
Demographic Dynamics in Multitype Populations with Migrations


{kind=link}
{kind=link}
Abstract
1. Introduction
2. Model Description
- 1.
- Let , ; ; , be independent random vectors, with representing the number of type-l individuals produced by the ith type-k individual living at population j in generation n. For fixed k, we also assume that:are identically distributed with probability generating function (pgf),Thus, individuals of the same type reproduce according to a common offspring probability distribution, independently of the generation and the population where they live.
- 2.
- Consider the random vector , with denoting the number of type-l individuals born at population j in generation n. Hence,Note that, conditioned on , requirements on , ; ; , guarantee that , , are independent and identically distributed with pgf given by:
- 3.
- Let us introduce the random vectors , where the variable is the number of individuals that emigrate to population l, among the type-k individuals born at population j in generation n. Clearly,
- 4.
- Given , denote by the probability for an individual born at population j to emigrate to population h ( will represent the probability that an individual born at population j stays at such a population). Notice that , .We will assume that , conditioned upon , is distributed according to a multinomial probability law with parameters . It is assumed also that, conditioned on , the vector , , are independent vectors.On the other hand, since emigration events take place independently in each population, if is the matrix with columns the vectors then, conditioned on , it is derived that are independent random matrices. In particular, for fixed k, conditioned on , are independent random vectors.
- 5.
- Finally, in generation ,
3. Results
3.1. Properties
- is said to be irreducible if, for every , there exists such that . Otherwise, it is said that is reducible.
- is said to be positively regular if there exists such that , for all , i.e., is irreducible and n does not depend on i and j.
- (a)
- If either or are reducible matrices then the MBP is also reducible.
- (b)
- is positively regular if and only if matrices and are positively regular.
- Assume that is reducible. Then, for some , for all n. Hence, for all n whatever be. Consequently, is also reducible. We proceed analogously if is reducible.
- Let us assume that is positively regular. Then, there exists n such that , for all ; . Using again that , we have that , for all , and , for all . Thus, and are positively regular.Reciprocally, assume and positively regular. Therefore, there exists and such that all elements of and of are positive. Since:it is determined that all elements of are positive. Hence, is a positively regular MBP.
3.2. Limiting Results
- becomes extinct if and only if .
- If andthen, there exists a non-negative non-degenerate random variable W such that:where denotes the -marginal distribution function obtained from the distribution function associated to .
- (a)
- There exists a non-negative non-degenerate random variable W such thatwith . Therefore,
- (b)
- There exists a non-negative non-degenerate random variable such that
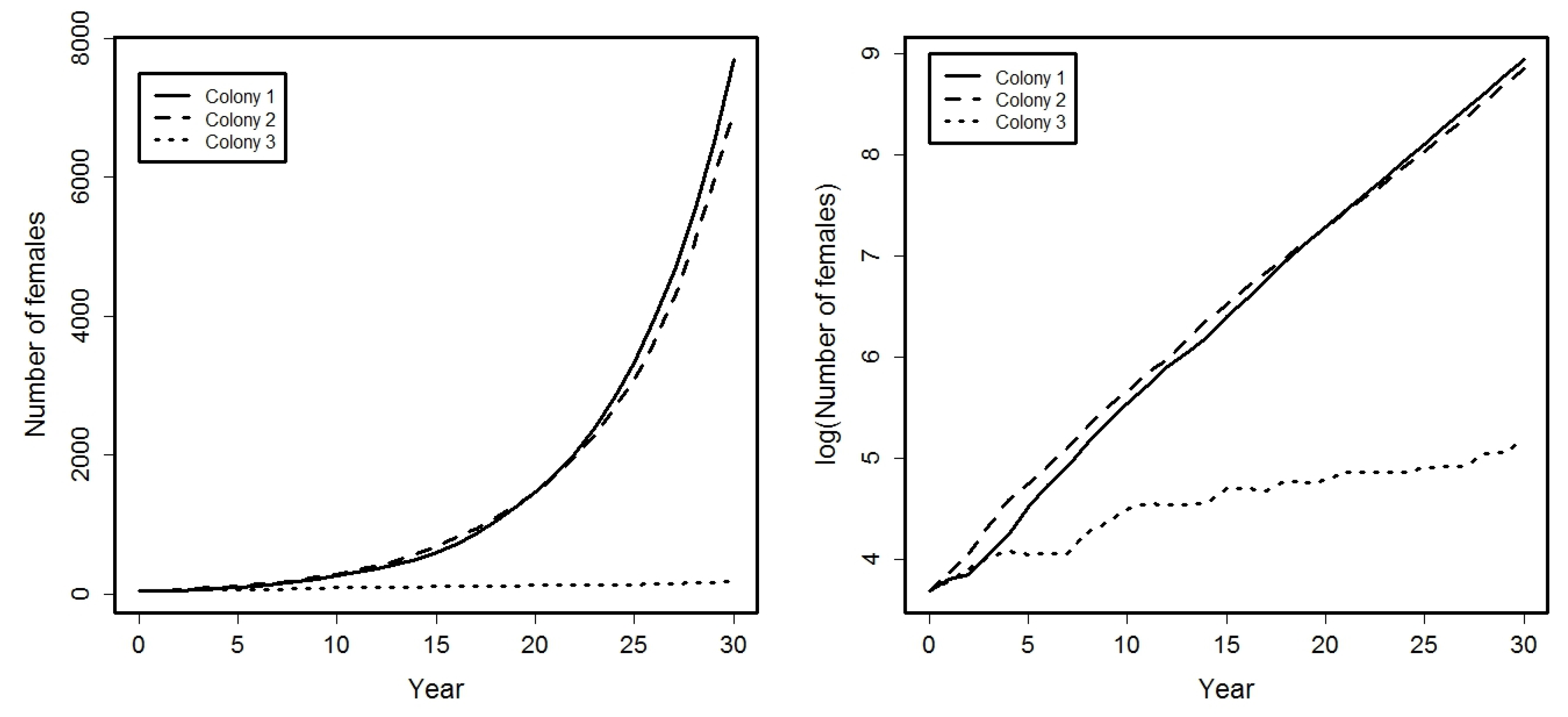
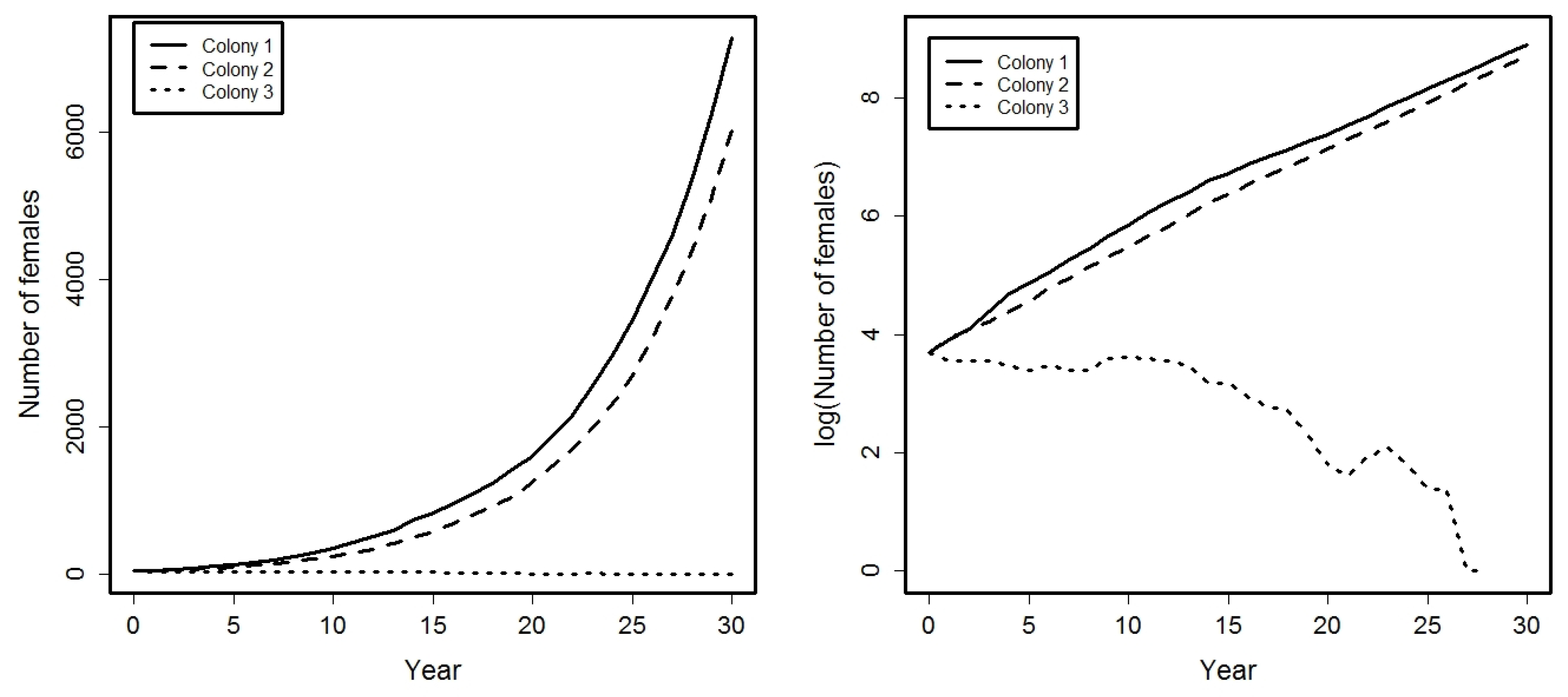
4. Application
- •
- The monogamous behavior throughout their life span.
- •
- The tendency to return to their birthplace or proximity in order to reproduce.
- •
- The common reproductive behavior. In fact, they have relatively stable populations and tend to produce a low numbers of offspring. The probability of breeding success depends on the age of the female.
- •
- The viability of their populations depends on the survival rate of the birds. Really, mortality rate in these raptor species during their reproductive life is very low.
- Success probabilities at different ages for the females are: , , , .
- A small percentage of females from colonies 1 and 2 emigrate among them: .
- Females from colony 3 can emigrate to colonies 1 or 2, being:
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Margalida, A.; Colomer, M.A.; Oro, D.; Arlettaz, R.; Donázar, J.A. Assesing the impact of removal scenarios on population viability of a threatened long-lived avian scavenger. Sci. Rep. 2015, 5, 16962. [Google Scholar] [CrossRef] [PubMed]
- Tauler, H.; Real, J.; Hernández-Matías, A.; Aymerich, P.; Baucells, J.; Martorell, C.; Santandreu, J. Identifying key demographic parameters for the viability of a growing population of the endangered Egyptian Vulture Neophron Pernopterus. Bird Conserv. Int. 2015, 25, 246–439. [Google Scholar] [CrossRef]
- Faddy, M.J. Stochastic compartmental models as approximations to more general stochastic systems with the general stochastic epidemic as an example. Adv. Appl. Prob. 1977, 9, 448–461. [Google Scholar] [CrossRef]
- Matis, J.H.; Wehrly, T.E. Stochastic models of compartmental systems. Biometrics 1979, 35, 199–220. [Google Scholar] [CrossRef]
- Asmussen, S.; Hering, H. Branching Processes; Birkhauser Boston, Inc.: Boston, MA, USA, 1983. [Google Scholar]
- Athreya, K.B.; Ney, P.E. Branching Processes; Springer: New York, NY, USA, 1972. [Google Scholar]
- Haccou, P.; Jagers, P.; Vatutin, V. Branching Processes: Variation, Growth and Extinction of Populations; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Mode, C.J. Multitype Branching Processes. Theory and Applications; American Elsevier Publishing Co., Inc.: New York, NY, USA, 1971. [Google Scholar]
- Quine, M.P. The multi-type Galton-Watson process with immigration. J. Appl. Prob. 1970, 7, 411–422. [Google Scholar] [CrossRef]
- Yakovlev, A.Y.; Yanev, N.M. Branching stochastic processes with immigration in analysis of renewing cell populations. Math. Biosci. 2006, 203, 37–63. [Google Scholar] [CrossRef] [PubMed]
- Yakovlev, A.Y.; Yanev, N.M. Limiting distributions for multitype branching processes. Stoch. Anal. Appl. 2010, 28, 1040–1060. [Google Scholar] [CrossRef] [PubMed]
- Crump, K.S.; Gillespie, J.H. The dispersion of a neutral allele considered as a branching process. J. Appl. Prob. 1976, 13, 208–218. [Google Scholar] [CrossRef]
- Pollak, E. Survival probabilities for some multitype branching processes in genetics. J. Math. Biol. 1992, 30, 583–596. [Google Scholar] [CrossRef]
- Durhan, S.D. An optimal branching migration process. J. Appl. Prob. 1975, 12, 569–573. [Google Scholar] [CrossRef]
- Dawson, D.A.; Greven, A. State dependent multitype spatial branching processes and their longtime behavior. Electron. J. Prob. 2003, 8, 1–93. [Google Scholar]
- Fairweather, D.W.; Shimi, I.N. A Multi-type branching process with immigration and random environment. Math. Biosci. 1972, 13, 299–324. [Google Scholar] [CrossRef]
- Vatutin, V.A. Multitype branching processes with immigration in random environment and polling systems. Siberian Adv. Math. 2011, 21, 42–72. [Google Scholar] [CrossRef]
- Horn, R.A.; Johnson, C.R. Topic in Matrix Analysis; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Kesten, H.; Stigum, B.P. A limit theorem for multidimensional Galton-Watson processes. Ann. Math. Statist. 1966, 37, 1211–1223. [Google Scholar] [CrossRef]
- Johnson, D. Substochastic Matrix Spectral Radius. Version: 2014-02-07. Available online: https://math.stackexchange.com/q/666603 (accessed on 18 December 2020).
- Kesten, H.; Stigum, B.P. Limit theorems for decomposble multi-dimensional Galton-Watson processes. J. Math. Anal. Appl. 1967, 17, 309–338. [Google Scholar] [CrossRef]
- Newton, I. Population Ecology of Raptors; T and AD Poyser Ltd.: London, UK, 1979. [Google Scholar]
- R Development Core Team. A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2009. Available online: http://www.r-project.org (accessed on 18 December 2020).


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molina-Fernández, M.; Mota-Medina, M. Demographic Dynamics in Multitype Populations with Migrations. Mathematics 2021, 9, 246. https://doi.org/10.3390/math9030246
Molina-Fernández M, Mota-Medina M. Demographic Dynamics in Multitype Populations with Migrations. Mathematics. 2021; 9(3):246. https://doi.org/10.3390/math9030246
Chicago/Turabian StyleMolina-Fernández, Manuel, and Manuel Mota-Medina. 2021. "Demographic Dynamics in Multitype Populations with Migrations" Mathematics 9, no. 3: 246. https://doi.org/10.3390/math9030246
APA StyleMolina-Fernández, M., & Mota-Medina, M. (2021). Demographic Dynamics in Multitype Populations with Migrations. Mathematics, 9(3), 246. https://doi.org/10.3390/math9030246