
The Cytosolic Oligosaccharide-Degrading Proteome of Butyrivibrio Proteoclasticus

Abstract
:
1. Introduction
2. Experimental Section
2.1. Sample Preparation
2.2. Two-Dimensional Electrophoresis and Image Analysis
2.3. Identification of Proteins Excised from 2DE Gels
2.4. LC-MS/MS
2.5. Bioinformatics
3. Results and Discussion
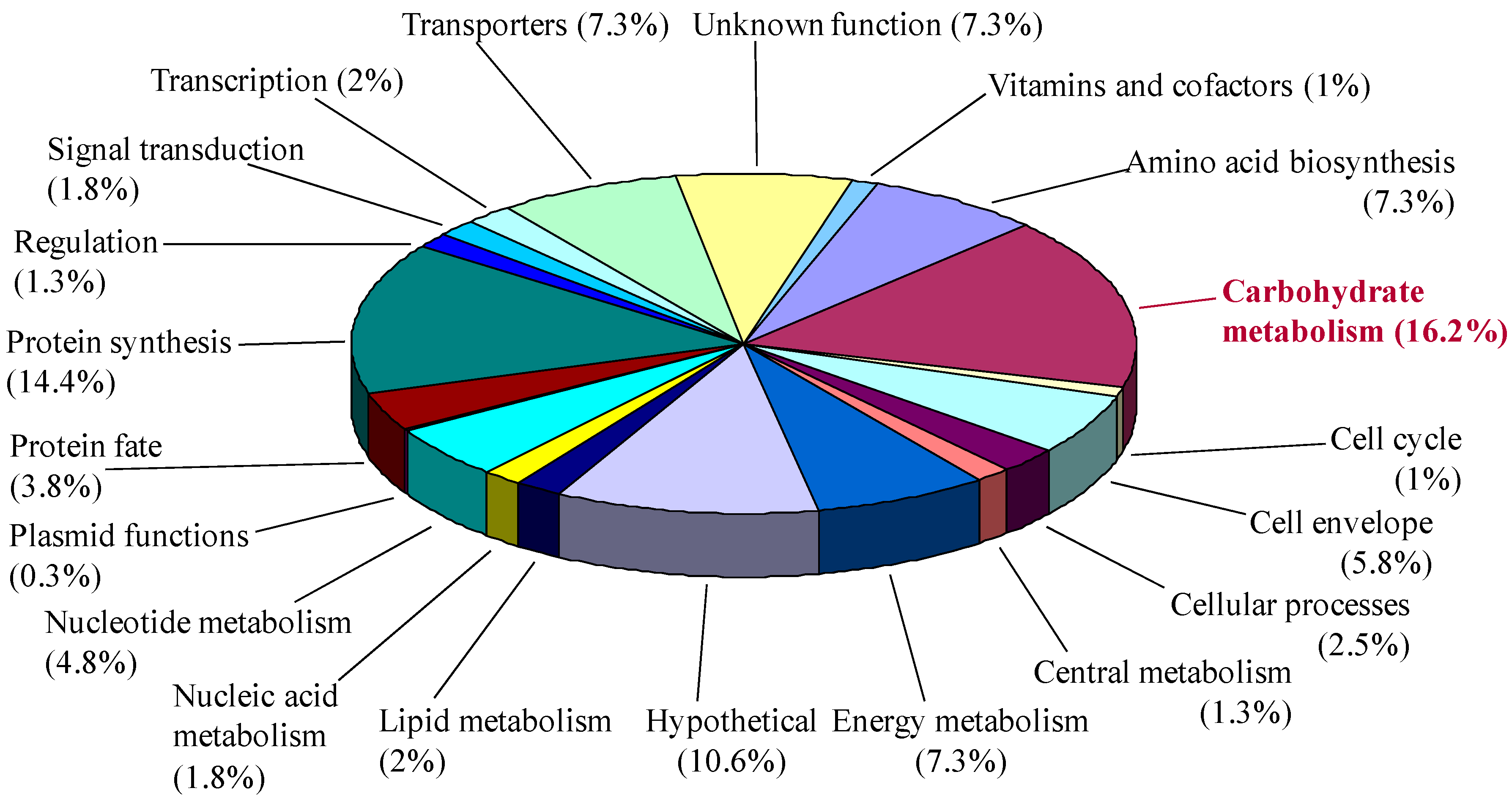
3.1. Theoretical Proteome
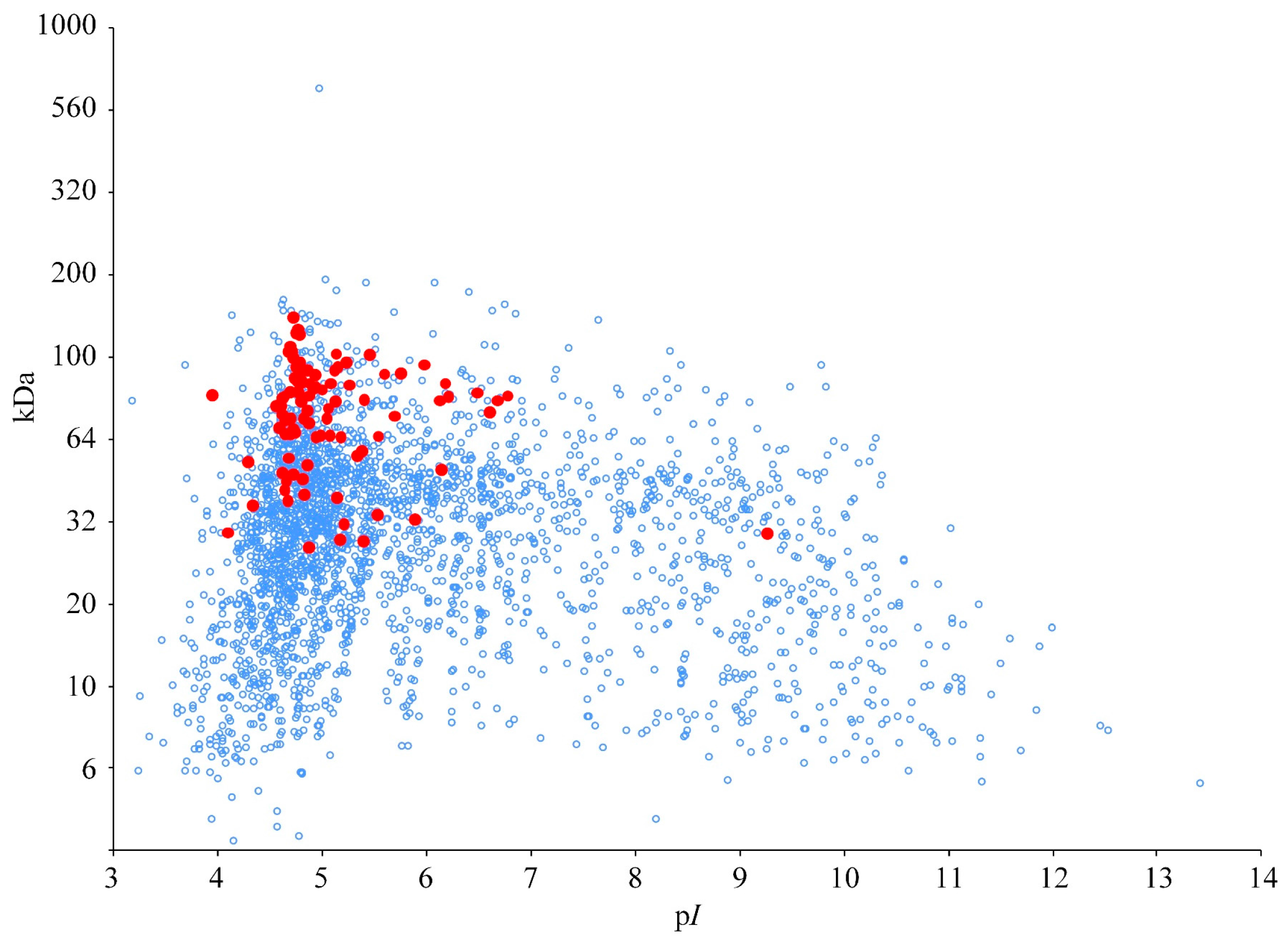
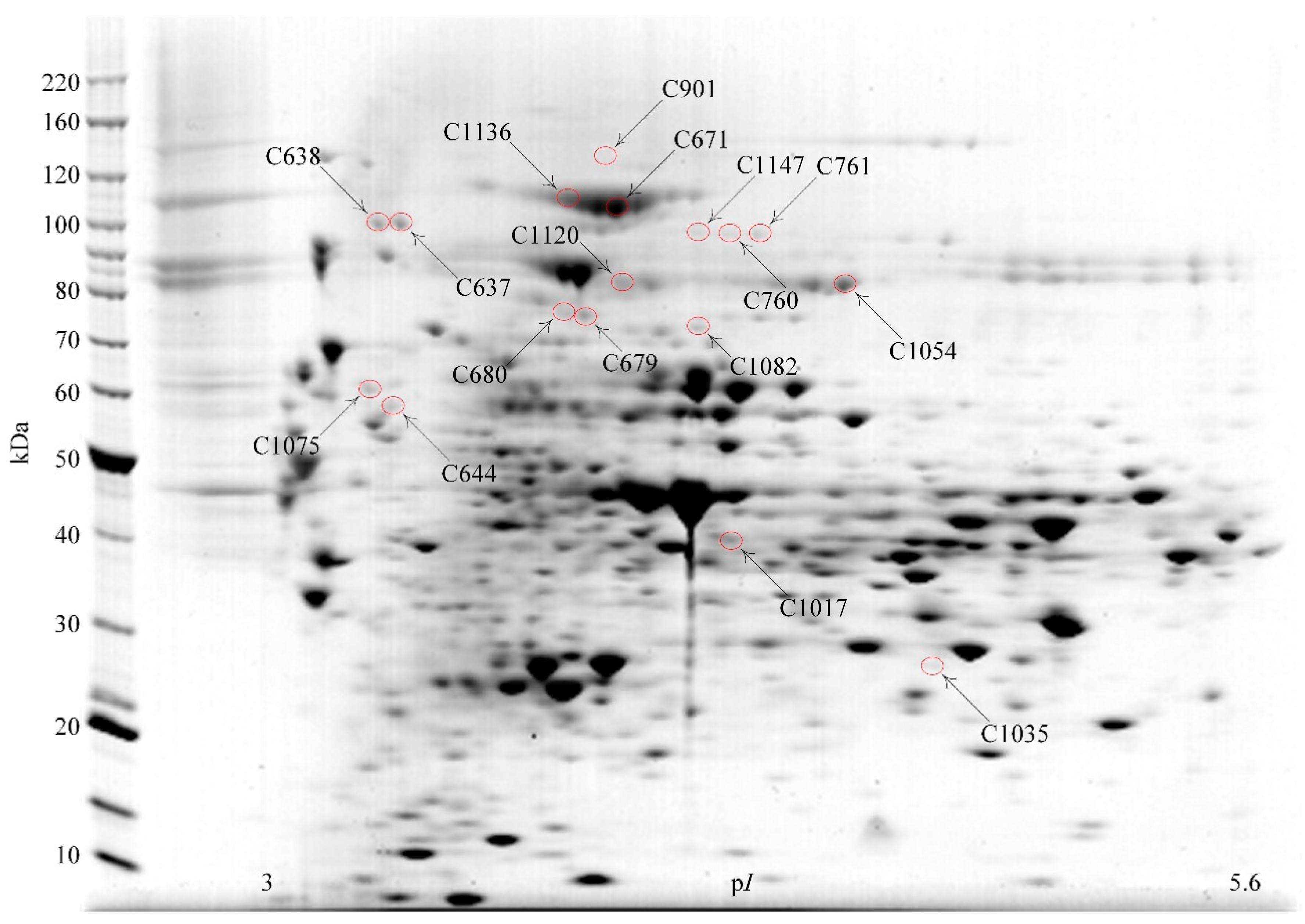
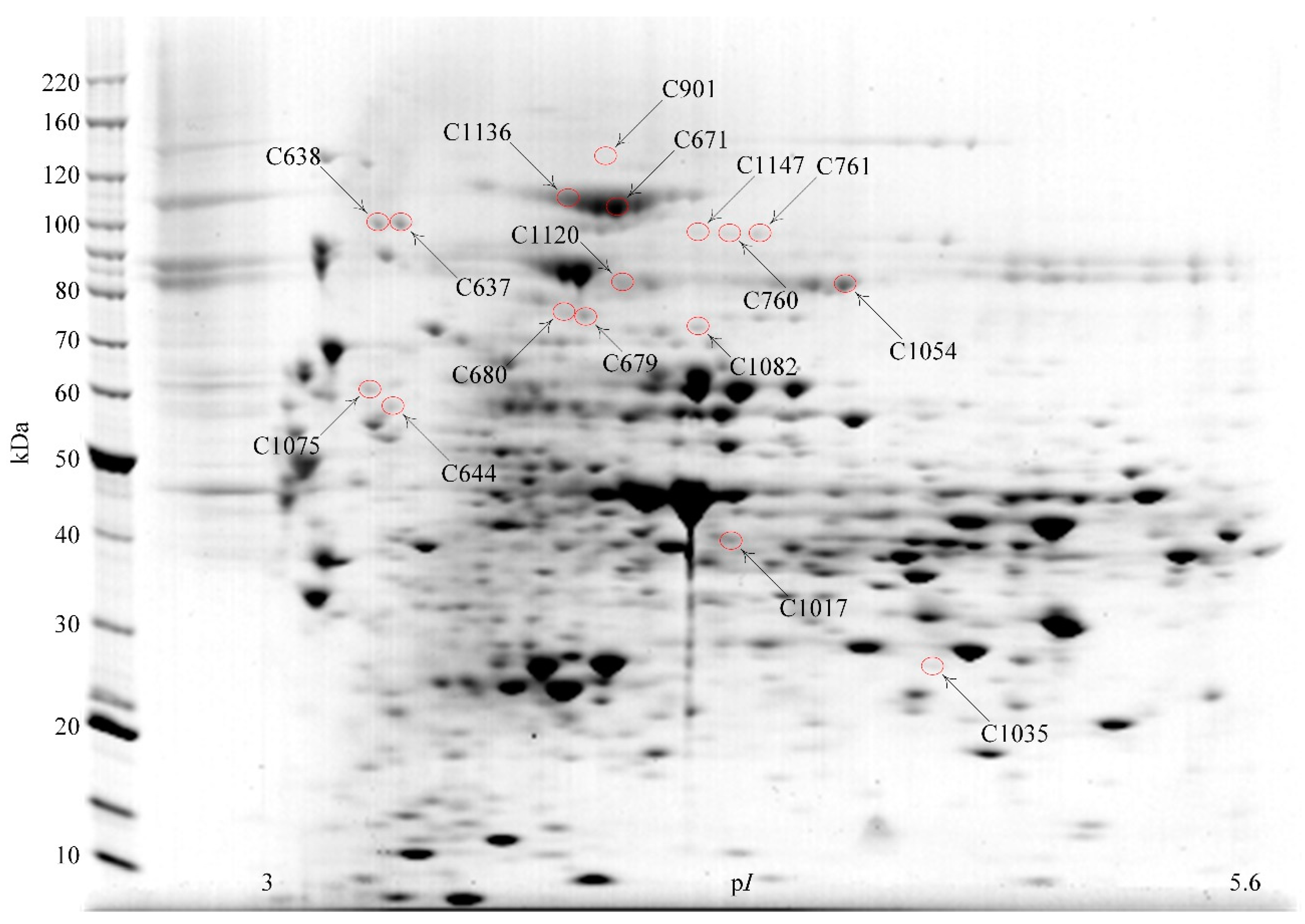
3.2. Identification of Proteins Separated by 2DE

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot | Protein | Locus | Protein Pilot Score | Expect. i | SigP ii | pI | kDa | Pep iii |
|---|---|---|---|---|---|---|---|---|
| C901 | β-Galactosidase, Bga2A | Bpr_I0279 | 239 | 4.8 × 10−21 | N | 4.7 | 118.7 | 31 |
| C1147 | β-Galactosidase, Bga2B | Bpr_III209 | 80 | 4.1 × 10−5 | N | 4.8 | 91.1 | 19 |
| C1147 | β-Mannosidase, Man2A | Bpr_III237 | 117 | 3.8 × 10−8 | N | 4.8 | 95.9 | 20 |
| C1147 | β-Glucosidase, Bgl3C | Bpr_I0138 | 73 | 1.8 × 10−4 | N | 4.9 | 91.5 | 10 |
| C1136 | β-Glucosidase, Bgl3B | Bpr_I0847 | 94 | 1.4 × 10−6 | N | 4.7 | 103.7 | 24 |
| C671 | β-Glucosidase, Bgl3B | Bpr_I0847 | 218 | 6.1 × 10−19 | N | 4.7 | 91.5 | 23 |
| C760 | β-Glucosidase, Bgl3C | Bpr_I0138 | 72 | 2.4 × 10−4 | N | 4.9 | 91.5 | 7 |
| C761 | β-Glucosidase, Bgl3C | Bpr_I0138 | 231 | 3.0 × 10−20 | N | 4.9 | 103.7 | 23 |
| C1120 | β-Xylosidase, Xyl3A | Bpr_I0184 | 397 | 7.6 × 10−37 | N | 4.8 | 78.2 | 28 |
| C1120 | β-Galactosidase, Bga35B | Bpr_I2006 | 87 | 6.9 × 10−6 | N | 4.9 | 83.2 | 12 |
| C637 | Pullulanase, Pul13A | Bpr_III161 | 191 | 3.0 × 10−16 | Y | 4.4 | 99.7 | 19 |
| C638 | Pullulanase, Pul13A | Bpr_III161 | 243 | 1.9 × 10−21 | Y | 4.4 | 99.7 | 22 |
| C644 | Glycoside hydrolase family 30, GH30A | Bpr_I2937 | 261 | 3.0 × 10−23 | Y | 4.3 | 67.0 | 23 |
| C679 | Glycoside hydrolase family 31, GH31C | Bpr_I1974 | 301 | 3.0 × 10−27 | N | 4.7 | 78.2 | 40 |
| C680 | Glycoside hydrolase family 31, GH31C | Bpr_I1974 | 248 | 6.1 × 10−22 | N | 4.7 | 78.2 | 39 |
| C1054 | α-Galactosidase, Aga36C | Bpr_III065 | 315 | 1.2 × 10−28 | N | 5.1 | 83.2 | 30 |
| C1075 | Xylosidase/arabino-furanosidase, Xsa43A | Bpr_I0302 | 110 | 3.8 × 10−8 | Y | 4.3 | 57.5 | 17 |
| C1082 | α-D-Glucuronidase, Agu67A | Bpr_I0177 | 341 | 3.0 × 10−31 | N | 4.9 | 76.1 | 32 |
| C1035 | Feruloyl esterase, Est1E | Bpr_I2870 | 82 | 2.7 × 10−5 | N | 5.2 | 27.8 | 13 |
| C1017 | Acetyl-xylan esterase, Est2A | Bpr_I2939 | 235 | 1.2 × 10−20 | N | 4.8 | 42.4 | 8 |
3.3. Protein Identification by LC-MS/MS
| Protein | Locus | Expect. ii | SigP iii | pI | kDa | Pep iv |
|---|---|---|---|---|---|---|
| β-Galactosidase, Bga2A | Bpr_I0279 | 1.5 × 10−12 | N | 4.7 | 118.7 | 8 |
| β-Mannosidase, Man2A | Bpr_III237 | 3.1 × 10−11 | N | 4.8 | 95.9 | 7 |
| β-Glucosidase, Bgl3A | Bpr_I0693 | 2.5 × 10−6 | Y | 4.1 | 115.6 | 23 |
| β-Glucosidase, Bgl3B | Bpr_I0847 | 2.6 × 10−11 | N | 4.7 | 103.7 | 7 |
| β-Glucosidase, Bgl3C | Bpr_I0138 | 8.1 × 10−7 | N | 4.9 | 91.5 | 2 |
| β-Xylosidase Xyl3A | Bpr_I0184 | 8.9 × 10−15 | N | 4.8 | 78.2 | 42 |
| Cellodextrinase, Cel9B | Bpr_I1593 | 1.0 × 10−30 | N | 4.6 | 61.0 | 22 |
| Endo-1,4-β-xylanase and esterase, Xyn10D | Bpr_I1083 | 3.6 × 10−10 | N | 5.0 | 79.7 | 5 |
| α-Amylase, Amy13G | Bpr_I0729 | 1.7 × 10−11 | N | 4.7 | 60.3 | 3 |
| Pullulanase, Pul13A | Bpr_III161 | 1.1 × 10−10 | Y | 4.4 | 99.7 | 7 |
| Glycoside hydrolase family 31, GH31C | Bpr_I1974 | 1.0 × 10-11 | N | 4.7 | 78.2 | 32 |
| β-Galactosidase, Bga35B | Bpr_I2006 | 3.1 × 10-10 | N | 4.9 | 83.2 | 4 |
| α-Galactosidase, Aga36C | Bpr_III065 | 9.3 × 10−5 | N | 5.1 | 83.2 | 7 |
| Xylosidase/arabinofuranosidase and esterase, Xsa43H | Bpr_I0301 | 1.6 × 10−8 | N | 4.7 | 107.9 | 7 |
| α-l-Arabinofuranosidase, Arf51A | Bpr_I0329 | 1.1 × 10−15 | N | 5.2 | 57.0 | 14 |
| α-d-Glucuronidase, Agu67A | Bpr_I0177 | 4.9 × 10−11 | N | 4.9 | 76.1 | 22 |
| α-l-Rhamnosidase, Rha78A | Bpr_I1686 | 99.9 | N | 4.8 | 85.0 | 3 |
| Cellobiose phosphorylase, Cbp94A | Bpr_I2447 | 1.1 × 10−16 | N | 5.1 | 91.5 | 6 |
| Feruloyl esterase, Est1E | Bpr_I2870 | 1.3 × 10−10 | N | 5.2 | 27.8 | 5 |
| Glycogen phosphorylase, Glgp2 | Bpr_I2847 | 1.8 × 10−10 | N | 6.0 | 94.8 | 10 |
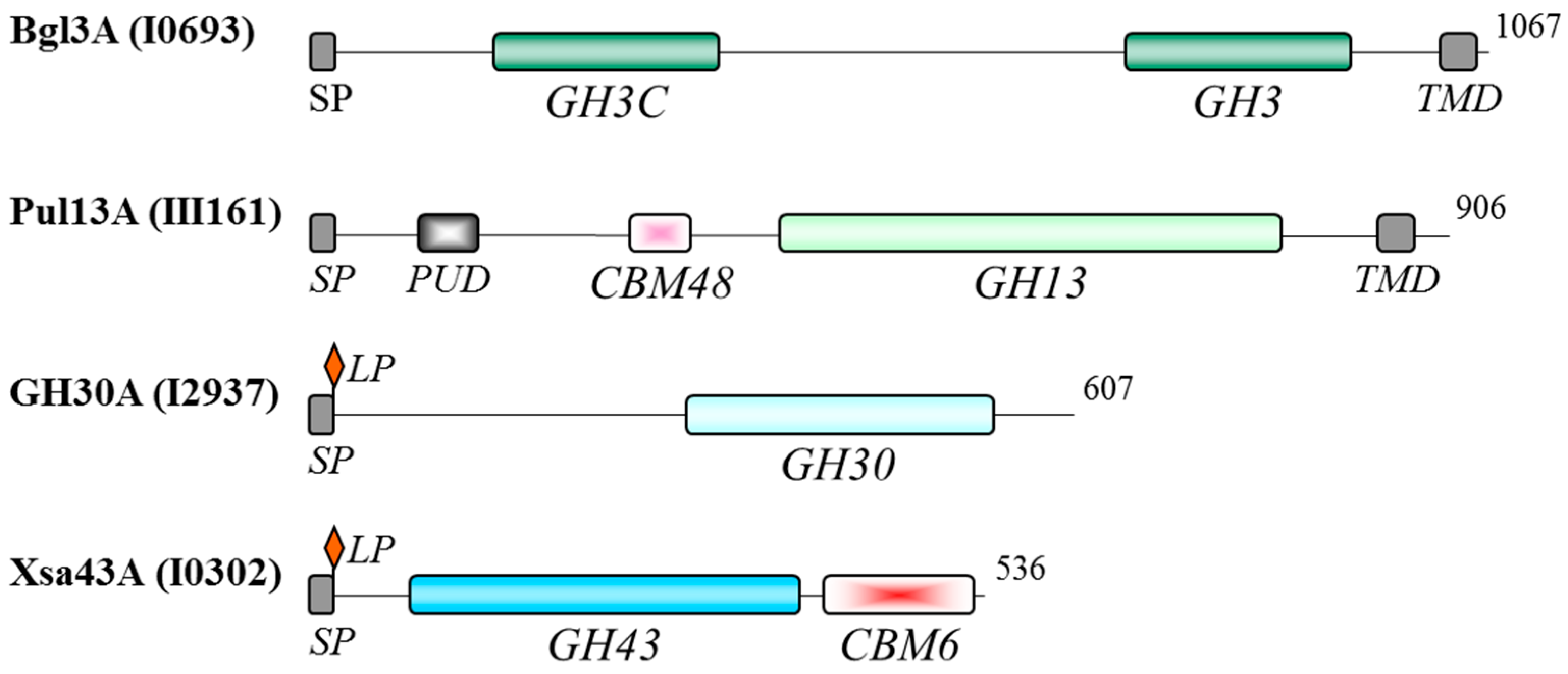
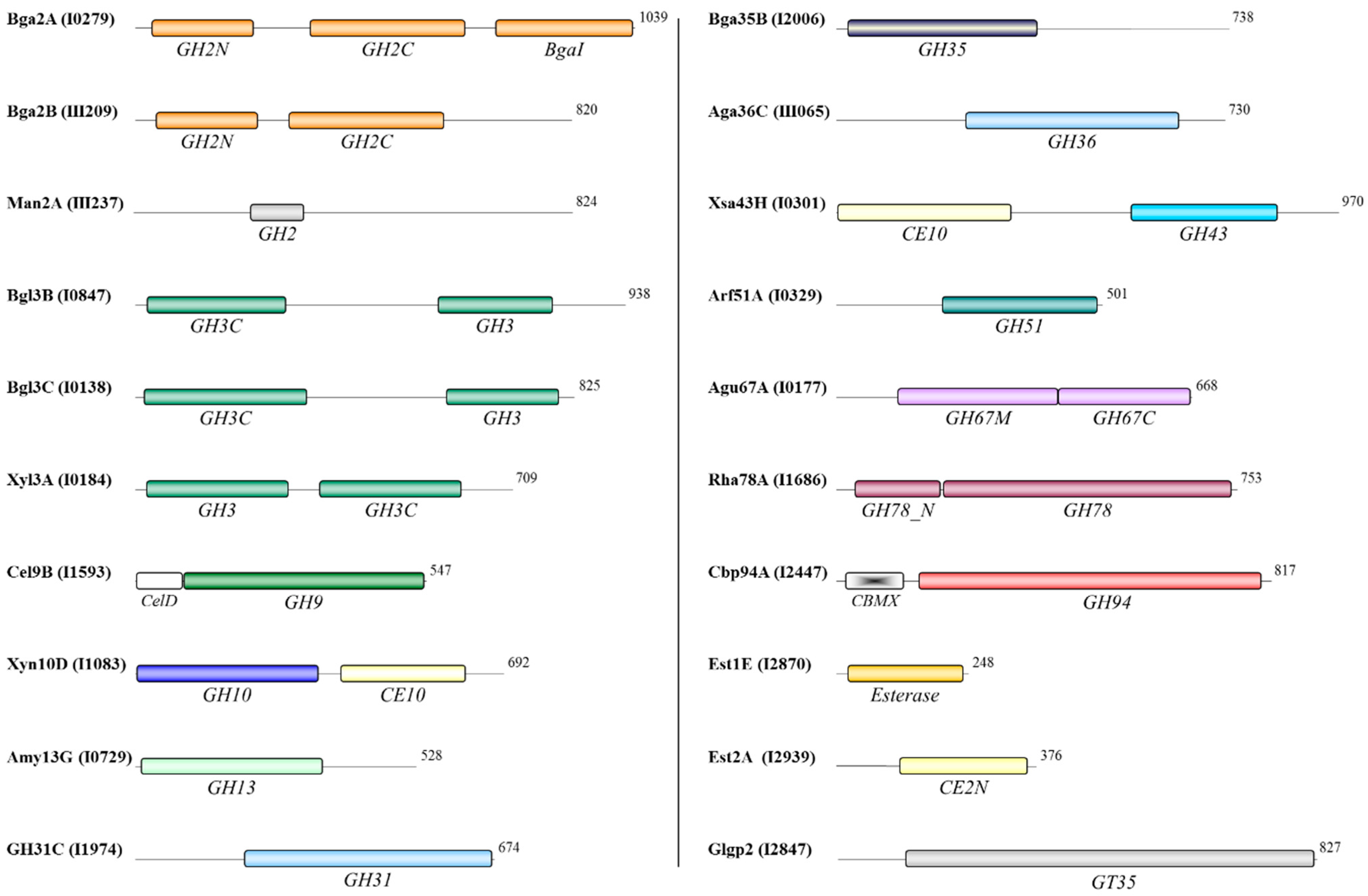
3.4. Functional Domains of the Oligosaccharidases
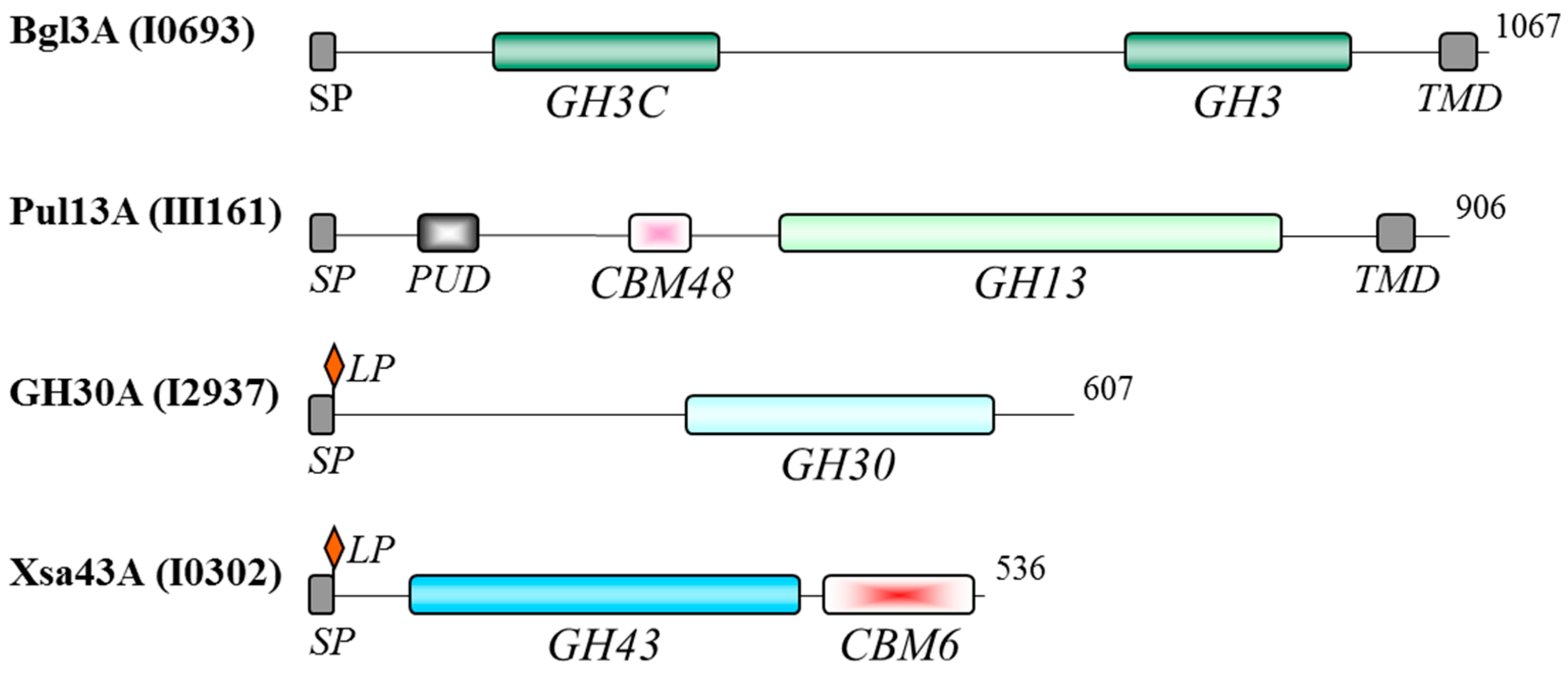
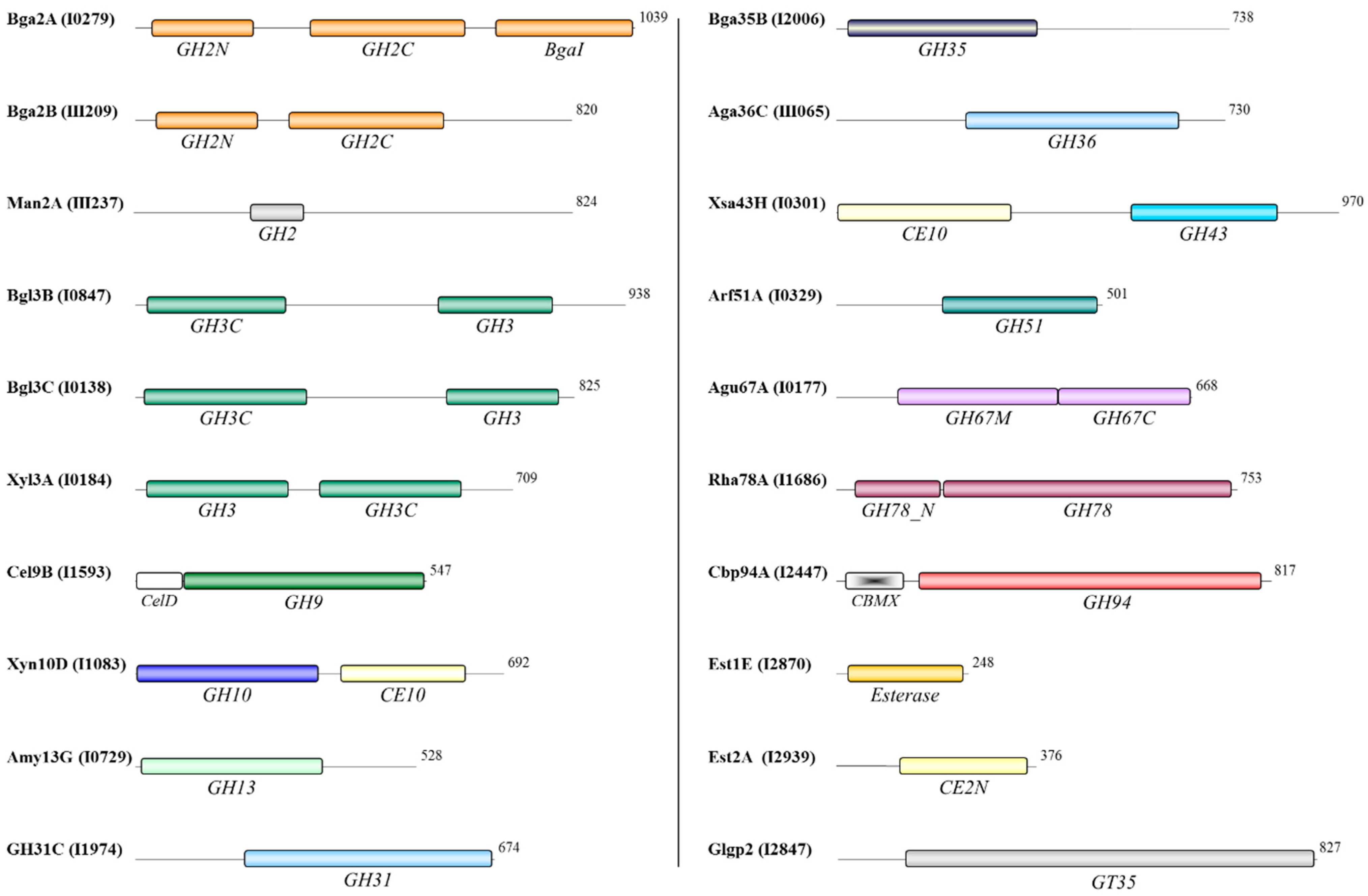
3.5. Effects of Growth on Xylan or Xylose
| Spot | Protein | Locus | Function i | Expect. ii | pI | kDa | Fold-Change iii | p Value |
|---|---|---|---|---|---|---|---|---|
| C853 | Adenylosuccinate lyase, PurB | Bpr_I2212 | O | 1.2 × 10−10 | 5.4 | 53.7 | −3.1 ± 0.6 | 0.001 |
| C1135 | Amino acid ABC transporter substrate-binding protein | Bpr_I1826 | T | 5.8 × 10−4 | 4.0 | 31.6 | Xylan | n/a |
| C1010 | Anti-sigma factor antagonist/phosphotransferase domain-containing protein | Bpr_I0249 | R | 3.0 × 10−30 | 4.6 | 49.8 | −12.0 ± 0.7 | 0.001 |
| Ribosomal protein S1, RpsA | Bpr_I2035 | Q | 4.6 | 41.9 | ||||
| C654 | DNA-directed RNA polymerase α subunit, RpoA | Bpr_I0623 | W | 3.0 × 10−17 | 4.4 | 35.1 | −8.7 ± 4.3 | 0.002 |
| C837 | Fructose-1,6-bisphosphate aldolase, FbaA | Bpr_I2903 | C | 3.0 × 10−13 | 5.1 | 30.5 | −2.1 ± 0.2 | 0.002 |
| C733 | IMP cyclohydrolase, PurO | Bpr_I0731 | O | 1.5 × 10−18 | 4.7 | 32.2 | 2.3 ± 0.5 | 0.001 |
| Translation elongation factor Tu, TufA | Bpr_I2364 | Q | 4.82 | 43.6 | ||||
| C792 | NADPH-dependent glutamate synthase, GltA3 | Bpr_I1306 | A | 1.9 × 10−4 | 5.0 | 49.3 | −15.8 ± 6.3 | 0.001 |
| C637 | Pullulanase, Pul13A | Bpr_III161 | C | 1.5 × 10−9 | 4.4 | 99.7 | 2.8 ± 0.3 | 0.001 |
| C638 | Pullulanase, Pul13A | Bpr_III161 | C | 1.8 × 10−6 | 4.4 | 99.7 | Xylan | n/a |
| C709 | Ribosomal protein S1, RpsA | Bpr_I2035 | Q | 9.6 × 10−19 | 4.6 | 41.9 | −4.1 ± 1.1 | 0.001 |
| C1072 | Serine protease subtilisin family | Bpr_I2629 | P | 3.8 × 10−8 | 3.8 | 153.3 | 5.8 ± 1.3 | 0.001 |
| C1074 | Sugar ABC transporter substrate-binding protein | Bpr_I0182 | T | 1.5 × 10−8 | 4.12 | 63.3 | Xylan | n/a |
| Spot | Protein | Locus | Function i | Expect. ii | pI | kDa | Fold-Change iii | p Value |
|---|---|---|---|---|---|---|---|---|
| C1135 | Amino acid ABC transporter substrate-binding protein | Bpr_I1826 | T | 5.8 × 10−4 | 4.0 | 31.6 | Xylan | n/a |
| C785 | Aminotransferase DegT/DnrJ/EryC1/StrS family | Bpr_I2311 | I | 1.5 × 10−10 | 5.0 | 51.0 | 4.2 ± 1.1 | 0.001 |
| Xylulokinase, XylB | Bpr_I0173 | C | 4.9 | 53.7 | ||||
| C1120 | β-Galactosidase, Bga35B | Bpr_I2006 | C | 1.9 × 10−27 | 4.9 | 83.2 | 4.5 ± 0.8 | 0.001 |
| β-Xylosidase, Xyl3A | Bpr_I0184 | C | 4.8 | 78.2 | ||||
| C704 | Hypothetical protein | Bpr_I2455 | H | 7.6 × 10−22 | 4.7 | 47.7 | 2.3 ± 0.1 | 0.001 |
| Phosphoribosylamine-glycine ligase, PurD | Bpr_I0870 | O | 4.7 | 46.3 | ||||
| C601 | Oligopeptide ABC transporter substrate-binding protein, OppA1 | Bpr_I1276 | T | 1.9 × 10−13 | 4.0 | 83.1 | 2.3 ± 0.5 | 0.001 |
| C784 | Peptidase U62 family | Bpr_I2456 | P | 6.1 × 10−9 | 4.8 | 52.3 | 14.7 ± 4.6 | 0.001 |
| C1074 | Sugar ABC transporter substrate-binding protein | Bpr_I0182 | T | 7.6 × 10−7 | 4.12 | 63.3 | Xylan | n/a |
| C1057 | Xylose ABC transporter substrate-binding protein | Bpr_I1173 | T | 9.6 × 10−12 | 4.2 | 38.4 | −3.9 ± 1.1 | 0.001 |
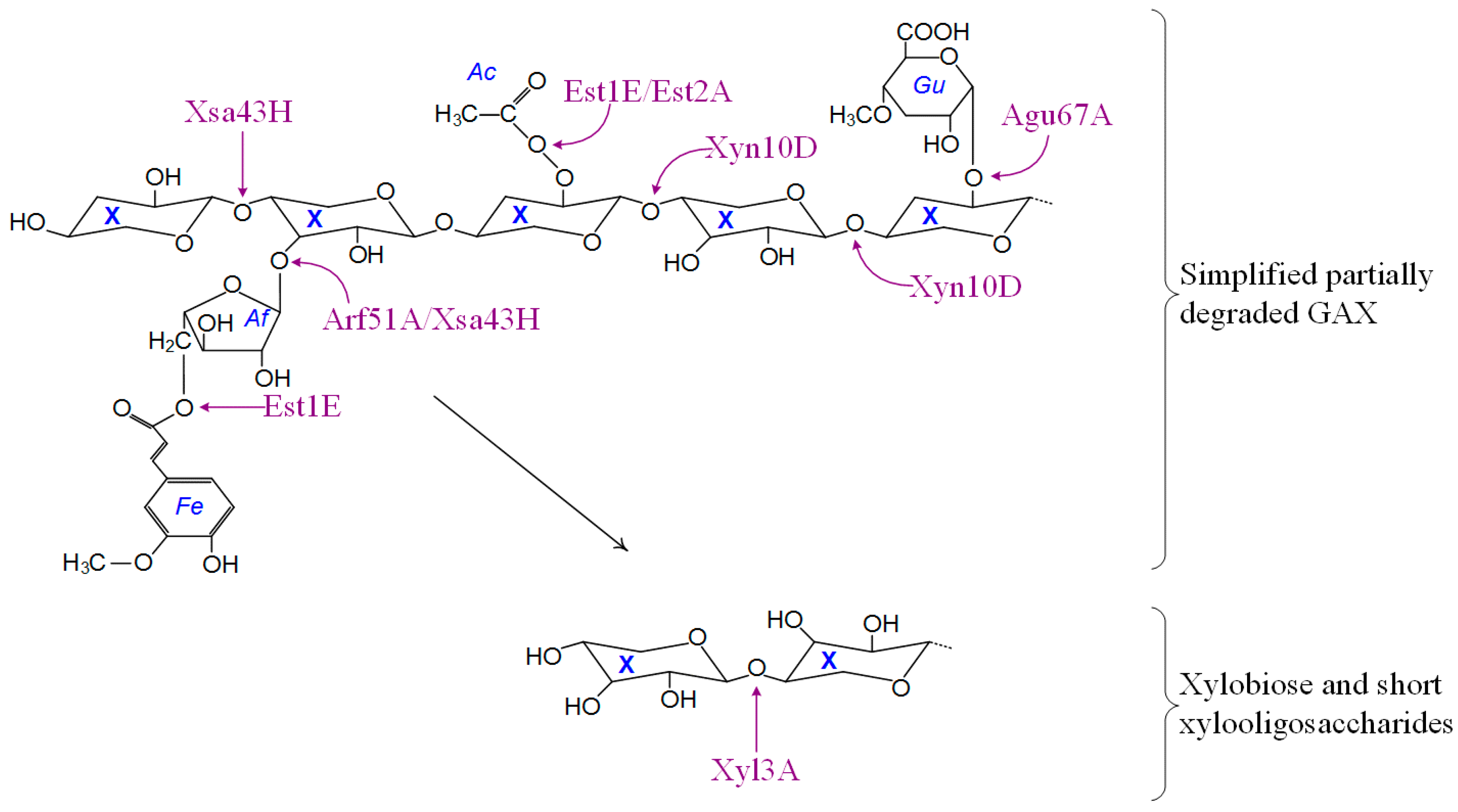
3.6. Predicted Activities and Pathways
| Protein | Locus | Substrate | Reaction Catalysed i |
|---|---|---|---|
| β-Galactosidase, Bga2A β-Galactosidase, Bga2B β-Galactosidase, Bga35B | Bpr_I0279 Bpr_III209 Bpr_I2006 | β-d-Galactosides | Hydrolysis of terminal, non-reducing β-d-galactose residues. |
| β-Mannosidase, Man2A | Bpr_III237 | β-d-Mannosides | Hydrolysis of terminal, non-reducing β-d-mannose residues. |
| β-Glucosidase, Bgl3B β-Glucosidase, Bgl3C | Bpr_I0847 Bpr_I0138 | β-d-Glucosides | Hydrolysis of terminal, non-reducing β-d-glucosyl residues with release of β-d-glucose. |
| β-Xylosidase, Xyl3A | Bpr_I0184 | 1,4-β-d-Xylans | Hydrolysis of terminal, non-reducing d-xylose residues. |
| Cellodextrinase, Cel9B | Bpr_I1593 | Cellulose, lichenin and cereal β-d-glucans | Endohydrolysis of (1→4)-β-d-glucosidic linkages. |
| Endo-1,4-β-xylanase and esterase, Xyn10D | Bpr_I1083 | 1,4-β-d-Xylans | Endohydrolysis of (1→4)-β-d-xylosidic linkages. |
| α-Amylase, Amy13G | Bpr_I0729 | Starch and glycogen | Endohydrolysis of (1→6)-α-d-glucosidic linkages. |
| Glycoside hydrolase family 31, GH31C | Bpr_I1974 | Unknown | Unknown. |
| α-Galactosidase, Aga36C | Bpr_III065 | α-d-Galactosides | Hydrolysis of terminal, non-reducing α-d-galactose residues. |
| Xylosidase/arabinofuranosidase and esterase, Xsa43H | Bpr_I0301 | α-l-Arabinosides and triacylglycerols | Hydrolysis of terminal, non-reducing D-xylose or α-l-arabinofuranoside residues/hydrolysis of triacylglycerols with release of a diacylglycerol and a carboxylate. |
| α-L-Arabinofuranosidase, Arf51A | Bpr_I0329 | α-l-Arabinofuranosides | Hydrolysis of terminal, non-reducing α-l-arabinofuranoside residues. |
| α-D-Glucuronidase, Agu67A | Bpr_I0177 | Glucuronoxylans | Hydrolysis of glucuronic acid substituted xylooligosaccharides. |
| α-L-Rhamnosidase, Rha78A | Bpr_I1686 | α-l-Rhamnosides | Hydrolysis of terminal, non-reducing α-l-rhamnose residues. |
| Cellobiose phosphorylase, Cbp94A | Bpr_I2447 | Cellobiose | Hydrolysis of cellobiose. |
| Feruloyl esterase, Est1E | Bpr_I2870 | Esterified oligosaccharides | Deferuloylation of esterified oligosaccharides. |
| Acetyl-xylan esterase, Est2A | Bpr_I2939 | Acetylated xylans and xylo-oligosaccharides | Deacetylation of xylans and xylo-oligosaccharides. |
| Glycogen phosphorylase, Glgp2 | Bpr_I2847 | 1,4-α-d-Glucans | Exohydrolysis and phosphorylation of (1→4)-α-d-glucan. |
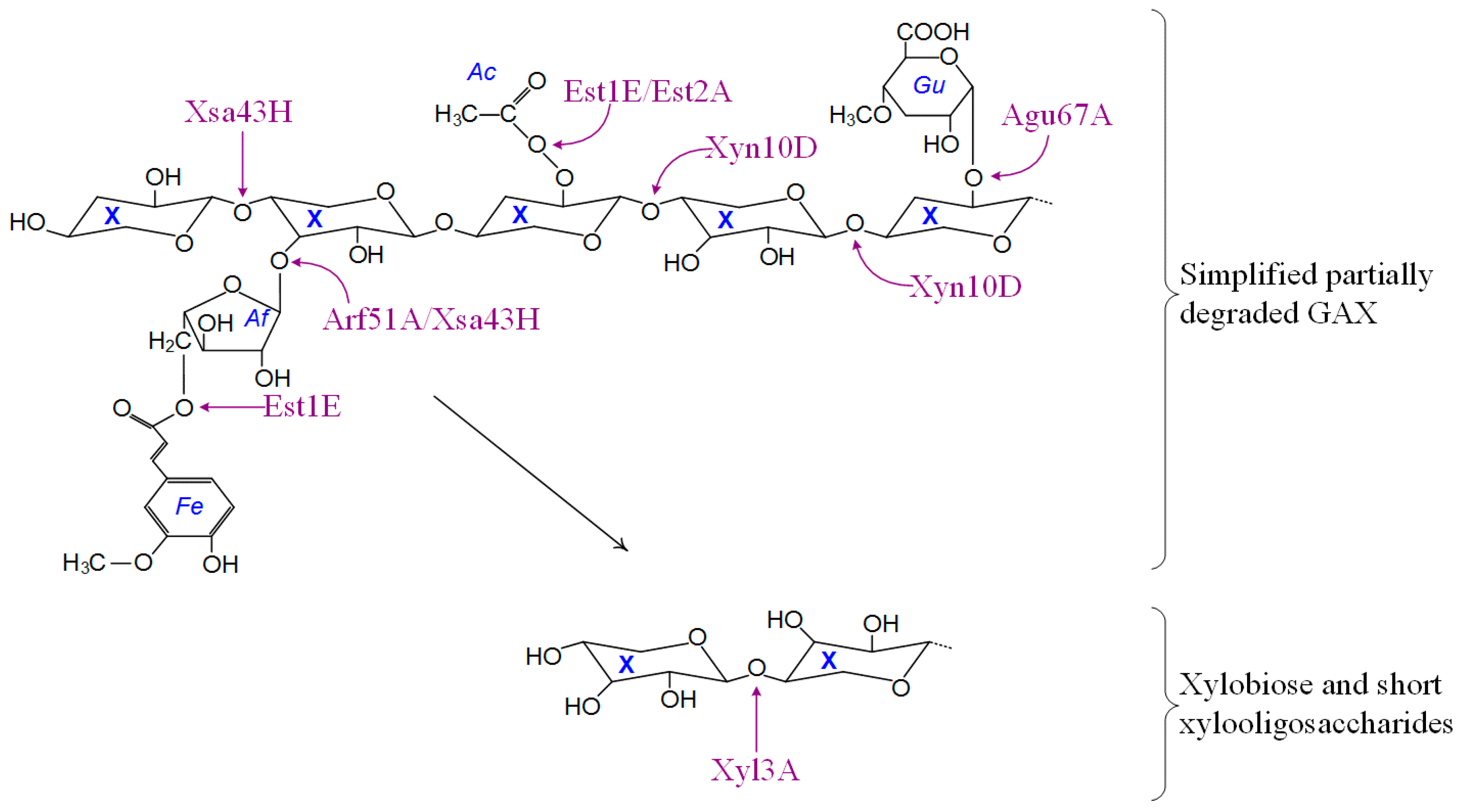
4. Conclusions
Supplementary Materials
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kulkarni, N.; Shendye, A.; Rao, M. Molecular and biotechnological aspects of xylanases. FEMS Microbiol. Rev. 1999, 23, 411–456. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The Carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed]
- Attwood, G.T.; Reilly, K.; Patel, B.K.C. Clostridium proteoclasticum sp nov, a novel proteolytic bacterium from the bovine rumen. Int. J. Syst. Bacteriol. 1996, 46, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Moon, C.D.; Pacheco, D.M.; Kelly, W.J.; Leahy, S.C.; Li, D.; Kopecny, J.; Attwood, G.T. Reclassification of Clostridium proteoclasticum as Butyrivibrio proteoclasticus comb. nov., a butyrate producing ruminal bacterium. Int. J. Syst. Evol. Microbiol. 2008, 58, 2041–2045. [Google Scholar] [CrossRef] [PubMed]
- Attwood, G.T.; Reilly, K. Identification of proteolytic rumen bacteria isolated from New Zealand cattle. J. Appl. Bacteriol. 1995, 79, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Paillard, D.; McKain, N.; Rincon, M.T.; Shingfield, K.J.; Givens, D.I.; Wallace, R.J. Quantification of ruminal Clostridium proteoclasticum by real-time PCR using a molecular beacon approach. J. Appl. Microbiol. 2007, 103, 1251–1261. [Google Scholar] [CrossRef] [PubMed]
- Reilly, K.; Attwood, G.T. Detection of Clostridium proteoclasticum and closely related strains in the rumen by competitive PCR. Appl. Environ. Microbiol. 1998, 64, 907–913. [Google Scholar] [PubMed]
- Edwards, J.E.; McEwan, N.R.; Travis, A.J.; Wallace, R.J. 16S rDNA library-based analysis of ruminal bacterial diversity. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2004, 86, 263–281. [Google Scholar] [CrossRef] [PubMed]
- Kelly, W.J.; Leahy, S.; Altermann, E.; Yeoman, C.J.; Dunne, J.C.; Kong, Z.; Pacheco, D.M.; Li, D.; Noel, S.; Moon, C.D.; et al. The glycobiome of the rumen bacterium Butyrivibrio proteoclasticus B316T highlights adaptation to a polysaccharide-rich environment. PLoS ONE 2010, 8, e11942. [Google Scholar] [CrossRef] [PubMed]
- Dunne, J.C.; Li, D.; Kelly, W.J.; Leahy, S.C.; Bond, J.J.; Attwood, G.T.; Jordan, T.W. Extracellular polysaccharide-degrading proteome of Butyrivibrio proteoclasticus. J. Proteome Res. 2012, 11, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Biely, P.; Vrsanska, M.; Tenkanen, M.; Kluepfel, D. Endo-β-1,4-xylanase families: Differences in catalytic properties. J. Bacteriol. 1997, 57, 151–166. [Google Scholar] [CrossRef]
- Taguchi, H.; Koike, S.; Kobayashi, Y.; Cann, I.K.O.; Karita, S. Partial characterization of structure and function of a xylanase gene from the rumen hemicellulolytic bacterium Eubacterium ruminantium. Anim. Sci. J. 2004, 75, 325–332. [Google Scholar] [CrossRef]
- Bond, J.J.; Dunne, J.C.; Kwan, F.Y.S.; Li, D.; Zhang, K.K.; Leahy, S.C.; Kelly, W.J.; Attwood, G.T.; Jordan, T.W. Carbohydrate transporting membrane proteins of the rumen bacterium, Butyrivibrio proteoclasticus. J. Proteomics 2012, 75, 3138–3144. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, S.C.; Witters, E.; Laukens, K.; Deckers, P.; Swennen, R.; Panis, B. Preparation of protein extracts from recalcitrant plant tissues: An evaluation of different methods for two-dimensional gel electrophoresis analysis. Proteomics 2005, 5, 2497–2507. [Google Scholar] [CrossRef] [PubMed]
- Rawson, P.; Stockum, C.; Peng, L.; Manivannan, B.; Lehnert, K.; Ward, H.E.; Berry, S.D.; Davis, S.R.; Snell, R.G.; McLauchlan, D.; et al. Metabolic proteomics of the liver and mammary gland during lactation. J. Proteomics 2012, 75, 4429–4435. [Google Scholar] [CrossRef] [PubMed]
- Smit, A.M.; Strabala, T.J.; Peng, L.; Rawson, P.; Lloyd-Jones, G.; Jordan, T.W. Proteomic phenotyping of Novosphingobium nitrogenifigens reveals a robust capacity for simultaneous nitrogen fixation, polyhydroxyalkanoate production, and resistance to reactive oxygen species. Appl. Environ. Microbiol. 2012, 78, 4802–4815. [Google Scholar] [CrossRef] [PubMed]
- MASCOT. Available online: http://www.matrixscience.com/ (accessed on 23 Ocotber 2015).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Finn, R.D.; Tate, J.; Mistry, J.; Coggill, P.C.; Sammut, S.J.; Hotz, H.R.; Ceric, G.; Forslund, K.; Eddy, S.R.; Sonnhammer, E.L.L.; et al. The Pfam protein families database. Nucleic Acids Res. 2008, 36, D281–D288. [Google Scholar] [CrossRef] [PubMed]
- Haft, D.H.; Selengut, J.D.; White, O. The TIGRFAMs database of protein families. Nucleic Acids Res. 2003, 31, 371–373. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, J.D.; Nielsen, H.; von Heijne, G.; Brunak, S. Improved prediction of signal peptides: SignalP 3.0. J. Mol. Biol. 2004, 340, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Juncker, A.S.; Willenbrock, H.; von Heijne, G.; Brunak, S.; Nielsen, H.; Krogh, A. Prediction of lipoprotein signal peptides in Gram-negative bacteria. Protein Sci. 2003, 12, 1652–1662. [Google Scholar] [CrossRef] [PubMed]
- Sutcliffe, I.C.; Harrington, D.J. Pattern searches for the identification of putative lipoprotein genes in Gram-positive bacterial genomes. Microbiology-SGM 2002, 148, 2065–2077. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Hirokawa, T.; Boon-Chieng, S.; Mitaku, S. SOSUI: Classification and secondary structure prediction system for membrane proteins. Bioinformatics 1998, 14, 378–379. [Google Scholar] [CrossRef] [PubMed]
- Machovic, M.; Janecek, S. Domain evolution in the GH13 pullulanase subfamily with focus on the carbohydrate-binding module family 48. Biologia 2008, 63, 1057–1068. [Google Scholar] [CrossRef]
- Correia, M.A.S.; Pires, V.M.R.; Gilbert, H.J.; Bolam, D.N.; Fernandes, V.O.; Alves, V.D.; Prates, J.A.M.; Ferreira, L.M.A.; Fontes, C. Family 6 carbohydrate-binding modules display multiple β-1,3-linked glucan-specific binding interfaces. FEMS Microbiol. Lett. 2009, 300, 48–57. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Henshaw, J.L.; Bolam, D.N.; Pires, V.M.R.; Czjzek, M.; Henrissat, B.; Ferreira, L.M.A.; Fontes, C.; Gilbert, H.J. The Family 6 carbohydrate binding module CmCBM6–2 contains two ligand-binding sites with distinct specificities. J. Biol. Chem. 2004, 279, 21552–21559. [Google Scholar] [CrossRef] [PubMed]
- Bourgois, T.M.; van Craeyveld, V.; van Campenhout, S.; Courtin, C.M.; Delcour, J.A.; Robben, J.; Volckaert, G. Recombinant expression and characterization of XynD from Bacillus subtilis subsp. subtilis ATCC 6051: A GH 43 arabinoxylan arabinofuranohydrolase. Appl. Microbiol. Biotechnol. 2007, 75, 1309–1317. [Google Scholar] [PubMed]
- Nolling, J.; Breton, G.; Omelchenko, M.V.; Makarova, K.S.; Zeng, Q.D.; Gibson, R.; Lee, H.M.; Dubois, J.; Qiu, D.Y.; Hitti, J.; et al. Genome sequence and comparative analysis of the solvent-producing bacterium Clostridium acetobutylicum. J. Bacteriol. 2001, 183, 4823–4838. [Google Scholar] [CrossRef] [PubMed]
- Borneman, W.S.; Hartley, R.D.; Himmelsbach, D.S.; Ljungdahl, L.G. Assay for trans-para-coumaroyl esterase using a specific substrate from plant-cell walls. Anal. Biochem. 1990, 190, 129–133. [Google Scholar] [CrossRef]
- Hartley, R.D.; Morrison, W.H.; Himmelsbach, D.S.; Borneman, W.S. Cross-linking of cell-wall phenolic arabinoxylans in gramineous plants. Phytochemistry 1990, 29, 3705–3709. [Google Scholar] [CrossRef]
- Mueller-Harvey, I.; Hartley, R.D.; Harris, P.J.; Curzon, E.H. Linkage of para-coumaroyl and feruloyl groups to cell-wall polysaccharides of barley straw. Carbohydr. Res. 1986, 148, 71–85. [Google Scholar] [CrossRef]
- Vietor, R.J.; Angelino, S.; Voragen, A.G.J. Structural features of arabinoxylans from barley and malt cell-wall material. J. Cereal Sci. 1992, 15, 213–222. [Google Scholar] [CrossRef]
- Till, M.; Goldstone, D.C.; Attwood, G.T.; Moon, C.D.; Kelly, W.J.; Arcus, V.L. Structure and function of an acetyl xylan esterase (Est2A) from the rumen bacterium Butyrivibrio proteoclasticus. Proteins-Struct. Funct. Bioinform. 2013, 81, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Goldstone, D.C.; Villas-Boas, S.G.; Till, M.; Kelly, W.J.; Attwood, G.T.; Arcus, V.L. Structural and functional characterization of a promiscuous feruloyl esterase (Est1E) from the rumen bacterium Butyrivibrio proteoclasticus. Proteins 2010, 78, 1457–1469. [Google Scholar] [PubMed]
- Schwarz, W.H.; Bronnenmeier, K.; Krause, B.; Lottspeich, F.; Staudenbauer, W.L. Debranching of arabinoxylan - properties of the thermoactive recombinant α-l-arabinofuranosidase from Clostridium-stercorarium (Arfb). Appl. Microbiol. Biotechnol. 1995, 43, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Zverlov, V.V.; Liebl, W.; Bachleitner, M.; Schwarz, W.H. Nucleotide sequence of arfB of Clostridium stercorarium, and prediction of catalytic residues of α-l-arabinofuranosidases based on local similarity with several families of glycosyl hydrolases. FEMS Microbiol. Lett. 1998, 164, 337–343. [Google Scholar] [CrossRef]
- Kosugi, A.; Murashima, K.; Doi, R.H. Characterization of two noncellulosomal subunits, ArfA and BgaA, from Clostridium cellulovorans that cooperate with the cellulosome in plant cell wall degradation. J. Bacteriol. 2002, 184, 6859–6865. [Google Scholar] [CrossRef] [PubMed]
- Golan, G.; Shallom, D.; Teplitsky, A.; Zaide, G.; Shulami, S.; Baasov, T.; Stojanoff, V.; Thompson, A.; Shoham, Y.; Shoham, G. Crystal structures of Geobacillus stearothermophilus α-glucuronidase complexed with its substrate and products—Mechanistic implications. J. Biol. Chem. 2004, 279, 3014–3024. [Google Scholar] [CrossRef] [PubMed]
- Chow, V.; Nong, G.; Preston, J.F. Structure, Function, and Regulation of the Aldouronate Utilization Gene Cluster from Paenibacillus sp. Strain JDR-2. J. Bacteriol. 2007, 189, 8863–8870. [Google Scholar] [CrossRef] [PubMed]
- Shallom, D.; Golan, G.; Shoham, G.; Shoham, Y. Effect of dimer dissociation on activity and thermostability of the α-glucuronidase from Geobacillus stearothermophilus: Dissecting the different oligomeric forms of Family 67 glycoside hydrolases. J. Bacteriol. 2004, 186, 6928–6937. [Google Scholar] [CrossRef] [PubMed]
- Sakka, K.; Yoshikawa, K.; Kojima, Y.; Karita, S.; Ohmiya, K.; Shimada, K. Nucleotide-sequence of the Clostridium stercorarium xylA gene encoding a bifunctional protein with β-d-xylosidase and α-l-arabinofuranosidase activities, and properties of the translated product. Biosci. Biotechnol. Biochem. 1993, 57, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.L.; Thomson, J.A. Cloning, sequencing and expression of a gene encoding a 73 kDa xylanase enzyme from the rumen anaerobe Butyrivibrio fibrisolvens H17c. Mol. Gen. Genet. 1991, 228, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, K.A.; Colombato, D.; Morgavi, D.P.; Yang, W.Z. Use of exogenous fibrolytic enzymes to improve feed utilization by ruminants. J. Anim. Sci. 2003, 81, E37–E47. [Google Scholar]
- Yang, H.J.; Xie, C.Y. Assessment of fibrolytic activities of 18 commercial enzyme products and their abilities to degrade the cell wall fraction of corn stalks in in vitro enzymatic and ruminal batch cultures. Anim. Feed Sci. Technol. 2010, 159, 110–121. [Google Scholar] [CrossRef]
- Phakachoed, N.; Suksombat, W.; Colombatto, D.; Beauchemin, K.A. Use of fibrolytic enzymes additives to enhance in vitro ruminal fermentation of corn silage. Livest. Sci. 2013, 157, 100–112. [Google Scholar] [CrossRef]
- Schingoethe, D.J.; Stegeman, G.A.; Treacher, R.J. Response of lactating dairy cows to a cellulase and xylanase enzyme mixture applied to forages at the time of feeding. J. Dairy Sci. 1999, 82, 996–1003. [Google Scholar] [CrossRef]
- Kung, L.; Treacher, R.J.; Nauman, G.A.; Smagala, A.M.; Endres, K.M.; Cohen, M.A. The effect of treating forages with fibrolytic enzymes on its nutritive value and lactation performance of dairy cows. J. Dairy Sci. 2000, 83, 115–122. [Google Scholar] [CrossRef]
- Yang, W.Z.; Beauchemin, K.A.; Rode, L.M. A comparison of methods of adding fibrolytic enzymes to lactating cow diets. J. Dairy Sci. 2000, 83, 2512–2520. [Google Scholar] [CrossRef]
- Cruywagen, C.W.; van Zyl, W.H. Effects of a fungal enzyme cocktail treatment of high and low forage diets on lamb growth. Anim. Feed Sci. Technol. 2008, 145, 151–158. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Rode, L.M.; Sewalt, V.J.H. Fibrolytic enzymes increase fiber digestibility and growth rate of steers fed dry forages. Can. J. Anim. Sci. 1995, 75, 641–644. [Google Scholar] [CrossRef]
- Titi, H.; Lubbadeh, W.F. Effect of feeding cellulase enzyme on productive responses of pregnant and lactating ewes and goats. Small Rumin. Res. 2004, 52, 137–143. [Google Scholar] [CrossRef]
- Pinos-Rodriguez, J.M.; Gonzalez, S.S.; Mendoza, G.D.; Barcena, R.; Cobos, M.A.; Hernandez, A.; Ortega, M.E. Effect of exogenous fibrolytic enzyme on ruminal fermentation and digestibility of alfalfa and rye-grass hay fed to lambs. J. Anim. Sci. 2002, 80, 3016–3020. [Google Scholar] [PubMed]
- ZoBell, D.R.; Wiedmeier, R.D.; Olson, K.C.; Treacher, R. The effect of an exogenous enzyme treatment on production and carcass characteristics of growing and finishing steers. Anim. Feed Sci. Technol. 2000, 87, 279–285. [Google Scholar] [CrossRef]
- Peters, A.; Lebzien, P.; Meyer, U.; Borchert, U.; Bulang, M.; Flachowsky, G. Effect of exogenous fibrolytic enzymes on ruminal fermentation and nutrient digestion in dairy cows. Arch. Anim. Nutr. 2010, 64, 221–237. [Google Scholar] [CrossRef] [PubMed]
- Aboagye, I.A.; Lynch, J.P.; Church, J.S.; Baah, J.; Beauchemin, K.A. Digestibility and growth performance of sheep fed alfalfa hay treated with fibrolytic enzymes and a ferulic acid esterase producing bacterial additive. Anim. Feed Sci. Technol. 2015, 203, 53–66. [Google Scholar] [CrossRef]
- Torres, N.; Mendoza, G.D.; Barcena, R.; Loera, O.; Gonzalez, S.; Aranda, E.; Hernandez, P.A.; Crosby, M. Effects of various fibrolytic enzyme extracts on digestibility and productive performance of lambs fed a forage-based diet. Anim. Nutr. Feed Technol. 2013, 13, 381–389. [Google Scholar]
- Colombatto, D.; Morgavi, D.P.; Furtado, A.F.; Beauchemin, K.A. Screening of exogenous enzymes for ruminant diets: Relationship between biochemical characteristics and in vitro ruminal degradation. J. Anim. Sci. 2003, 81, 2628–2638. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dunne, J.C.; Kelly, W.J.; Leahy, S.C.; Li, D.; Bond, J.J.; Peng, L.; Attwood, G.T.; Jordan, T.W. The Cytosolic Oligosaccharide-Degrading Proteome of Butyrivibrio Proteoclasticus. Proteomes 2015, 3, 347-368. https://doi.org/10.3390/proteomes3040347
Dunne JC, Kelly WJ, Leahy SC, Li D, Bond JJ, Peng L, Attwood GT, Jordan TW. The Cytosolic Oligosaccharide-Degrading Proteome of Butyrivibrio Proteoclasticus. Proteomes. 2015; 3(4):347-368. https://doi.org/10.3390/proteomes3040347
Chicago/Turabian StyleDunne, Jonathan C., William J. Kelly, Sinead C. Leahy, Dong Li, Judy J. Bond, Lifeng Peng, Graeme T. Attwood, and T. William Jordan. 2015. "The Cytosolic Oligosaccharide-Degrading Proteome of Butyrivibrio Proteoclasticus" Proteomes 3, no. 4: 347-368. https://doi.org/10.3390/proteomes3040347
APA StyleDunne, J. C., Kelly, W. J., Leahy, S. C., Li, D., Bond, J. J., Peng, L., Attwood, G. T., & Jordan, T. W. (2015). The Cytosolic Oligosaccharide-Degrading Proteome of Butyrivibrio Proteoclasticus. Proteomes, 3(4), 347-368. https://doi.org/10.3390/proteomes3040347