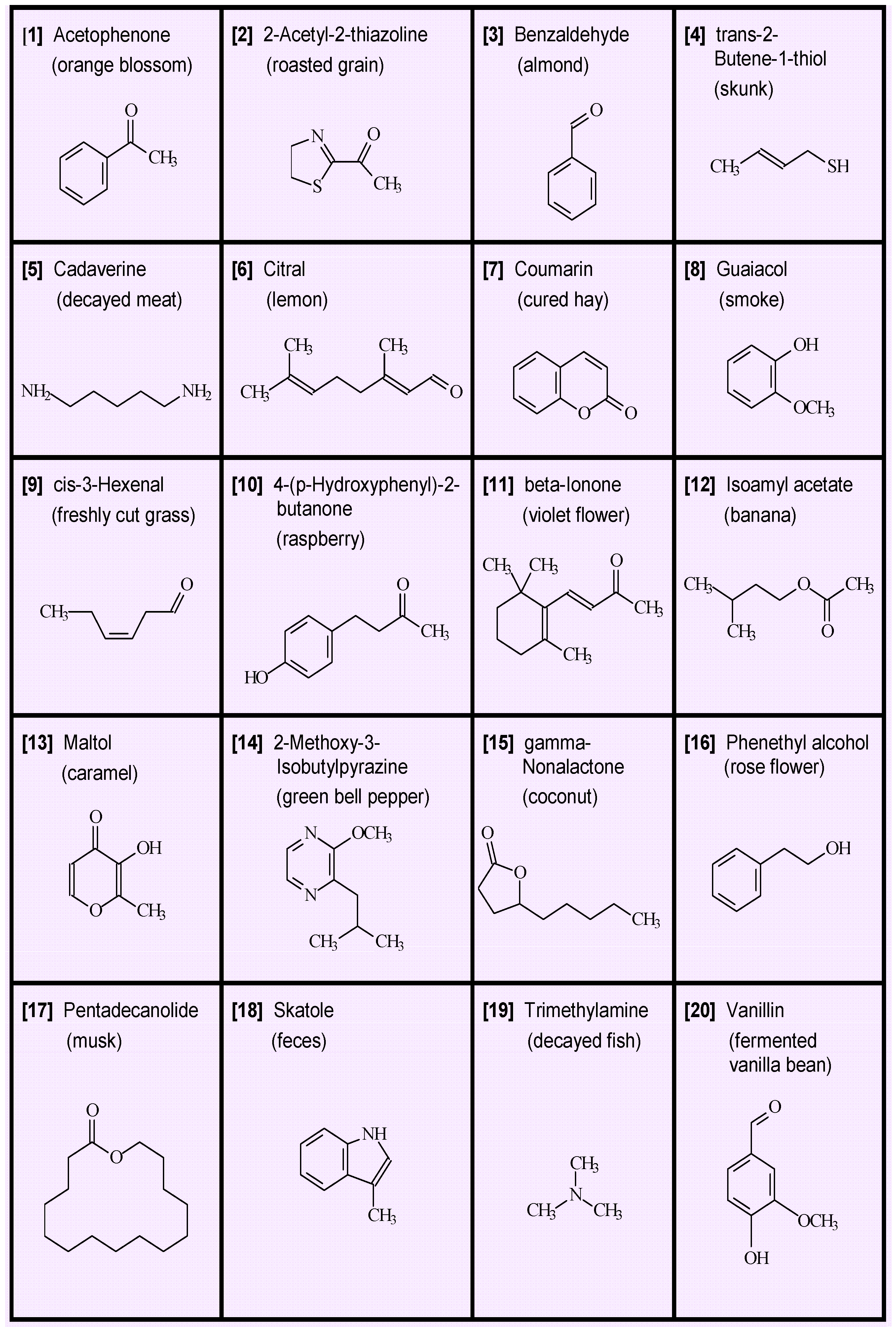
- Single odors. Perhaps the most straightforward aspect of smell is odor identification, which generally attaches a name or label to an odor object. There are, nevertheless, challenges in naming odors because odor objects are not always completely defined and because subjects may have different perceptions and experiences. A key observation that has been made numerous times in the history of odor chemistry is that many natural odors can be imitated by a single chemical substance. We present in Figure 1 an array of 20 diverse chemicals that by themselves epitomize a characteristic odor object [1]. For example, single compounds such as vanillin convey a pleasant food odor but a decayed odor is conveyed by cadaverine. We note that in the case of the latter compound, numerous metabolic decomposition products can be present, but odor of decay can be represented by a single chemical [2].
- Number of odorants in single odor objects. The natural environment is replete with numerous odors, but most of the time we are not aware of any odors. This is partly due to the effect of adaptation, but it is also due to the great dilution of odors compared to the concentration at their source. Even here, we should recognize that flowers are most often associated with the idea of odor, yet flowers are often completely odorless. If half of all flowers are odorless, we can speculate on the chemical composition of odorants in flowers that have odors. We would not expect nature to pack many odorants into some flowers and none in others. Suppose there were just eight possible odorants, and a given flower had an 8% chance of producing each odorant, then the probability that the flower would be odorless is 0.928, or about 51% by chance. In this assumed situation, about 49% of flowers would have an odor and binomial calculations show about 73% of them would have one odorant, about 22% would have two odorants, and about 4% would have three odorants. Similar results are obtained for a larger number of potential odorants and a correspondingly lower chance occurrence per odorant. The conclusion would be the odor of the majority of flowers is due to a single component, and only about 1% is due to more than three components. This argument can be extended to other sources of odor than flowers, but with less certainty.
- Few odors dominate perception. The sparse existence of odorants in the environment and in odor objects in general is a concept at odds with accepted views of olfaction. Many articles repeat the refrain that objects such as coffee or grilled meat contain hundreds of odorants that act in a combinatorial way to produce their characteristic odors. While it is true that many odorants are present, they are mostly present at concentrations below threshold and therefore contribute little or nothing to the overall odor. Odorants are present at widely different perceptual intensities, with the strongest odors dominating the perception. In the paper by Bushdid et al. [3], the calculation of the possibility of a trillion distinguishable odors was based in part on the use of artificial mixtures of as many as 30 components that were presumed to be equally intense. Such a situation is unrealistic and difficult to achieve. If there was an imbalance in intensity of the components, the calculated value could be too large by many orders of magnitude. In simple binary mixtures, the dominance of one component over the other is achieved when the intensities of the components differ by a factor of more than 1.4. An extensive discussion of the perception of binary mixtures is presented in Section 2.
1.3. Chemical Identification and Purity
1.4. Odor Intensity
- Stevens’ power law, subjective and no threshold. After chemical components of odor objects are properly identified, it is essential to determine the relative intensities or salience of the components. In the sensory psychology literature, the perceptual intensity of an odorant is generally estimated from Stevens’ power law, which states that intensity (I) is a power function of concentration (C) such that , where k is a constant and the exponent (a) can vary between about 0.1 and 1, depending on the odorant and conditions of measurement. An exponent below 1 is interpreted to mean that perceptual intensity is a ‘compressed’ function of concentration. In our treatment, we assume that the typical value of the exponent is approximately 0.5, that is, the odor intensity is proportional to the square root of the concentration (). A problem with the Stevens’ power law is the need for the subject to make what is essentially a subjective estimate of intensity by the method of magnitude estimation. Also, the power law does not provide an absolute reference point for intensity and does not establish a threshold value.
- Odor activity value, stimulus/threshold concentrations. A way around these problems is to establish a reasonably objective measurement of threshold and then to infer relative stimulus intensity from the ratio of suprathreshold concentration (C) to threshold concentration (T). In the food and fragrance literature, this ratio (C/T) is called the odor activity value (OAV) [4]. The OAV is not equal to perceptual intensity (I), but a mathematical relationship can be established as follows: Because and , then . Thus, the ratio of perceived odor intensity to its intensity at threshold is approximately equal to the square root of the odor activity value. This relationship will be used later in the treatment of perception of mixtures (Section 2) and development of stochastic models (Section 4).
1.5. Threshold Determination
1.6. Receptor Mechanisms
- Numbers of receptors. Chemosensory receptors for taste and smell have distinctly different chemistries (Table 1). Humans have about 390 types of functional olfactory receptors and only about 34 types of taste receptors, most of which serve to detect bitter stimuli.
- Combinatorial odor coding. In developing the combinatorial concept for receptor activation for odors, Malnic et al. [6] found that receptors appeared to be broadly tuned and that many odorants activated multiple receptors. This may be the case when relatively high concentrations of odorants in the range of 10 to 100 micromolar are used. Some stimuli having little volatility and no odor, such as dicarboxylic acids, were used in developing the combinatorial theory. At much lower concentrations that exist in the natural world, a more sparse activation of odorant receptors would be expected. Odorants are probably identifiable in many cases at concentrations of less than 1 micromole. Activation of a few receptors or even a single receptor might be responsible of the perception of characteristic odor notes. This point has been emphasized in Figure 1, where single odorants can often define odor objects. A single compound could also be functionally heterogeneous and have more than one odor note, but one note is likely to be dominant because of mixture suppression, discussed in Section 2.
- Odor antagonism in mixtures. A recently described model [7] of odor antagonism in the molecular receptors (ORs) or olfactory receptor neurons is hypothetical and rather questionable, as the main antagonism occurs in the olfactory bulb with stimuli that are orthogonal or distinctly different. OR antagonism has only been reported in a few cases [8,9] and would be expected mostly for similar odorants rather than different ones. Each OR has the potential to suppress all the other ORs represented in the olfactory bulb. The perception of odor quality is critically dependent on the balance of excitation and inhibition in neural circuits of the olfactory bulb [10,11]. We discuss the perception of mixtures in Section 2 and present a mathematical model for component identification in Section 4.
2. Perception of Mixtures
2.1. Preamble
2.2. Mixture Component Dominance
- Relative perceived intensity. Olsson [15] studied the important phenomenon of component dominance in binary mixtures. As the relative intensity of the components diverge, one or the other component quality dominates the perception. Olsson questioned the idea of specific odor “maskers”, and concluded that the ability of one odor to mask another was “just a matter of relative perceived intensity of the masker”. Various models of component interactions are discussed in Section 4.
- Mixture interactions. The mathematical assessment of mixture interactions in binary odor mixtures has been reviewed extensively by Ferreira [16]. Olsson [15] and Ferreira indicated that for the most part binary mixtures of diverse odors retain the odors of the components. In a range of mixtures when both components are present at similar perceptual intensities, they can both be identified. The odors are considered to be co-dominant in this range. For a typical case (and also for an average case), when the component intensities (when measured in isolation by psychophysical tests) differ by a factor of about 1.4, the dominant component is recognized about twice as often as the suppressed component.
- Odor activity values. In most cases in the literature and in our studies on mixtures, magnitude estimation was used to establish equal perceptual intensity of components. To obtain a more objective estimate of concentrations producing equal perceptual intensity of components in mixtures, odor activity values (OAVs) can be used. The OAV procedure derived from the food and fragrance literature [4] defines the OAV as the ratio of odorant concentration (C) to its threshold value (T). We discussed in Section 1 the development of the relation of perceptual intensity to OAV, which is represented by the equation . For binary mixtures, intensities are presumed to be equal if the concentrations of both components have the same OAV. Thus OAV1 = C1/T1 = OAV2 = C2/T2 when components are equally intense. The OAV is equal to 1 at threshold and is <1 at sub-threshold concentrations and >1 at supra-threshold concentrations. This is an easy way to directly match perceived intensity of components in binary mixtures. In order to relate the concentrations to perceived intensity when the intensities are not matched, we use the equations and and making the reasonable assumption that , we get ). Thus, the ratio of the perceived intensities of the two components is equal to the square root of the ratio of their odor activity values.
- Component dominance/co-dominance. Because the perceived intensity is a compressed function of concentration, related approximately to the square root of concentration, the transition from co-dominance to dominance (at a 2:1 ratio of detection probability in the mixture) occurs when the perceived intensities of the components in isolation differ by a factor of about 1.4 or when the OAV of the dominant component is twice as large as the OAV of the suppressed component.
- e.
- Probabilities determine component detection. When the intensity of components of binary odor mixtures are not matched, the more intense component tends to suppress or dominate the weaker component. Olsson [15] noted that the dominance is not simply proportional to the relative intensity of the components in the mixture, but is exaggerated. For example, in a study of butanol-pyridine mixtures, if the intensity of one of the components was 1.4 (about the square root of 2) times the other, the probability of detecting the stronger component was about twice the probability of detecting the weaker component. Olsson and Cain [17] provided a simple equation for this relationship: P(a) = RA2/(RA2 + RB2), where P(a) is the probability of detecting component A, RA is the relative intensity of component A in isolation and RB is the relative intensity of component B in isolation. Olsson and Cain [17] summarized their findings as follows: “The probability (P) that a substance A will dominate a mixture of A and B depends on the perceived intensity (R) such that P(a) = RA2/(RA2 + RB2). In other words, the olfactory system seems to accentuate the difference in intensity between two odors when determining a mixture quality”. This concept is discussed later in Section 4, where we present a general model of component interactions.
- f.
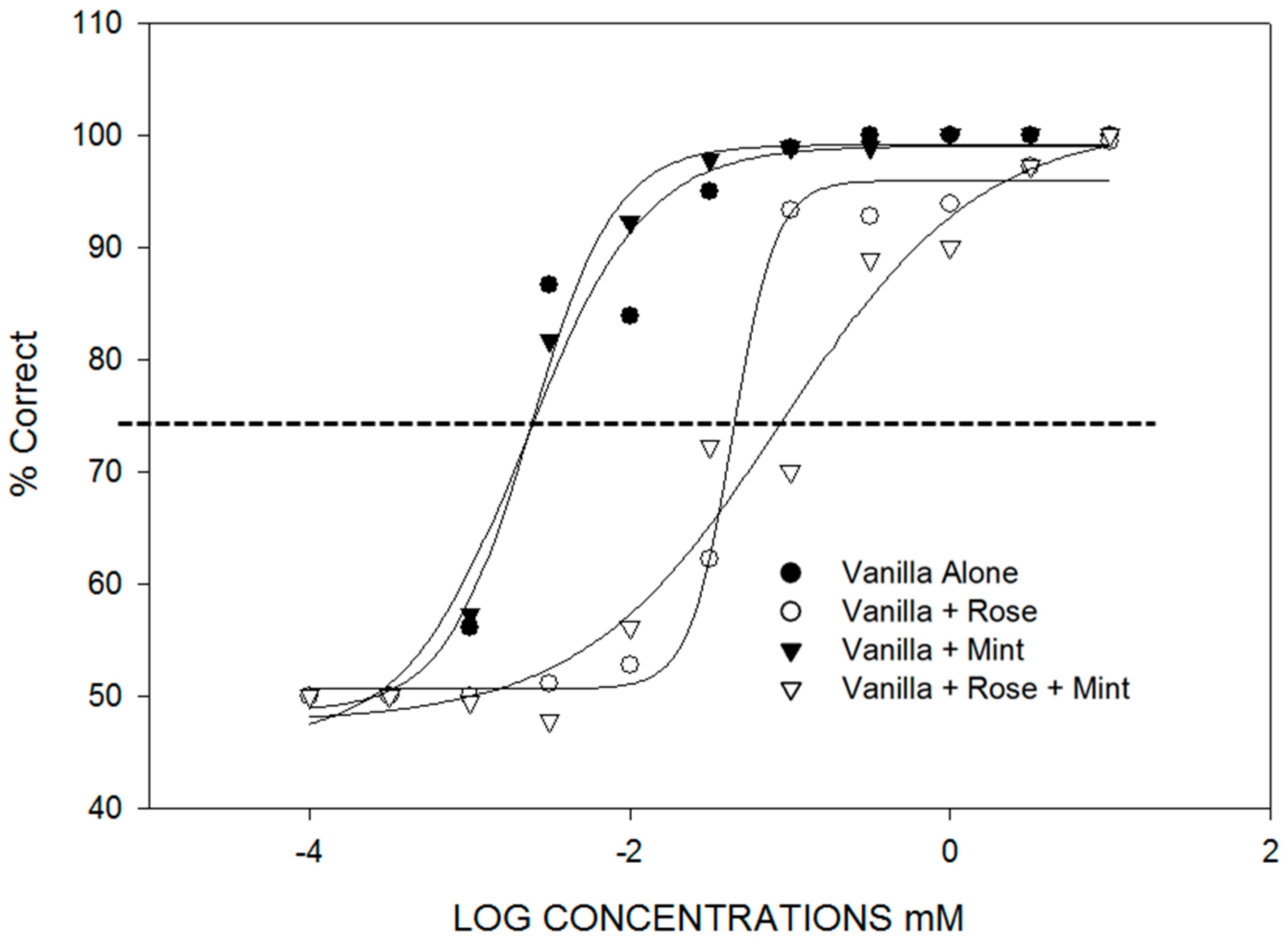
- Selective adaptation, background, and genetics affect component identification. There are three situations where identification of components in mixtures reflects the relative intensities of the components in unique ways. The first is the identification of components in mixtures following selective adaptation. This is presented in Section 3. The second case is the identification of a particular odorant in a background of another odorant (Figure 2). In [21], binary and ternary mixtures containing vanillin were found to have vanillin thresholds dependent on the background odorant concentration. For example, vanillin thresholds were raised an order of magnitude in the presence of a background containing phenethyl alcohol. The third case is the identification of odor components in a mixture where genetic differences determine the perception of the components and the mixtures (Figure 3). In [22], individual differences were found in the perception of a mixture of vanillin and beta-ionone that depended on whether the subject had a low or high threshold for beta-ionone, for which a genetic basis is established [23]. “Smellers” and “non-smellers” of beta-ionone differed significantly in their beta-ionone thresholds, but not in their vanillin thresholds. Because of component dominance, “smellers” tended to perceive a mixture of vanillin and beta-ionone to be “violet” and “non-smellers” tended to perceive the same mixture as “vanilla” (Table 2).
3. Selective Adaptation
3.1. Preamble
3.2. Selective Adaptation Defined
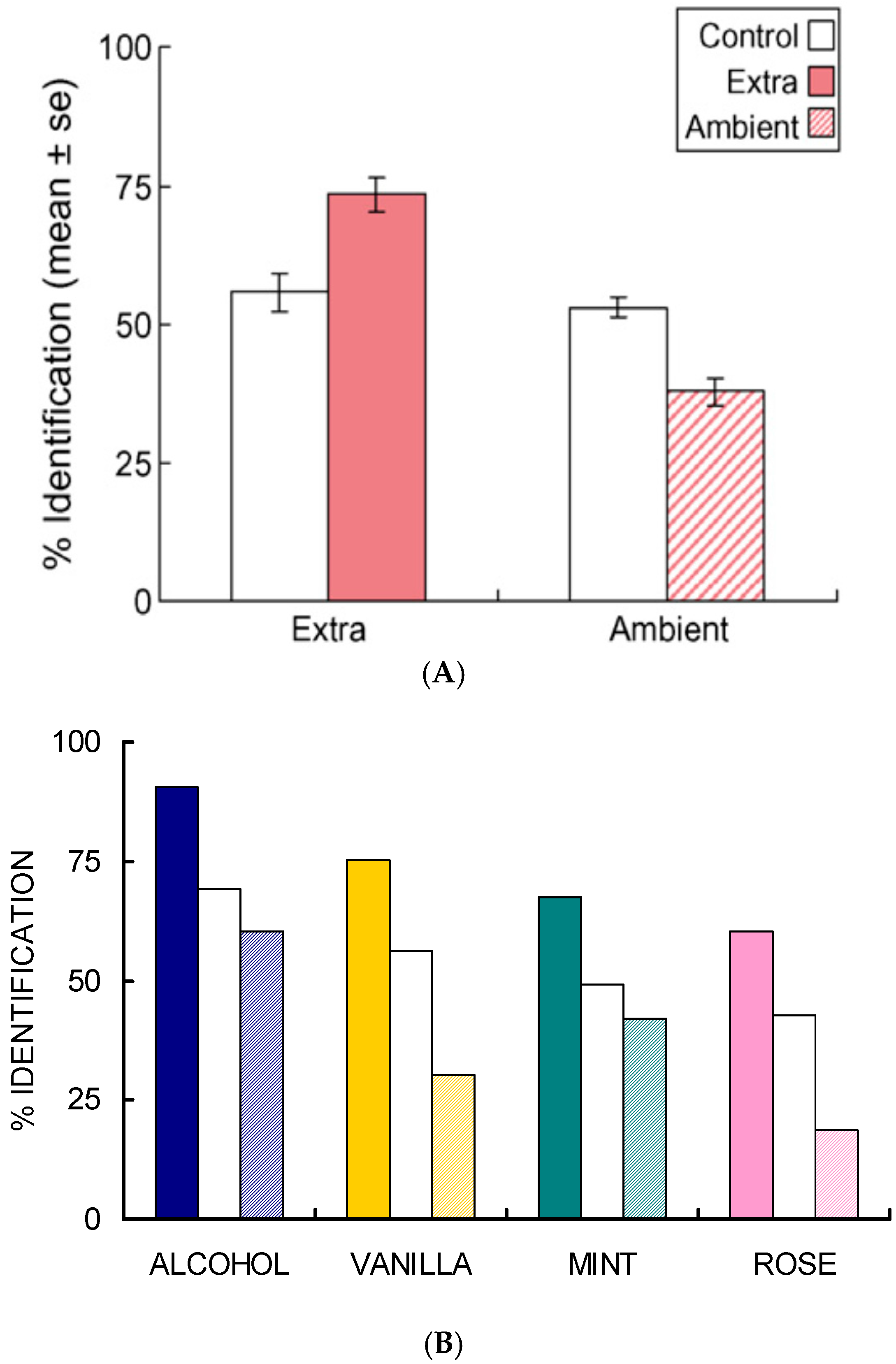
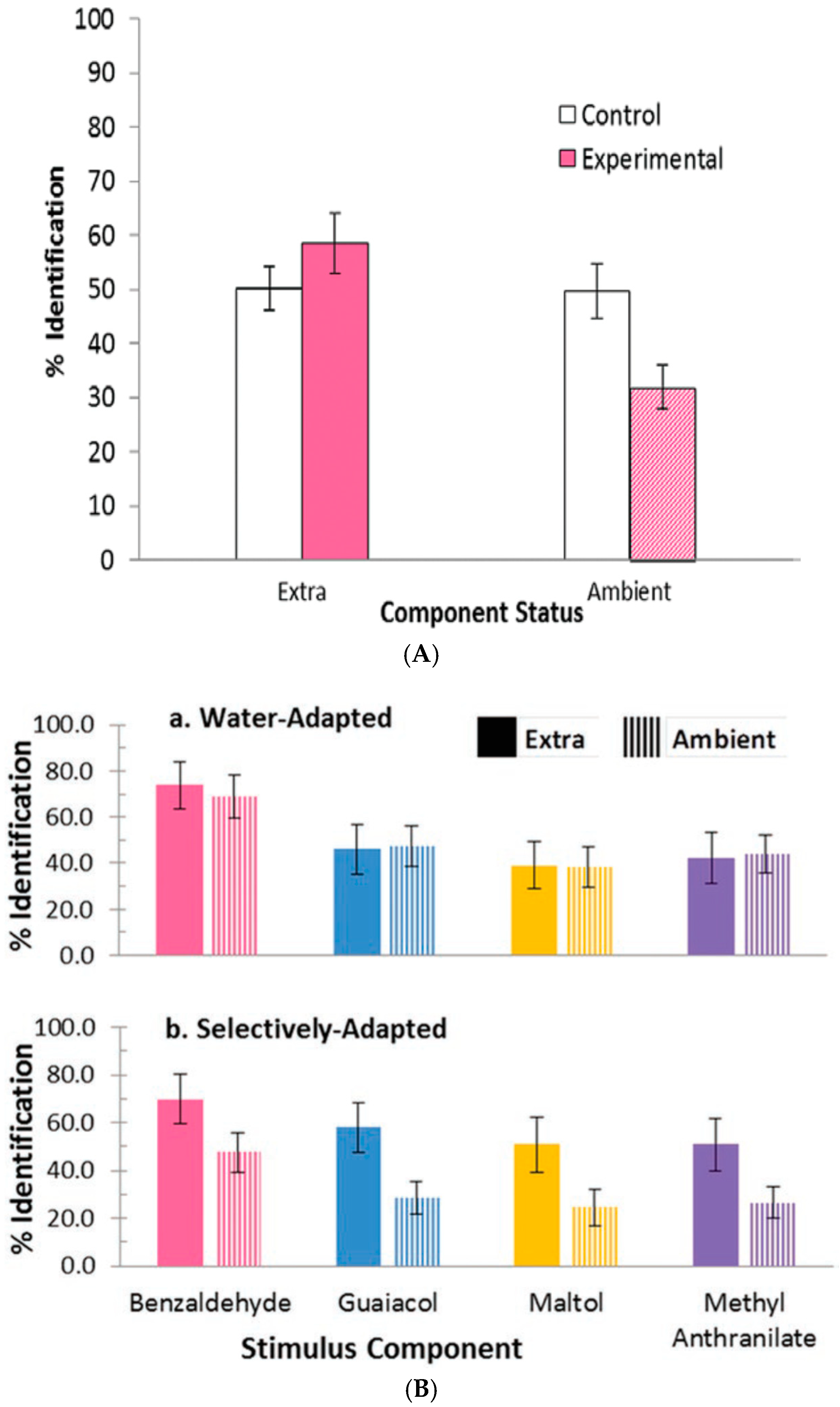
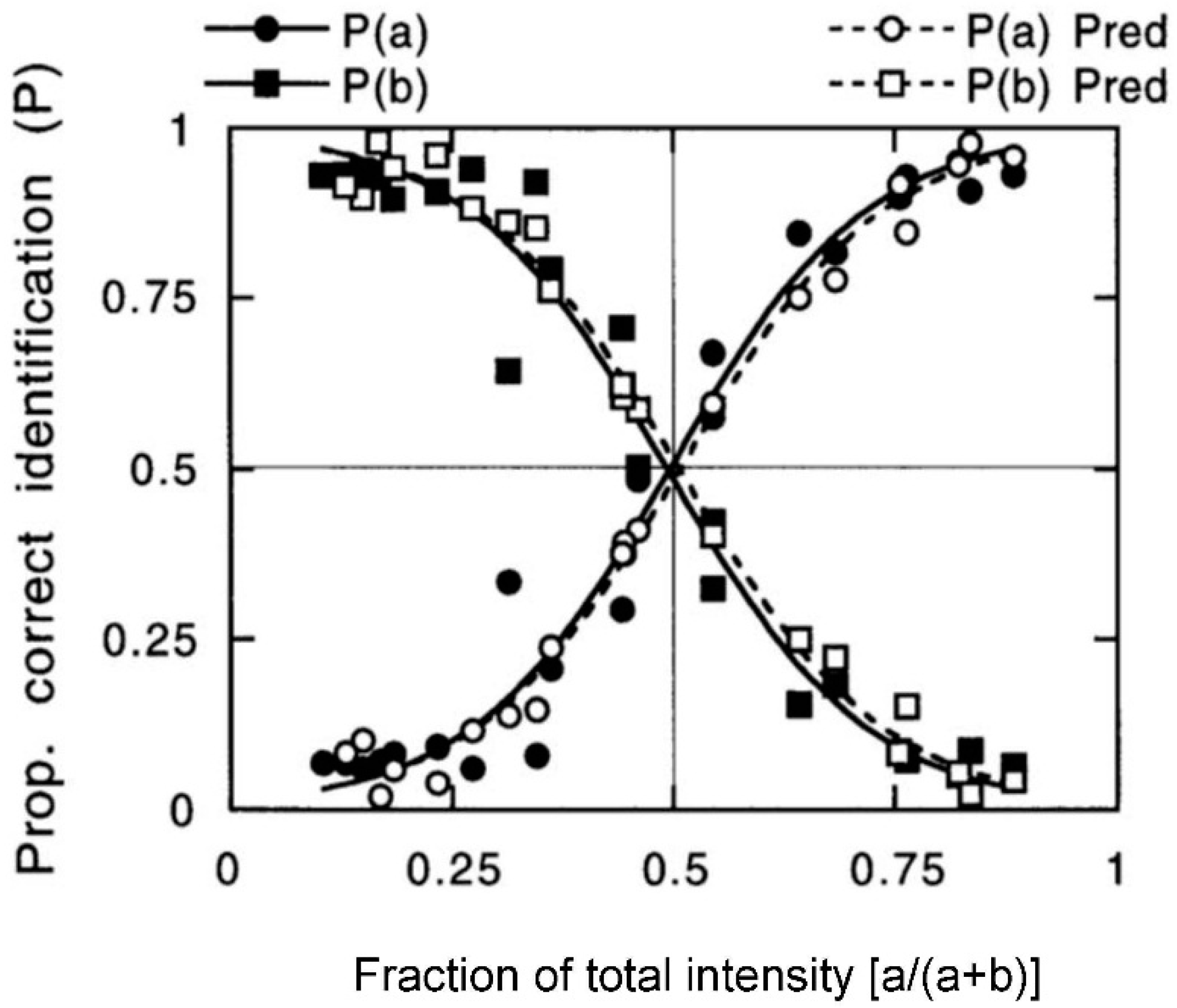
- Studies of selective adaptation. In our first study [26,27], we used mixtures containing (quality names in parentheses) isopropyl alcohol (alcohol), vanillin (vanilla), l-carvone (mint), and phenethyl alcohol (rose) to determine the effect of selective adaptation (Figure 4). In our second study [28], we used mixtures of benzaldehyde (cherry, almond), guaiacol (smoke), maltol (caramel, cotton candy), and methyl anthranilate (grape) (Figure 5). These two studies gave consistent results for the different four-component mixtures and showed improved ‘extra’ component identification as a result of selective adaptation of ‘ambient’ components.
- Salient mixture components retain qualities. These different odor sets [26,27,28] suggest that equally salient components retain, to a certain extent, the qualities of the mixture components. In the case of selective adaptation (Figure 4 and Figure 5), odor intensities of the components become unbalanced. Then it is possible to estimate the relative effective concentrations of the ambient and extra components in the mixture. After selective adaptation for about 4 s, identification of the extra component is about twice as likely as identification of the adapted component. The extra component is considered to be dominant. We thus conclude that the process of selective adaptation effectively reduces the apparent concentration of the adapted component to a level about one-half the actual value in the mixture.
- Adaptation half-life. We would then estimate (assuming first-order kinetics) the half-life (t1/2) for adaptation (in concentration terms) of about 4 s. Using the half-life equation for first-order reactions (k = 0.693/t1/2) we calculate the rate constant for adaptation to be about 0.17 s−1. Of course these are very approximate, but it is important to produce an estimate to compare with known values for adaptation at various levels of the nervous system.
- Selective taste adaptation. The phenomenon of selective adaptation also occurs in the gustatory system. Our results [29] for taste mixtures of sucrose and sodium chloride showed that extra component identification is improved by selective adaptation to ambient components.
4. Models of Component Interactions in Binary Odor Mixtures
4.1. Preamble
4.2. Probabilities of Component Identification
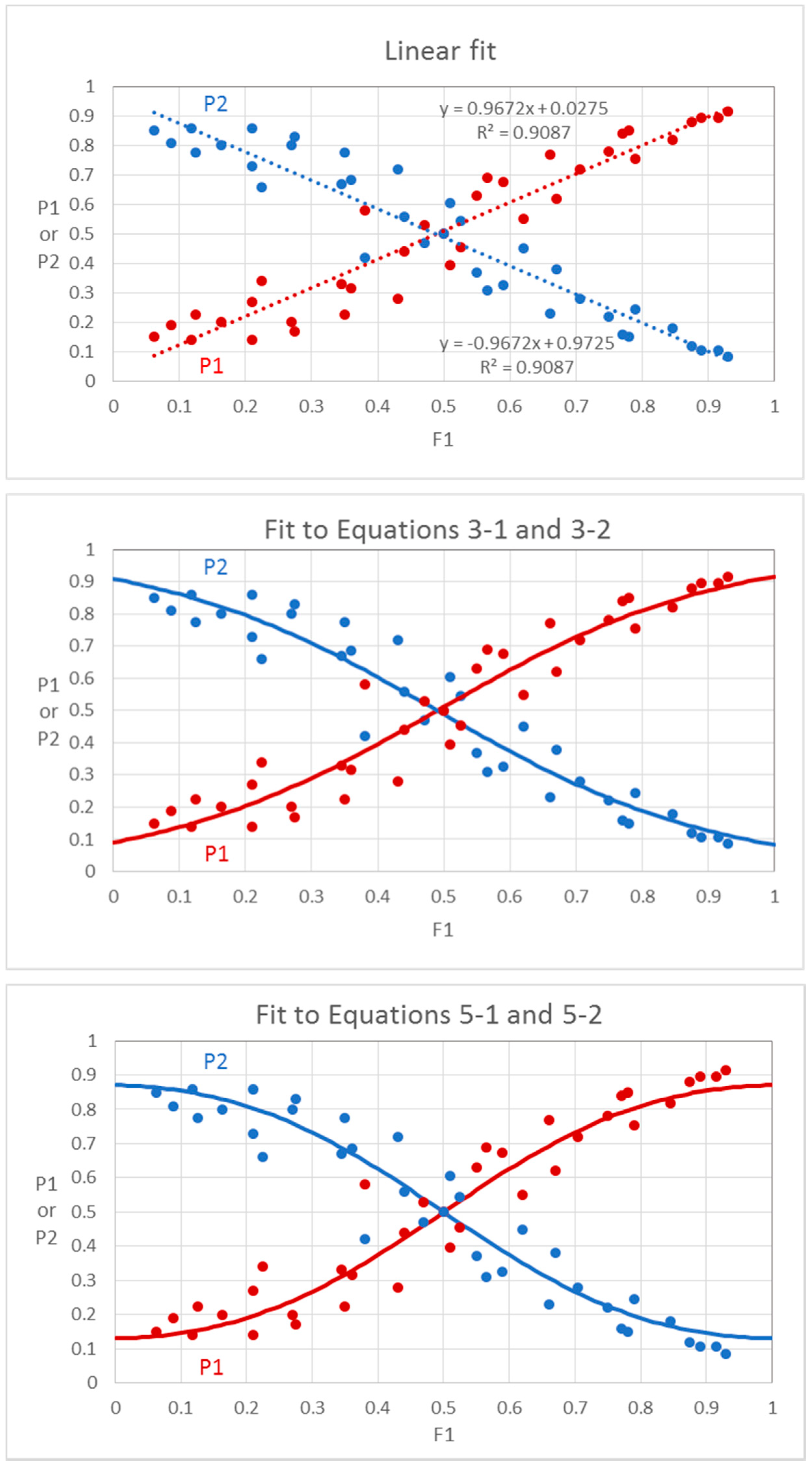
- Sigmoid/logistic functions. It is notable that a plot of P1 or P2 versus F1 using Equations (2-1) and (2-2) produces sigmoid functions within the defined limits of 0 to 1 for F1. These plots are similar to logistic plots of Ferreira [16], but without logarithmic or exponential operations. The squared intensity terms provide an exaggerated or relatively steep relationship between P1 and F1 in the central region of the plot compared to a linear function. The slope of the equation at the inflection point (0.5, 0.5) is 2, the value of the F1 exponent. It is possible that the exponent could be different for different mixtures, but we consider a value of 2 to be typical for odorants that are orthogonal and have non-overlapping odors. This typical case is shown in Figure 7.
- b.
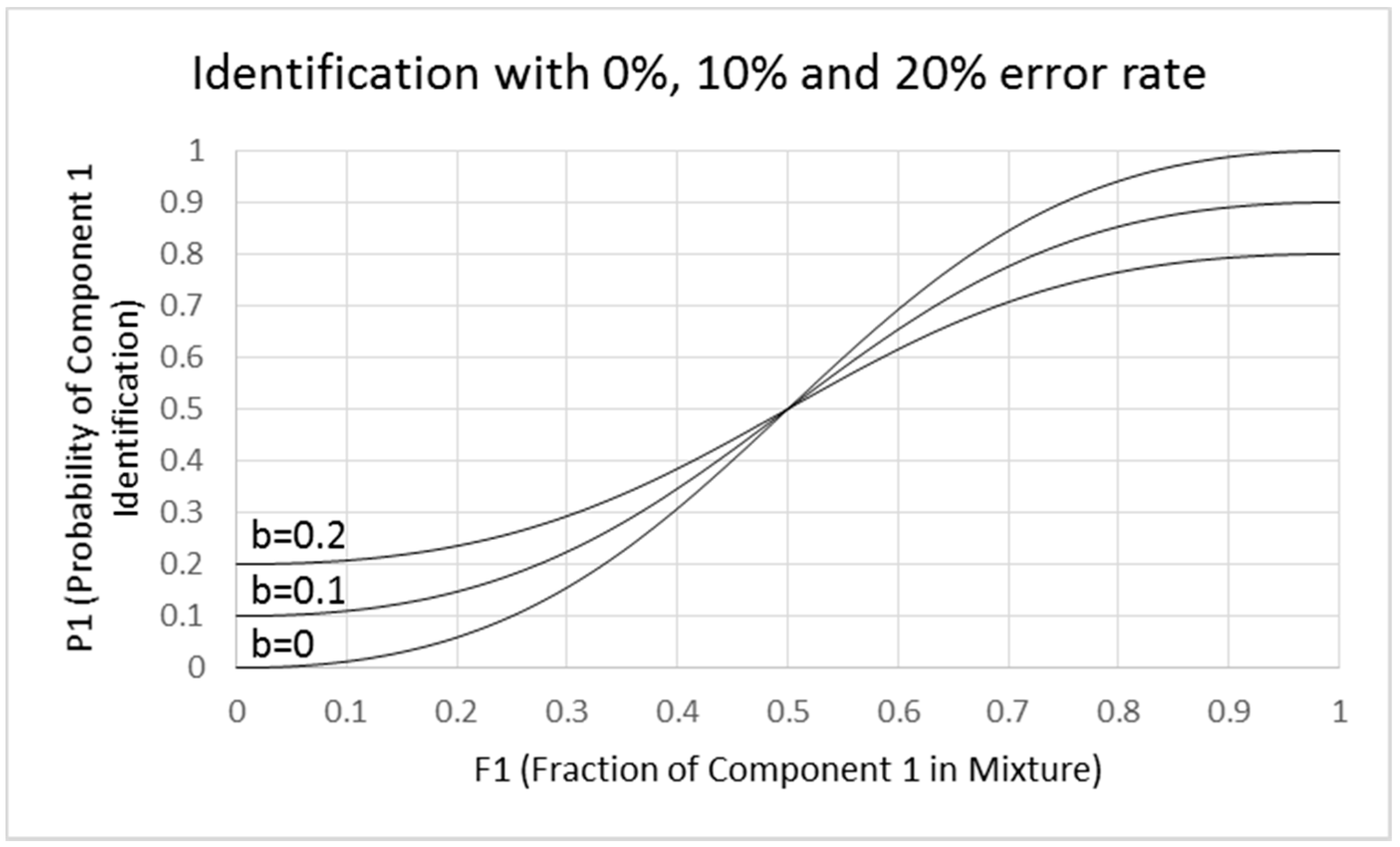
- Misidentifications. Though Ferreira did not consider identification errors in explaining the logistic fits, it is likely that the flat functions are due to misidentifications. We show in Figure 8 examples of how this could occur. We made pragmatic adjustments of Equations (2-1) and (2-2) to develop Equations (4-1) and (4-2), which show increasing flattening of the sigmoid function with increasing error rate defined by intercept b. The slope at the inflection point is 2(1 − 2b). When b = 0, the curve is identical to that shown in Figure 8 with a slope of 2. When b = 0.1, the slope is reduced to 1.6, and when b = 0.2, the slope is 1.2. This result suggests that the flattening of the function can be due to identification errors rather than a change in the exponent of the fractional component intensity. It also indicates that analysis of mixtures with extensively overlapping odor qualities would be difficult. Analysis of mixtures works best when odors are orthogonal and non-overlapping. An exponent of 2 for the fractional component intensities in binary mixtures may be a general property for orthogonal odorants.
- c.
- Exponent effects. Finally, we also consider equations where the exponent of F1 may not be 2 but could be any positive value = n (Equations (5-1) and (5-2)). The value of n is analogous to the Hill coefficient in enzyme kinetics where it is related to cooperativity among subunits. The value of n has no clear meaning for dominance in odor mixtures, but it may be speculated that values of n greater than 1 represent convergent neural activity in the CNS. Because Equations (5-1) and (5-2) cannot be linearized, the b and n parameters are determined by successive approximation and minimizing the sum of squared deviations of P1 from the fitted line. We provide an example in Supplementary Material of how data can be fit to Equations (5-1) and (5-2) in an Excel program.
- d.
- One or two sniffs can reduce ambient component intensity by a factor of 1.4. In our studies [26,27,28] on selective adaptation (Figure 4 and Figure 5), we observed that, for two different datasets each involving four components in various mixture combinations, extra components were on average identified twice as often as ambient components which had been partially adapted over a few seconds. This suggests that the intensity of the ambient components had been reduced by a factor of about 1.4 during one or two sniffs. Of the eight different odorants used in the two studies, the individual components were identified with an accuracy of about 90%, except for methyl anthranilate, which was correctly identified only about 60% of the time. It is difficult to generate mixtures of four or more components where each component has an odor completely distinct from the others.
- e.
- Detecting odor mixture interactions. Models of odor interactions in mixtures are dependent on how odor intensity is measured and how component intensities are matched. In the sensory psychology literature, perceived odor intensity is considered to be a power function of odorant concentration (Stevens’ power law). We assumed a typical value for the concentration exponent to be about 0.5 according to the equation I = kC0.5, where I is the perceived intensity, C is the odorant concentration and k is a constant. When comparing different odorants with different and unknown values of k, odor intensities are typically determined by magnitude estimation, and matched by direct comparison of perceived odor intensities or by cross-modality matching. This is a difficult procedure that involves subjective judgments that are prone to bias and other errors. As noted in Section 2, in an attempt to produce a more objective measure of odor intensity, flavor, and fragrance studies use the concept of odor activity value (OAV), which is the ratio of odorant concentration to its threshold value [4]. This method has been criticized because in comparing different odorants in mixtures, OAVs were not considered to be additive in the same way as odor intensities. However, as shown in Section 2 for a binary mixture, we were able to eliminate k from the paired power law equations and generate a new equation for the relationship between perceived intensity and OAV for mixture components: ). This means that the ratio of odor intensities is (approximately) equal to the square root of the ratio of their respective OAV values.
- f.
- Analytic odor perception. Our treatment is explicitly analytical and does not consider odor mixtures to have synthetic properties. The treatments of Olsson [15] and Ferreira [16] are also analytical, while there are many researchers who consider olfaction to be synthetic or combinatorial like color vision. Thomas-Danguin et al. [18] and Wilson [19] have argued for configural or synthetic processing of odor mixtures. We do not disagree with their approach, but we think there are strong arguments for treating odor mixtures as resolvable into components. Combinatorial arguments based on receptor activation [6] for pure compounds and mixtures use the relatively broad tuning of receptors to imply that odor perception is essentially synthetic. However, activation of broadly tuned receptors could be neutralized or suppressed by activation of other more efficacious and selective receptors. In this way, the combinatorial process is more a process of subtraction than addition, with the more intense components dominating the perception.
- g.
- A unique or synthesized perception? A combinatorial process was also invoked for binary odor mixtures in mice [20]. Though not explicitly stated, the combinatorial model involves a unique or synthetic perception for binary mixtures. Olsson [15], Olsson and Cain [17], and Ferreira [16] have discussed typical situations where binary odor mixtures are perceived as two distinct components, and we have argued that both components are often perceived in binary mixtures if the components are present at equal intensities. When the components are not balanced in intensity, generally one component will dominate the perception. Even when perceptual intensities are matched and both components are recognized, it is likely that the individual components are identified in a probabilistic manner, so both components are not recognized exactly at the same time. If we were to propose that both components could be recognized simultaneously, this would amount to a synthetic composition, in which neither component could actually be recognized, negating the original premise. As in the case of color vision, red and green cannot be simultaneously recognized.
5. Summary and Conclusions
Supplementary Materials
Acknowledgments
Conflicts of Interest
References
- Frank, M.E. Chapter 10, Chemoreception and Perception. In Fundamentals of Oral Histology and Physiology; Hand, A.R., Frank, M.E., Eds.; Wiley and Sons: Hoboken, NJ, USA, 2014; pp. 191–217. [Google Scholar]
- Dieris, M.; Ahuja, G.; Krishna, V.; Korsching, S.I. A single identified glomerulus in the zebrafish olfactory bulb carries the high-affinity response to death-associated odor cadaverine. Sci Rep. 2017, 7, 40892. [Google Scholar] [CrossRef] [PubMed]
- Bushdid, C.; Magnasco, M.O.; Vosshall, L.B.; Keller, A. Humans can discriminate more than 1 trillion olfactory stimuli. Science 2014, 343, 1370–1372. [Google Scholar] [CrossRef] [PubMed]
- Grosch, W. Determination of potent odourants in foods by aroma extract dilution analysis (AEDA) and calculation of odour activity values (OAVs). Flavour Fragr. J. 1994, 9, 147–158. [Google Scholar] [CrossRef]
- Cometto-Muñiz, J.E.; Abraham, M.H. Dose-Response Functions for the Olfactory, Nasal Trigeminal, and Ocular Trigeminal Detectability of Airborne Chemicals by Humans. Chem. Senses 2016, 41, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Malnic, B.; Hirono, J.; Sato, T.; Buck, L.B. Combinatorial receptor codes for odors. Cell 1999, 96, 713–723. [Google Scholar] [CrossRef]
- Reddy, G.; Zak, J.D.; Vergassola, M.; Murthy, V.N. Antagonism in olfactory receptor neurons and its implications for the perception of odor mixtures. eLife 2018, 7, e34958. [Google Scholar] [CrossRef] [PubMed]
- Oka, Y.; Nakamura, A.; Watanabe, H.; Touhara, K. An odorant derivative as an antagonist for an olfactory receptor. Chem. Senses 2004, 29, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Brodin, M.; Laska, M.; Olsson, M.J. Odor interaction between Bourgeonal and its antagonist undecanal. Chem. Senses 2009, 34, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Migliore, M.; Hines, M.L.; Shepherd, G.M. Sparse coding and lateral inhibition arising from balanced and unbalanced dendrodendritic excitation and inhibition. J. Neurosci. 2014, 34, 13701–13713. [Google Scholar] [CrossRef] [PubMed]
- Storace, D.A.; Cohen, L.B. Measuring the olfactory bulb input-output transformation reveals a contribution to the perception of odorant concentration invariance. Nat. Commun. 2017, 8, 81. [Google Scholar] [CrossRef] [PubMed]
- Laing, D.G. Natural sniffing gives optimum odour perception for humans. Perception 1983, 12, 99–117. [Google Scholar] [CrossRef] [PubMed]
- Livermore, A.; Laing, D.G. Influence of training and experience on the perception of multicomponent odor mixtures. J. Exp. Psychol. Hum. Percept. Perform. 1996, 65, 267–277. [Google Scholar] [CrossRef]
- Livermore, A.; Laing, D.G. The influence of odor type on the discrimination and identification of odorants in multicomponent odor mixtures. Physiol. Behav. 1998, 65, 311–320. [Google Scholar] [CrossRef]
- Olsson, M.J. An integrated model of intensity and quality of odor mixtures. Ann. N. Y. Acad. Sci. 1998, 30, 837–840. [Google Scholar] [CrossRef]
- Ferreira, V. Revisiting psychophysical work on the quantitative and qualitative odour properties of simple odour mixtures: A flavour chemistry view. Part 2, qualitative aspects. A review. Flavour Fragr. J. 2012, 27, 201–215. [Google Scholar] [CrossRef]
- Olsson, M.J.; Cain, W.S. Psychometrics of odor quality discrimination: Method for threshold determination. Chem. Senses 2000, 25, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Thomas-Danguin, T.; Sinding, C.; Romagny, S.; El Mountassir, F.; Atanasova, B.; Le Berre, E.; Le Bon, A.M.; Coureaud, G. The perception of odor objects in everyday life: A review on the processing of odor mixtures. Front. Psychol. 2014, 5, 504. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.A. Pattern separation and completion in olfaction. Ann. N. Y. Acad. Sci. 2009, 1170, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, L.R.; Kondoh, K.; Ye, X.; Yoon, K.H.; Hernandez, M.; Buck, L.B. Combinatorial effects of odorants on mouse behavior. Proc. Natl. Acad. Sci. USA 2016, 113, E3300–E3306. [Google Scholar] [CrossRef] [PubMed]
- Phares, A.N.; Frank, M.E.; Hettinger, T.P. Effects of background stimuli on odor detection thresholds. In Proceedings of the AChemS Meeting, St. Pete Beach, FL, USA, 13–17 April 2011; UCONN Health, Dental Medicine: Farmington, CT, USA, 2011. [Google Scholar]
- Dulieu, L.K.; Frank, M.E.; Hettinger, T.P. Discovering Qualities of Single-Stimuli Mixtures Having Double Odors; UCONN Health, Dental Medicine: Farmington, CT, USA, 2015. [Google Scholar]
- McRae, J.F.; Jaeger, S.R.; Bava, C.M.; Beresford, M.K.; Hunter, D.; Jia, Y.; Chheang, S.L.; Jin, D.; Peng, M.; Gamble, J.C.; et al. Identification of regions associated with variation in sensitivity to food-related odors in the human genome. Curr. Biol. 2013, 23, 1596–1600. [Google Scholar] [CrossRef] [PubMed]
- Zufall, F.; Leinders-Zufall, T. The cellular and molecular basis of odor adaptation. Chem. Senses 2000, 25, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hertz, J. Odour recognition and segmentation by a model olfactory bulb and cortex. Netw. Comput. Syst. 2000, 11, 83–102. [Google Scholar] [CrossRef]
- Goyert, H.F.; Frank, M.E.; Gent, J.F.; Hettinger, T.P. Characteristic component odors emerge from mixtures after selective adaptation. Brain Res. Bull. 2007, 72, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.E.; Goyert, H.F.; Hettinger, T.P. Time and intensity factors in identification of components of odor mixtures. Chem. Senses 2010, 35, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.E.; Fletcher, D.B.; Hettinger, T.P. Recognition of the Component Odors in Mixtures. Chem. Senses 2017, 42, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.E.; Goyert, H.F.; Formaker, B.K.; Hettinger, T.P. Effects of selective adaptation on coding sugar and salt tastes in mixtures. Chem. Senses 2012, 37, 701–770. [Google Scholar] [CrossRef] [PubMed]
- Poivet, E.; Tahirova, N.; Peterlin, Z.; Xu, L.; Zou, D.J.; Acree, T.; Firestein, S. Functional odor classification through a medicinal chemistry approach. Sci. Adv. 2018, 4, eaao6086. [Google Scholar] [CrossRef] [PubMed]
- Doszczak, L.; Kraft, P.; Weber, H.P.; Bertermann, R.; Triller, A.; Hatt, H.; Tacke, R. Prediction of perception: Probing the hOR17-4 olfactory receptor model with silicon analogues of bourgeonal and lilial. Angew. Chem. Int. Ed. Engl. 2007, 46, 3367–3371. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.E.; Formaker, B.K.; Hettinger, T.P. Taste responses to mixtures: Analytic processing of quality. Behav. Neurosci. 2003, 117, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.E.; Lundy, R.F., Jr.; Contreras, R.J. Cracking taste codes by tapping into sensory neuron impulse traffic. Prog. Neurobiol. 2008, 86, 245–263. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Variants | ||||
|---|---|---|---|---|
| Receptors | Receptor Cells | Sensory Neurons | ||
| Total Olfactory Receptors (OR) | 390 | 390 | 390 | |
| Total Taste Receptors (TR) | 34 | 5 | 5 | |
| Taste Receptor (TR) | T1R dimer | 2 | 2 | 2 |
| T2R | 30 | 1 | 1 | |
| ENaC | 1 | 1 | 1 | |
| PKD dimer | 1 | 1 | 1 | |
| Olfactory and Taste | 424 | 395 | ||
| β-Ionone ppb | ‘Smellers’ (n = 12) | ‘Non-Smellers’ (n = 4) |
|---|---|---|
| (1) 10 ppb | Identifications | |
| Violet only | 1 | 0 |
| Vanilla only | 12 | 4 |
| Both | 3 | 0 |
| (2) 100 ppb | ||
| Violet only | 8 | 0 |
| Vanilla only | 7 | 4 |
| Both | 8 | 0 |
| (3) 1000 ppb | ||
| Violet only | 12 | 3 |
| Vanilla only | 0 | 1 |
| Both | 4 | 4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hettinger, T.P.; Frank, M.E. Stochastic and Temporal Models of Olfactory Perception. Chemosensors 2018, 6, 44. https://doi.org/10.3390/chemosensors6040044
Hettinger TP, Frank ME. Stochastic and Temporal Models of Olfactory Perception. Chemosensors. 2018; 6(4):44. https://doi.org/10.3390/chemosensors6040044
Chicago/Turabian StyleHettinger, Thomas P., and Marion E. Frank. 2018. "Stochastic and Temporal Models of Olfactory Perception" Chemosensors 6, no. 4: 44. https://doi.org/10.3390/chemosensors6040044
APA StyleHettinger, T. P., & Frank, M. E. (2018). Stochastic and Temporal Models of Olfactory Perception. Chemosensors, 6(4), 44. https://doi.org/10.3390/chemosensors6040044