New Insights into Inflammatory Bowel Diseases from Proteomic and Lipidomic Studies


 , , ,
, , , 

Abstract

1. Introduction
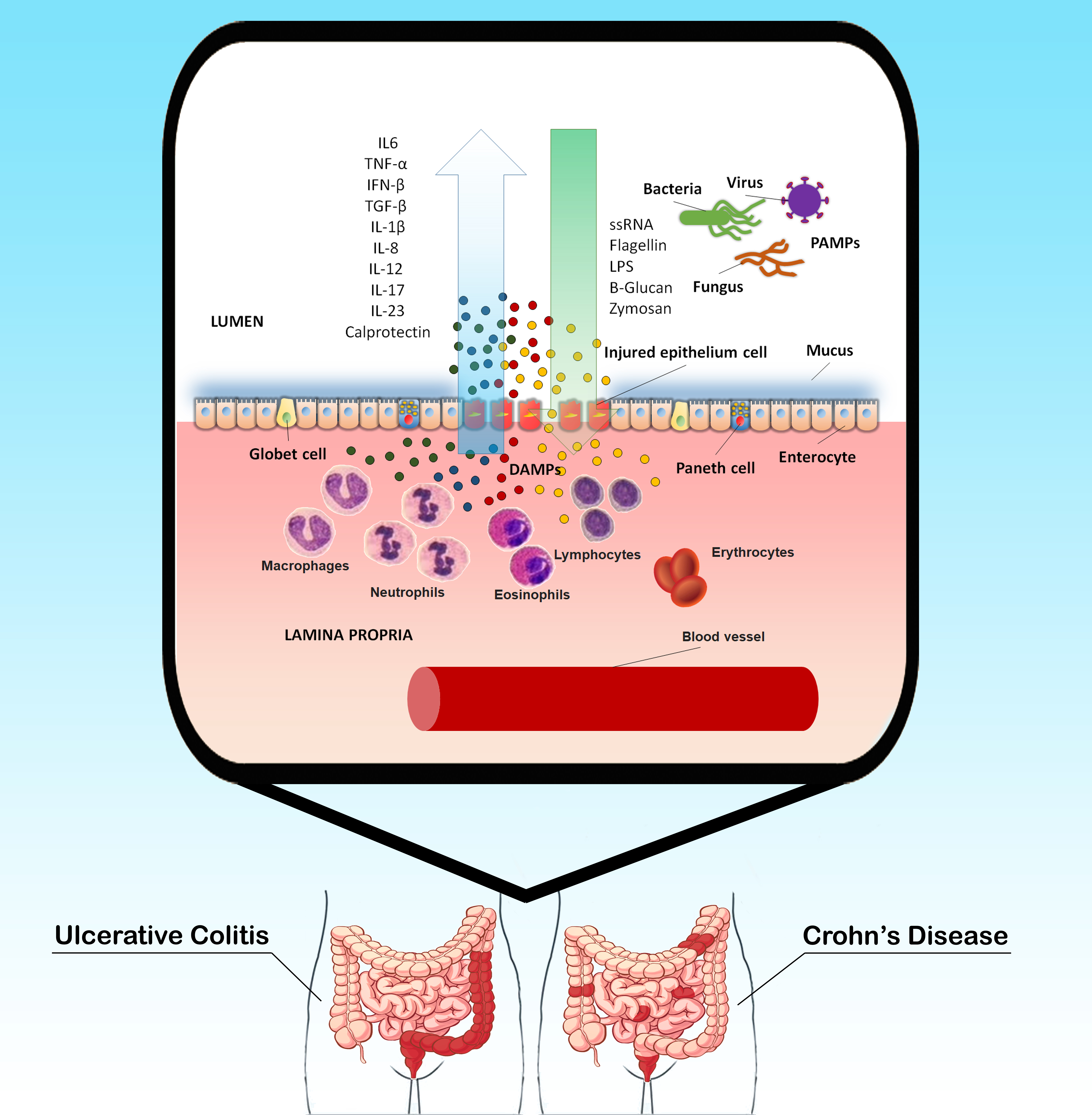
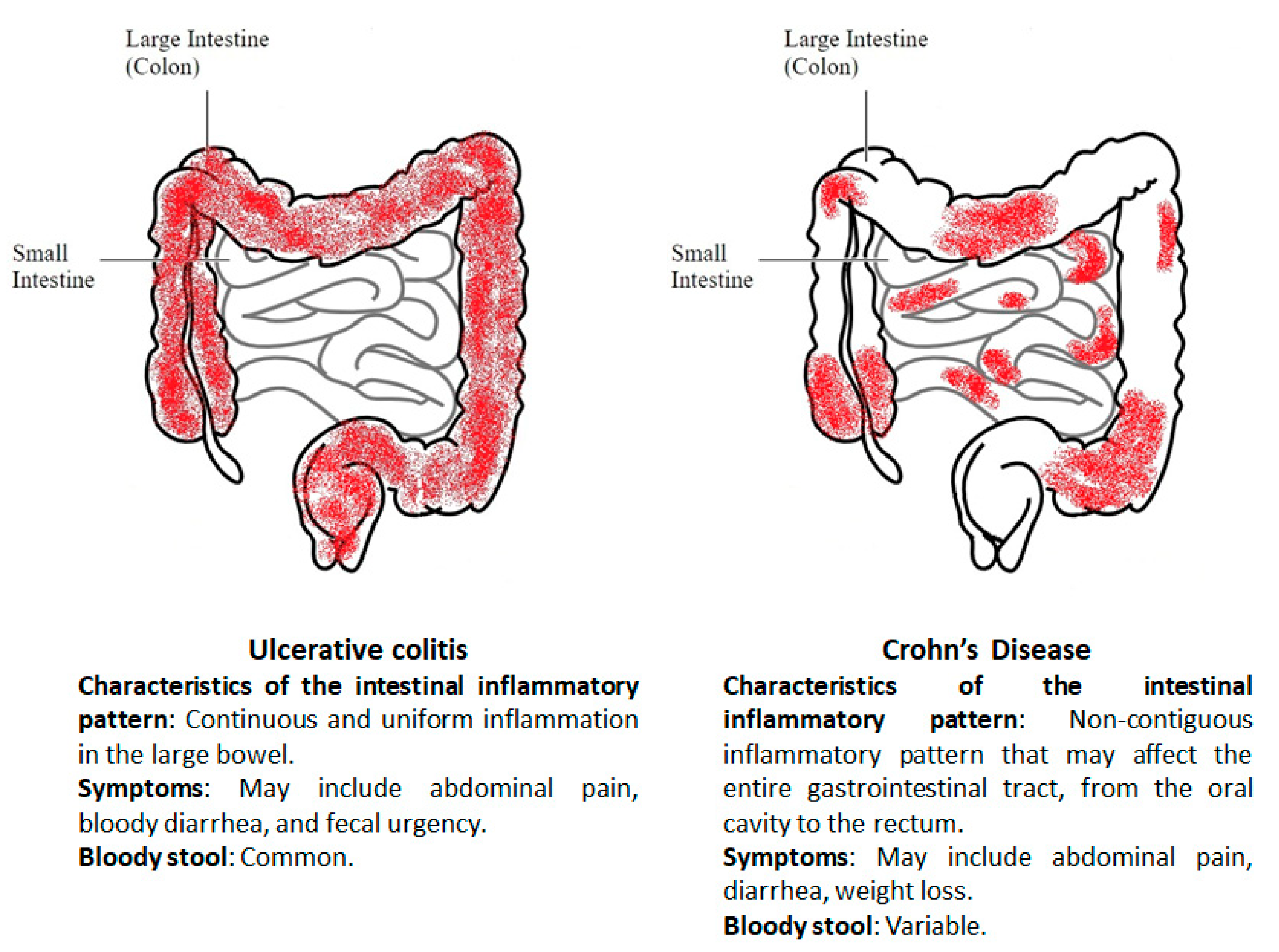
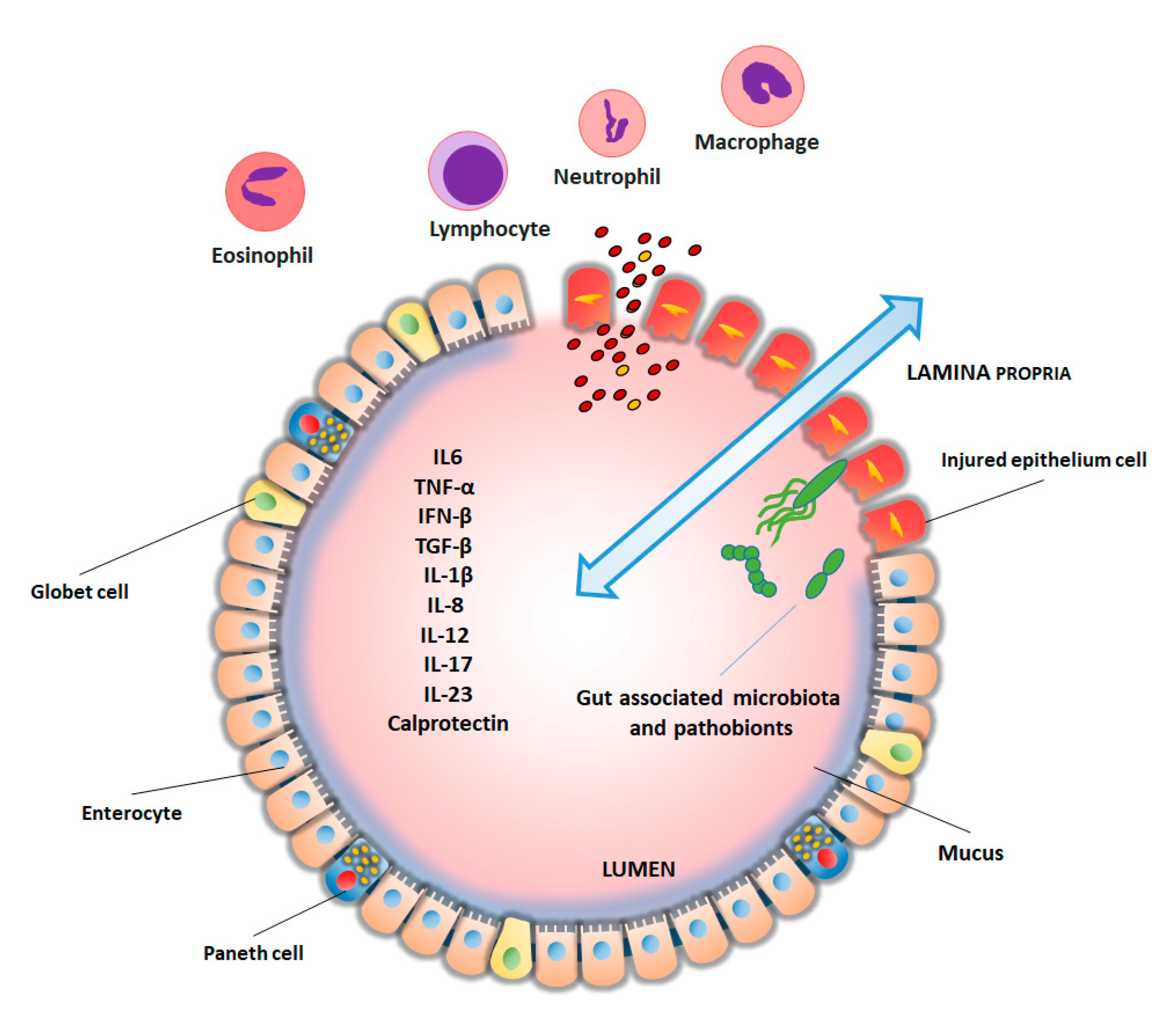
2. Biomarkers in Inflammatory Bowel Disease
3. New IBD Markers from Mass Spectrometry
Proteomic Analyses
4. Lipidomic Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Corridoni, D.; Arseneau, K.O.; Cominelli, F. Inflammatory bowel disease. Immunol. Lett. 2014, 161, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, B.A.; Gokhale, R.; Cho, J.H. Clinical aspects and pathophysiology of inflammatory bowel disease. Clin. Microbiol. Rev. 2002, 15, 79–94. [Google Scholar] [CrossRef]
- Gordon, I.O.; Agrawal, N.; Willis, E.; Goldblum, J.R.; Lopez, R.; Allende, D.; Liu, X.; Patil, D.J.; Yerian, L.; El-Khider, F.; et al. Fibrosis in ulcerative colitis is directly linked to severity and chronicity of mucosal inflammation. Aliment. Pharmacol. Ther. 2018, 47, 922–939. [Google Scholar] [CrossRef] [PubMed]
- Parray, F.Q.; Wani, M.L.; Malik, A.A.; Wani, S.N.; Bijli, A.H.; Nayeem-Ul, I.I. Ulcerative Colitis: A challenge to surgeons. Int. J. Prev. Med. 2012, 3, 749–763. [Google Scholar]
- Fuss, I.J.; Strober, W. The role of IL-13 and NK T cells in experimental and human ulcerative colitis. Mucosal Immunol. 2008, 1, S31–S33. [Google Scholar] [CrossRef]
- Parray, F.Q.; Wani, M.L.; Bijli, A.H.; Thakur, N.; Irshad, I.; Nayeem-ul-Hassan. Crohn’s Disease: A surgeon’s perspective. Saudi J. Gastroenterol. 2011, 17, 6–15. [Google Scholar]
- Nemeth, Z.H.; Bogdanovski, D.A.; Barratt-Stopper, P.; Paglinco, S.R.; Antonioli, L.; Rolandelli, R.H. Crohn’s disease and ulcerative colitis show unique cytokine profiles. Cureus 2017, 9, e1177. [Google Scholar] [CrossRef] [PubMed]
- Ananthakrishnan, A.N. Environmental risk factors for inflammatory bowel disease. Gastroenterol. Hepatol. 2013, 9, 367–374. [Google Scholar] [CrossRef]
- Mehta, M.; Ahmed, S.; Dryden, G. Immunopathophysiology of inflammatory bowel disease: How genetics link barrier dysfunction and innate immunity to inflammation. Innate Immun. 2017, 23, 497–505. [Google Scholar] [CrossRef]
- Liso, M.; De Santis, S.; Verna, G.; Dicarlo, M.; Calasso, M.; Santino, A.; Gigante, I.; Eri, R.; Raveenthiraraj, S.; Sobolewski, A.; et al. Specific mutation in muc2 determines early dysbiosis in colitis-prone winnie mice. Inflamm. Bowel Dis. 2020, 26, 546–556. [Google Scholar] [CrossRef]
- Rescigno, M.; Chieppa, M. Gut-level decisions in peace and war. Nat. Med. 2005, 11, 254–255. [Google Scholar] [CrossRef] [PubMed]
- Eri, R.; Chieppa, M. Messages from the Inside. The dynamic environment that favors intestinal homeostasis. Front. Immunol. 2013, 4, 323. [Google Scholar] [CrossRef] [PubMed]
- Vermeire, S.; Van Assche, G.; Rutgeerts, P. Laboratory markers in IBD: Useful, magic, or unnecessary toys? Gut 2006, 55, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.; Rosa, A.D.; Serao, R.; Picone, I.; Vietri, M.T. Laboratory markers in ulcerative colitis: Current insights and future advances. World. J. Gastrointest. Pathophysiol. 2015, 6, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Saverymuttu, S.H.; Hodgson, H.J.; Chadwick, V.S.; Pepys, M.B. Differing acute phase responses in Crohn’s disease and ulcerative colitis. Gut 1986, 27, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Shine, B.; Berghouse, L.; Jones, J.E.; Landon, J. C-reactive protein as an aid in the differentiation of functional and inflammatory bowel disorders. Clin. Chim. Acta 1985. [Google Scholar] [CrossRef]
- Papp, M.; Lakatos, P.L. Serological studies in inflammatory bowel disease: How important are they? Curr. Opin. Gastroenterol. 2014, 30, 359–364. [Google Scholar] [CrossRef]
- Mitsuyama, K.; Niwa, M.; Takedatsu, H.; Yamasaki, H.; Kuwaki, K.; Yoshioka, S.; Yamauchi, R.; Fukunaga, S.; Torimura, T. Antibody markers in the diagnosis of inflammatory bowel disease. World J. Gastroenterol. 2016, 22, 1304–1310. [Google Scholar] [CrossRef]
- Prideaux, L.; De Cruz, P.; Ng, S.C.; Kamm, M.A. Serological antibodies in inflammatory bowel disease: A systematic review. Inflamm. Bowel Dis. 2012, 18, 1340–1355. [Google Scholar] [CrossRef]
- Reese, G.E.; Constantinides, V.A.; Simillis, C.; Darzi, A.W.; Orchard, T.R.; Fazio, V.W.; Tekkis, P.P. Diagnostic precision of anti-Saccharomyces Cerevisiae antibodies and perinuclear antineutrophil cytoplasmic antibodies in inflammatory bowel disease. Am. J. Gastroenterol. 2006, 101, 2410–2422. [Google Scholar] [CrossRef]
- Konrad, A.; Rütten, C.; Flogerzi, B.; Styner, M.; Göke, B.; Seibold, F. Immune sensitization to yeast antigens in ASCA-positive patients with Crohn’s disease. Inflamm. Bowel Dis. 2004, 10, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Smids, C.; Horjus Talabur Horje, C.S.; Groenen, M.J.M.; van Koolwijk, E.H.M.; Wahab, P.J.; van Lochem, E.G. The value of serum antibodies in differentiating inflammatory bowel disease, predicting disease activity and disease course in the newly diagnosed patient. Scand. J. Gastroenterol. 2017, 52, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Peeters, M.; Joossens, S.; Vermeire, S.; Vlietinck, R.; Bossuyt, X.; Rutgeerts, P. Diagnostic value of anti-Saccharomyces Cerevisiae and antineutrophil cytoplasmic autoantibodies in inflammatory bowel disease. Am. J. Gastroenterol. 2001, 96, 730–734. [Google Scholar] [CrossRef] [PubMed]
- Joossens, S.; Reinisch, W.; Vermeire, S.; Sendid, B.; Poulain, D.; Peeters, M.; Geboes, K.; Bossuyt, X.; Vandewalle, P.; Oberhuber, G.; et al. The value of serological markers in indeterminate colitis: A prospective follow-up study. Gastroenetrology 2002, 122, 1242–1247. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.I.; Subramaniam, K.; Hawkins, C.A.; Randall, K.L. The significance of ANCA positivity in patients with inflammatory bowel disease. Pathology 2019, 51, 634–639. [Google Scholar] [CrossRef]
- Kuna, A.T. Serological markers in inflammatory bowel disease. Biochem. Med. 2013, 23, 28–42. [Google Scholar] [CrossRef]
- Hamilton, A.L.; Kamm, M.A.; De Cruz, P.; Wright, E.K.; Selvaraj, F.; Princen, F. Serological antibodies in relation to outcome in postoperative Crohn’s disease. J. Gastroenetrol. Hepatol. 2017, 32, 1195–1203. [Google Scholar] [CrossRef]
- Xiong, Y.; Wang, G.Z.; Zhou, J.Q.; Xia, B.Q.; Wang, X.Y.; Jiang, B. Serum antibodies to microbial antigens from Crohn’s disease progression: A mera analysis. Eur. J. Gastroenterol. Hepatol. 2014, 26, 733–742. [Google Scholar] [CrossRef]
- Kevans, D.; Waterman, M.; Milgrom, R.; Xu, W.; Van Assche, G.; Silverberg, M. Serological markers associated with disease behaviour and response to anti-tumor necrosis factor therapy in ulcerative colitis. J. Gastroenetrol. Hepatol. 2015, 30, 64–70. [Google Scholar] [CrossRef]
- Elkadri, A.A.; Stempak, J.M.; Walters, T.D.; Lal, S.; Griffiths, A.M.; Steinhart, A.H. Serum antibodies associated with complex inflammatory bowel disease. Inflamm. Bowel Dis. 2013, 19, 1499–1505. [Google Scholar] [CrossRef]
- Zhou, G.; Song, Y.; Yang, W.; Guo, Y.; Fang, L.; Chen, Y. ASCA, ANCA, ALCA, and many more: Are they useful in the diagnosis of inflammatory bowel disease? Dig. Dis. 2016, 34, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.; Riegle, G.; Vietri, M.T.; Pilla, P.; Caserta, L.; Carratù, R.; Sica, V.; Molinari, A.M. Serum p53 antibodies in patients affected with ulcerative colitis. Inflamm. Bowel Dis. 2004, 10, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Gisbert, J.P.; McNicholl, A.G.; Gomollon, F. Questions and answers on the role of fecal lactoferrin as a biological marker in inflammatory bowel disease. Inflamm. Bowel Dis. 2009, 15, 1746–1754. [Google Scholar] [CrossRef] [PubMed]
- Barnes, E.L.; Burakoff, R. The new biomarkers for diagnosing inflammatory bowel disease and assessing treatment outcomes. Inflamm. Bowel Dis. 2016, 22, 2956–2965. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.D. The utility of biomarkers in the diagnosis and therapy of inflammatory bowel disease. Gastroenterology 2011, 140, 1817–1826.e2. [Google Scholar] [CrossRef] [PubMed]
- Rugtveit, J.; Fagerhol, M.K. Age-dependent variations in fecal calprotectin concentrations in children. J. Pediatr. Gastroenterol. Nutr. 2002, 34, 324–325. [Google Scholar] [CrossRef]
- Bennike, T.; Birkelund, S.; Stensballe, A.; Andersen, V. Biomarkers in inflammatory bowel diseases: Current status and proteomics identification strategies. World J. Gastroenterol. 2014, 20, 3231–3244. [Google Scholar] [CrossRef]
- Vergara, D.; Simeone, P.; Franck, J.; Trerotola, M.; Giudetti, A.; Capobianco, L.; Tinelli, A.; Bellomo, C.; Fournier, I.; Gaballo, A.; et al. Translating epithelial mesenchymal transition markers into the clinic: Novel insights from proteomics. EuPA Open Proteom. 2016, 10, 31–41. [Google Scholar] [CrossRef]
- De Domenico, S.; Vergara, D. The Many-Faced Program of Epithelial-Mesenchymal Transition: A System Biology-Based View. Front. Oncol. 2017, 7, 274. [Google Scholar]
- Swietlik, J.J.; Sinha, A.; Meissner, F. Dissecting intercellular signaling with mass spectrometry-based proteomics. Curr. Opin. Cell Biol. 2020, 63, 20–30. [Google Scholar] [CrossRef]
- Bludau, I.; Aebersold, R. Proteomic and interactomic insights into the molecular basis of cell functional diversity. Nat. Rev. Mol. Cell. Biol. 2020, 21, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Saudemont, P.; Quanico, J.; Robin, Y.M.; Baud, A.; Balog, J.; Fatou, B.; Tierny, D.; Pascal, Q.; Minier, K.; Pottier, M.; et al. Real-Time molecular diagnosis of tumors using water-assisted laser desorption/ionization Mass Spectrometry technology. Cancer Cell. 2018, 34, 840–851. [Google Scholar] [CrossRef] [PubMed]
- Golf, O.; Strittmatter, N.; Karancsi, T.; Pringle, S.D.; Speller, A.V.; Mroz, A.; Kinross, J.M.; Abbassi-Ghadi, N.; Jones, E.A.; Takats, Z. Rapid evaporative ionization mass spectrometry imaging platform for direct mapping from bulk tissue and bacterial growth media. Anal. Chem. 2015, 87, 2527–2534. [Google Scholar] [CrossRef] [PubMed]
- Larance, M.; Lamond, A.I. Multidimensional proteomics for cell biology. Nat. Rev. Mol. Cell Biol. 2015, 16, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, Y.; Lamond, A.I. A perspective on proteomics in cell biology. Trends Cell Biol. 2014, 24, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Barceló-Batllori, S.; André, M.; Servis, C.; Lévy, N.; Takikawa, O.; Michetti, P.; Reymond, M.; Felley-Bosco, E. Proteomic analysis of cytokine induced proteins in human intestinal epithelial cells: Implications for inflammatory bowel diseases. Proteomics 2002, 2, 551–560. [Google Scholar] [CrossRef]
- Poulsen, N.A.; Andersen, V.; Møller, J.C.; Møller, H.S.; Jessen, F.; Purup, S.; Larsen, L.B. Comparative analysis of inflamed and non-inflamed colon biopsies reveals strong proteomic inflammation profile in patients with ulcerative colitis. BMC Gastroenterol. 2012, 24, 12–76. [Google Scholar] [CrossRef]
- M’Koma, A.E.; Seeley, E.H.; Washington, M.K.; Schwartz, D.A.; Muldoon, R.L.; Herline, A.J.; Wise, P.E.; Caprioli, R.M. Proteomic profiling of mucosal and submucosal colonic tissues yields protein signatures that differentiate the inflammatory colitis. Inflamm. Bowel Dis. 2011, 17, 875–883. [Google Scholar] [CrossRef]
- Seeley, E.H.; Washington, M.K.; Caprioli, R.M.; M’Koma, A.E. Proteomic patterns of colonic mucosal tissues delineate Crohn’s colitis and ulcerative colitis. Proteom. Clin. Appl. 2013, 7, 541–549. [Google Scholar] [CrossRef]
- Arafah, K.; Kriegsmann, M.; Renner, M.; Lasitschka, F.; Fresnais, M.; Kriegsmann, K.; von Winterfeld, M.; Goeppert, B.; Kriegsmann, J.; Casadonte, R.; et al. Microproteomics and immunohistochemistry reveal differences in aldo-keto reductase family 1 member C3 in tissue specimens of Ulcerative Colitis and Crohn’s Disease. Proteom. Clin. Appl. 2020, 31, e1900110. [Google Scholar] [CrossRef]
- Hsieh, S.Y.; Shih, T.C.; Yeh, C.Y.; Lin, C.J.; Chou, Y.Y.; Lee, Y.S. Comparative proteomic studies on the pathogenesis of human ulcerative colitis. Proteomics 2006, 6, 5322–5331. [Google Scholar] [CrossRef] [PubMed]
- Hatsugai, M.; Kurokawa, M.S.; Kouro, T.; Nagai, K.; Arito, M.; Masuko, K.; Suematsu, N.; Okamoto, K.; Itoh, F.; Kato, T. Protein profiles of peripheral blood mononuclear cells are useful for differential diagnosis of ulcerative colitis and Crohn’s disease. J. Gastroenterol. 2010, 45, 488–500. [Google Scholar] [CrossRef] [PubMed]
- Shkoda, A.; Werner, T.; Daniel, H.; Gunckel, M.; Rogler, G.; Haller, D. Differential protein expression profile in the intestinal epithelium from patients with inflammatory bowel disease. J. Proteome Res. 2007, 6, 1114–1125. [Google Scholar] [CrossRef] [PubMed]
- Basso, D.; Padoan, A.; D’Incà, R.; Arrigoni, G.; Scapellato, M.L.; Contran, N.; Franchin, C.; Lorenzon, G.; Mescoli, C.; Moz, S.; et al. Peptidomic and proteomic analysis of stool for diagnosing IBD and deciphering disease pathogenesis. Clin. Chem. Lab. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ning, L.; Shan, G.; Sun, Z.; Zhang, F.; Xu, C.; Lou, X.; Li, S.; Du, H.; Chen, H.; Xu, G. Quantitative proteomic analysis reveals the deregulation of nicotinamide adenine dinucleotide metabolism and CD38 in Inflammatory Bowel Disease. Biomed. Res. Int. 2019, 2019, 3950628. [Google Scholar] [CrossRef]
- van der Post, S.; Jabbar, K.S.; Birchenough, G.; Arike, L.; Akhtar, N.; Sjovall, H.; Johansson, M.E.V.; Hansson, G.C. Structural weakening of the colonic mucus barrier is an early event in ulcerative colitis pathogenesis. Gut 2019, 68, 2142–2151. [Google Scholar] [CrossRef]
- Moriggi, M.; Pastorelli, L.; Torretta, E.; Tontini, G.E.; Capitanio, D.; Bogetto, S.F.; Vecchi, M.; Gelfi, C. Contribution of extracellular matrix and signal mechanotransduction to epithelial cell damage in inflammatory bowel disease patients: A proteomic study. Proteomics 2017, 17, 23–24. [Google Scholar] [CrossRef]
- Starr, A.E.; Deeke, S.A.; Ning, Z.; Chiang, C.K.; Zhang, X.; Mottawea, W.; Singleton, R.; Benchimol, E.I.; Wen, M.; Mack, D.R.; et al. Proteomic analysis of ascending colon biopsies from a paediatric inflammatory bowel disease inception cohort identifies protein biomarkers that differentiate Crohn’s disease from UC. Gut 2017, 66, 1573–1583. [Google Scholar] [CrossRef]
- Magnusson, M.K.; Strid, H.; Isaksson, S.; Bajor, A.; Lasson, A.; Ung, K.A.; Öhman, L. Response to infliximab therapy in ulcerative colitis is associated with decreased monocyte activation, reduced CCL2 expression and downregulation of Tenascin C. J. Crohns. Colitis. 2015, 9, 56–65. [Google Scholar] [CrossRef]
- Bennike, T.B.; Carlsen, T.G.; Ellingsen, T.; Bondeup, O.K.; Glerup, H.; Bøgsted, M.; Christiansen, G.; Birkelund, S.; Stensballe, A.; Andersen, V. Neutrophil extracellular traps in ulcerative colitis: A proteome analysis of intestinal biopsies. Inflamm Bowel. Dis. 2015, 21, 2052–2067. [Google Scholar] [CrossRef]
- Nanni, P.; Mezzanotte, L.; Roda, G.; Caponi, A.; Levander, F.; James, P.; Roda, A. Differential proteomic analysis of HT29 Cl.16E and intestinal epithelial cells by LC ESI/QTOF mass spectrometry. J. Proteom. 2009, 72, 865–873. [Google Scholar] [CrossRef] [PubMed]
- German, J.B.; Gillies, L.A.; Smilowitz, J.T.; Zivkovic, A.M.; Watkins, S.M. Lipidomics and lipid profiling in metabolomics. Curr. Opin. Lipidol. 2007, 18, 66–71. [Google Scholar] [PubMed]
- Feng, L.; Prestwich, G.D. Functional Lipidomics; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2006. [Google Scholar]
- Kraegen, E.W.; Cooney, G.J.; Ye, J.M.; Thompson, A.L.; Furler, S.M. The Role of Lipids in the Pathogenesis of Muscle Insulin Resistance and Beta Cell Faiture in Type II Diabetes and Obesity. Exp. Clin. Endocrinol. Diabetes 2001, 109, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.; Watkins, S.M. Focused metabolomic profiling in the drug development process: Advances from lipid profiling. Curr. Opin. Chem. Biol. 2005, 9, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Wenk, M.R. The Emerging Field of Lipidomics. Nat. Rev. Drug Discov. 2005, 4, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Bazarganipour, S.; Hausmann, J.; Oertel, S.; El-Hindi, K.; Brachtendorf, S.; Blumenstein, I.; Kubesch, A.; Sprinzl, K.; Birod, K.; Hahnefeld, L.; et al. The lipid status in patients with Ulcerative Colitis: Sphingolipids are disease-dependent degulated. J. Clin. Med. 2019, 8, 971. [Google Scholar] [CrossRef]
- Diab, J.; Al-Mahdi, R.; Gouveia-Figueira, S.; Hansen, T.; Jensen, E.; Goll, R.; Moritz, T.; Florholmen, J.; Forsdahl, G.A. Quantitative analysis of colonic mucosal oxylipins and endocannabinoids in treatment-naïve and deep remission Ulcerative Colitis patients and the potential link with cytokine gene expression. Inflamm. Bowel Dis. 2019, 25, 490–497. [Google Scholar] [CrossRef]
- Masoodi, M.; Pearl, D.S.; Eiden, M.; Shute, J.K.; Brown, J.F.; Calder, P.C.; Trebble, T.M. Altered colonic mucosal Polyunsaturated Fatty Acid (PUFA) derived lipid mediators in ulcerative colitis: New insight into relationship with disease activity and pathophysiology. PLoS ONE 2013, 8, e76532. [Google Scholar] [CrossRef]
- Ehehalt, R.; Wagenblast, J.; Erben, G.; Lehmann, W.D.; Hinz, U.; Merle, U.; Stremmel, W. Phosphatidylcholine and lysophosphatidylcholine in intestinal mucus of ulcerative colitis patients. A quantitative approach by nanoElectrospray-tandem mass spectrometry. Scand. J. Gastroenterol. 2004, 39, 737–742. [Google Scholar] [CrossRef]
- Braun, A.; Treede, I.; Gotthardt, D.; Tietje, A.; Zahn, A.; Ruhwald, R.; Schoenfeld, U.; Welsch, T.; Kienle, P.; Erben, G. Alterations of phospholipid concentration and species composition of the intestinal mucus barrier in ulcerative colitis: A clue to pathogenesis. Inflamm. Bowel Dis. 2009, 15, 1705–1720. [Google Scholar] [CrossRef]
- Fan, F.; Mundra, P.A.; Fang, L.; Galvin, A.; Moore, X.L.; Weir, J.M.; Wong, G.; White, D.A.; Chin-Dusting, J.; Sparrow, M.P.; et al. Lipidomic profiling in inflammatory bowel disease: Comparison between ulcerative colitis and Crohn’s disease. Inflamm. Bowel Dis. 2015, 21, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Santoru, M.L.; Piras, C.; Murgia, A.; Palmas, V.; Camboni, T.; Liggi, S.; Ibba, I.; Lai, M.A.; Orrù, S.; Blois, S.; et al. Cross sectional evaluation of the gut-microbiome metabolome axis in an Italian cohort of IBD patients. Sci. Rep. 2017, 7, 9523. [Google Scholar] [CrossRef]
- Scoville, E.A.; Allaman, M.M.; Brown, C.T.; Motley, A.K.; Horst, S.N.; Williams, C.S.; Koyama, T.; Zhao, Z.; Adams, D.W.; Beaulieu, D.B.; et al. Alterations in lipid, amino acid, and energy metabolism distinguish Crohn’s Disease from Ulcerative Colitis and control subjects by serum Metabolomic Profiling. Metabolomics 2018, 14, 17. [Google Scholar] [CrossRef] [PubMed]
- Murgia, A.; Hinz, C.; Liggi, S.; Denes, J.; Hall, Z.; West, J.; Santoru, M.L.; Piras, C.; Manis, C.; Usai, P.; et al. Italian cohort of patients affected by inflammatory bowel disease is characterised by variation in glycerophospholipid, free fatty acids and amino acid levels. Metabolomics 2018, 14, 140. [Google Scholar] [CrossRef] [PubMed]
- Usami, M.; Komurasaki, T.; Hanada, A.; Kinoshita, K.; Ohata, A. Effect of gamma-linolenic acid or docosahexaenoic acid on tight junction permeability in intestinal monolayer cells and their mechanism by proteinkinase C activation and/or eicosanoid formation. Nutrition 2003, 19, 150–156. [Google Scholar] [CrossRef]
- Barbosa, D.S.; Cecchini, R.; El Kadri, M.Z.; Rodriguez, M.A.; Burini, R.C.; Dichi, I. Decreased oxidative stress in patients with ulcerative colitis supplemented with fish oil omega-3 fatty acids. Nutrition 2003, 19, 837–842. [Google Scholar] [CrossRef]
- Heimerl, S.; Moehle, C.; Zahn, A.; Boettcher, A.; Stremmel, W.; Langmann, T.; Schmitz, G. Alterations in intestinal fatty acid metabolism in inflammatory bowel disease. Biochim. Biophys. Acta 2006, 1762, 341–350. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Biomarker | Source | Ref. |
|---|---|---|
| C-reactive protein | Serum | [16] |
| Erythrocyte sedimentation rate | Blood | [13] |
| Anti-Saccharomyces cerevisiae (ASCA) | Serum | [21] |
| Perinuclear antineutrophil cytoplasmic antibody (pANCA) | Serum | [19,20] |
| Anti-porin (Anti-OmpC) | Serum | [26,27,28] |
| Anti-Cbir1 Flagellin (anti-CBir1) | Serum | [29] |
| Anti-Pseudomonas fluorescens-associated sequence I-2 (Anti-I2) | Serum | [30] |
| Fecal calprotectin | Stool | [32] |
| Lactoferrin | Stool | [36] |
| Key Findings | Biological Sample | Separation | Instrument | Ref. |
|---|---|---|---|---|
| Higher abundance, in IBD vs. normal mucosa, of indoleamine-2,3-dioxygenase. | Colonic epithelial cells from ulcerative colitis (UC) and Crohn’s disease (CD) patients | 2-DE | MALDI-TOF-MS | [46] |
| Distinct profile in UC vs. controls of proteins involved in energy metabolism and oxidative stress. | Colonic biopsies from UC and controls | 2-DE | MALDI-TOF-MS | [47] |
| Identification of a different proteomic signature between CD and CD. | Colonic mucosal and submucosal layers from CD and UC | On-tissue analysis | Histology-directed MALDI-TOF-MS | [48] |
| Identification of a different proteomic signature between CD and CD. | Histologic layers from UC and CD | On-tissue analysis | Histology-directed MALDI-TOF-MS | [49] |
| Higher abundance in CD compared to UC of the Aldo-keto reductase family 1 member C3 (AKR1C3) protein. | Colonic tissue specimens from UC and CD | Laser microdissection and chromatography | LC-MS/MS | [50] |
| Down-regulation of different mitochondrial proteins involved in energy generation and stress response. | Colonic biopsies from UC, nonspecific colitis patients and controls | 2-DE | MALDI-TOF-MS | [51] |
| Identification of a different UC and CD protein signature, comprising proteins associated with inflammation, oxidation/reduction, the cytoskeleton, endocytic trafficking and transcription. | Peripheral blood mononuclear cells from UC and CD | 2-DE | MALDI-TOF-MS | [52] |
| Up-regulation of Rho-GDP dissociation inhibitor alpha in CD and UC patients. In UC vs. noninflamed higher level of programmed cell death proteins and annexin 2A. | Human primary intestinal epithelial cells obtained from CD, UC and control patients | 2-DE | MALDI-TOF-MS | [53] |
| Higher level in IBD of immunoglobulins and neutrophil proteins. Lower level in IBD of the nucleic acid assembly proteins or OLFM4, ENPP7, related to cancer risk. | Stools from IBD and control patients | Peptide analysis and chromatography | MALDI-TOF-MS/MS and LC-MS/MS | [54] |
| Higher level in CD and UC of CD38 and angiotensin-converting enzymes 1 and 2. | Colonic biopsies from UC, CD and controls | Chromatography | LC-MS/MS | [55] |
| Reduced amount in active UC of mucin MUC2 and SLC26A3. | Mucus samples from inflamed or in remission UC patients | Chromatography | LC-MS/MS | [56] |
| Changes in UC and CD of proteins of extracellular matrix, cytoskeletal, cellular metabolism, and autophagy. | Colonic biopsies from UC patients and controls | 2DE and chromatography | MALDI-TOF-MS and ICPL-LC-MS/MS | [57] |
| 5 different proteins discriminate IBD from control patients and a 12-protein panel was proposed to distinguish CD from UC patients. | Mucosal biopsies from IBD children and controls | Chromatography | LC-MS/MS | [58] |
| Increased amount in UC of proteins involved in the innate immune system. | Colonic biopsies from controls and UC patients | Chromatography | LC-MS/MS | [60] |
| Up-regulation in CD patients of nuclear histones and ubiquitin, of cytosolic tryptase alpha-1 precursor and of membrane ATP synthase subunit beta and Heat shock 70kDa protein 5. | Subcellular fractions of intestinal epithelium cells from healthy donors and CD patients | Chromatography | LC-MS/MS | [61] |
| Abbreviations: two-dimensional electrophoresis (2-DE), matrix assisted laser desorption ionization-mass spectrometry (MALDI-MS), time of flight (TOF), difference gel electrophoresis (DIGE), liquid chromatography-mass spectrometry/mass spectrometry (LC-MS/MS), Isotope-Coded Protein Labeling (ICPL). | ||||
| Key Findings | Biological Samples | Instrument | Ref. |
|---|---|---|---|
| In inflamed colon tissue de novo-synthesis of sphingolipids was reduced, whereas lactosylceramides were increased. Plasma sphingolipids, free fatty acids, lyso-PC and triacylglycerols changed significantly in UC in comparison to healthy controls. | Blood and colon tissue samples from UC patients and healthy controls | LC-MS/MS and LC-QTOF-MS | [67] |
| Debut of UC is associated with increased levels of ω-6-related oxylipins and decreased levels of ω-3-related endocannabinoids. | Colon biopsies from treatment-naïve UC patients, deep remission UC patients, and healthy controls | GC-TOF-MS and UHPLC-M | [68] |
| Levels of PGE2, PGD2, TXB2, 5-HETE, 11-HETE, 12-HETE and 15-HETE were significantly elevated in inflamed mucosa and correlated with severity of inflammation. | Mucosal biopsies from relapsing UC | LC-MS/MS | [69] |
| Significantly less PC and lyso-PC in patients with inactive UC compared to CD and control subjects. | Rectal mucus from patients with UC, CD and healthy controls | Nano-ESI-MS/MS | [70] |
| Lower PC concentration in UC compared to CD patients and controls. Independent of disease activity, patients suffering from UC showed an increased saturation grade of PC fatty acid residues and a higher lyso-PC-to-PC ratio. | Colonic mucus from UC, CD and healthy controls | Nano ESI-MS/MS | [71] |
| Lipid species belonging to ethers and plasmalogens were significantly changed in CD patients compared with controls. Only 5 lipid species significantly differed between UC and controls. | Plasma from IBD and controls | LC-ESI-MS/MS | [72] |
| Biogenic amines, amino acids, lipids, were significantly increased in IBD, while others, such as two B group vitamins, were decreased in IBD compared to healthy subjects. | Stools from UC, CD and healthy controls | GC-MS and LC-QTOF-MS | [73] |
| Several lipid-, amino acid-, and tricarboxylic acid cycle-related metabolites were significantly altered in CD. Instead, only 5 metabolites decreased in UC with respect to control subjects. | Serum from UC, CD and healthy controls | UPLC-MS/MS | [74] |
| PC, lyso-PC and fatty acids were significantly changed among pathological samples. Variations in the levels of cholesteryl esters and glycerophospholipids were also found. | Plasma from IBD and healthy controls | LC-TOF-MS | [75] |
| Abbreviations: phosphatidylcholine (PC), lyso-phosphatidylcholines (lyso-PC), prostaglandin (PG), tromboxane 2 (TXB2), hydroxyeicosatetraenoic acid (HETE). | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Longo, S.; Chieppa, M.; Cossa, L.G.; Spinelli, C.C.; Greco, M.; Maffia, M.; Giudetti, A.M. New Insights into Inflammatory Bowel Diseases from Proteomic and Lipidomic Studies. Proteomes 2020, 8, 18. https://doi.org/10.3390/proteomes8030018
Longo S, Chieppa M, Cossa LG, Spinelli CC, Greco M, Maffia M, Giudetti AM. New Insights into Inflammatory Bowel Diseases from Proteomic and Lipidomic Studies. Proteomes. 2020; 8(3):18. https://doi.org/10.3390/proteomes8030018
Chicago/Turabian StyleLongo, Serena, Marcello Chieppa, Luca G. Cossa, Chiara C. Spinelli, Marco Greco, Michele Maffia, and Anna M. Giudetti. 2020. "New Insights into Inflammatory Bowel Diseases from Proteomic and Lipidomic Studies" Proteomes 8, no. 3: 18. https://doi.org/10.3390/proteomes8030018
APA StyleLongo, S., Chieppa, M., Cossa, L. G., Spinelli, C. C., Greco, M., Maffia, M., & Giudetti, A. M. (2020). New Insights into Inflammatory Bowel Diseases from Proteomic and Lipidomic Studies. Proteomes, 8(3), 18. https://doi.org/10.3390/proteomes8030018