
Enzymatic Investigation of Spongospora subterranea Zoospore Attachment to Roots of Potato Cultivars Resistant or Susceptible to Powdery Scab Disease

 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. S. subterranea sporosori Collection and Germination
2.2. Plant Materials and Growth Conditions
2.3. Enzyme Treatments, Including the Trypsin Shaving Time-Course Study
2.4. Spongospora subterranea Zoospore Root-Attachment Assay
2.5. Proteomic Analysis and Data Processing
2.6. Bioinformatics and Statistical Analysis
3. Results
3.1. Effects of Enzyme Treatments on Zoospore Root Attachment
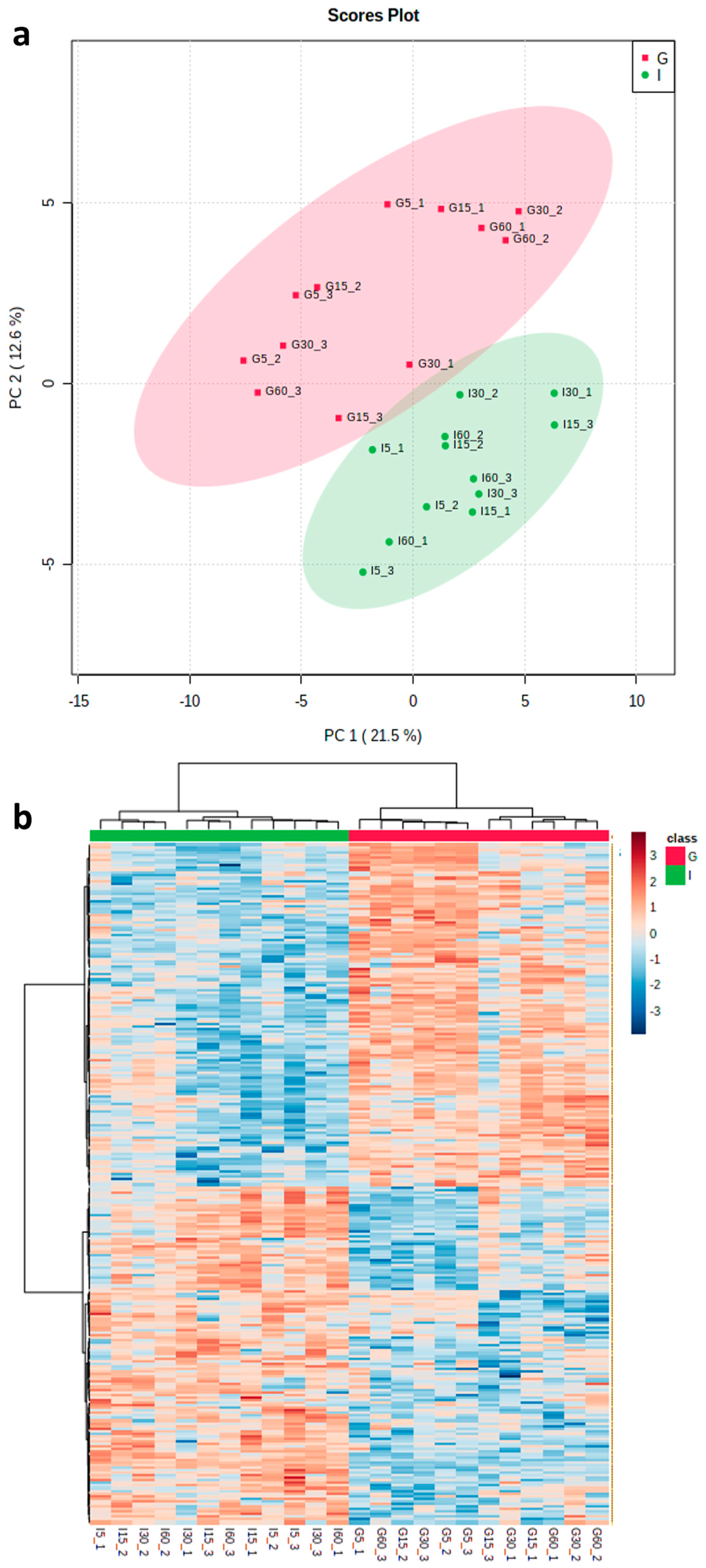
3.2. Analysis of Proteins Released by Trypsin Shaving Treatment of Potato Roots
3.3. Overall Functional Classification and Pathway Analysis of Differentially Abundant Proteins
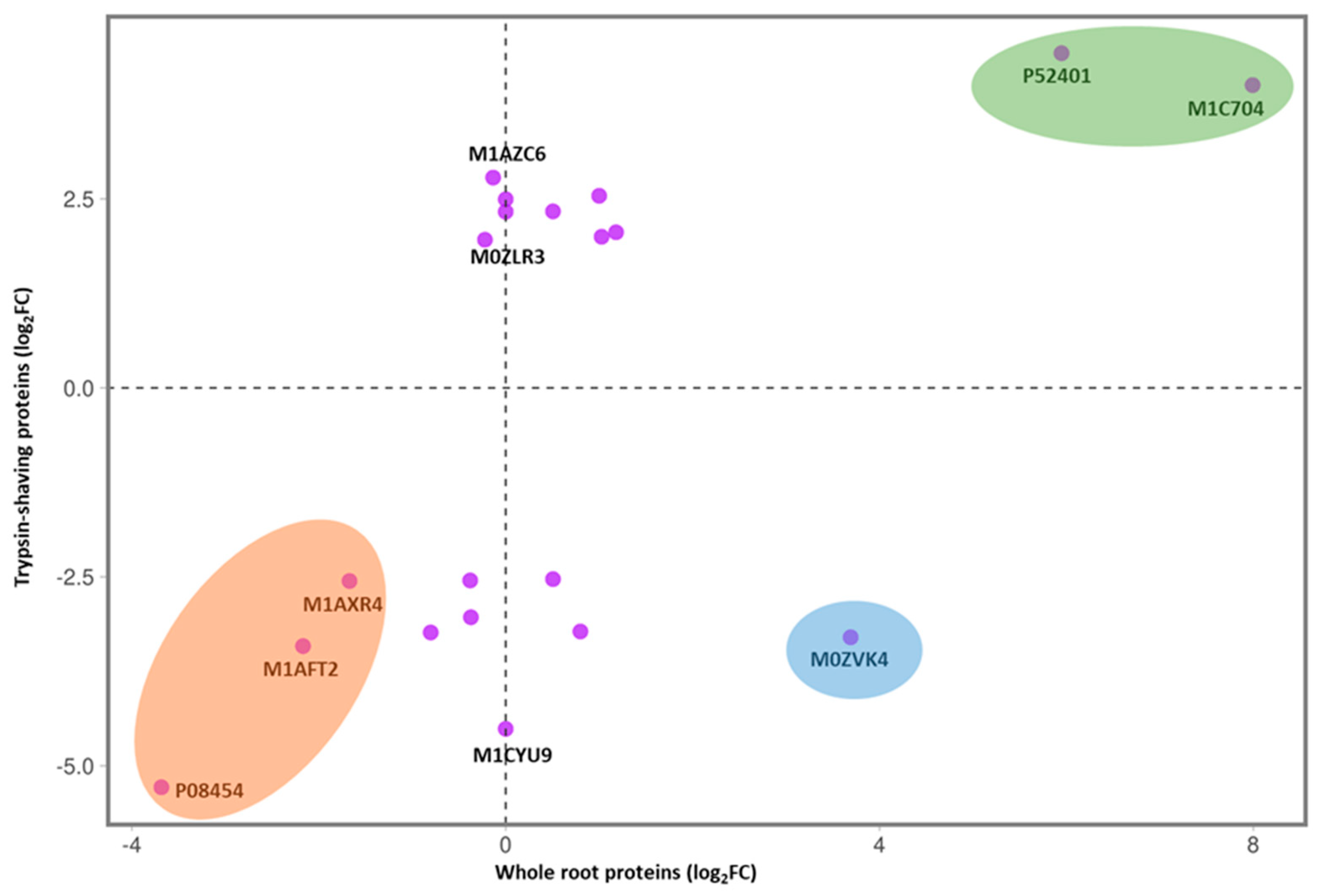
3.4. Comparison of Proteins Identified by TS with Whole-Root Proteomics and Transcriptomics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Balendres, M.A.; Tegg, R.S.; Wilson, C.R. Key events in pathogenesis of Spongospora diseases in potato: A review. Australas. Plant Pathol. 2016, 45, 229–240. [Google Scholar] [CrossRef]
- Burki, F.; Kudryavtsev, A.; Matz, M.V.; Aglyamova, G.V.; Bulman, S.; Fiers, M.; Keeling, P.J.; Pawlowski, J. Evolution of Rhizaria: New insights from phylogenomic analysis of uncultivated protists. Bmc Evol. Biol. 2010, 10, 377. [Google Scholar] [CrossRef] [PubMed]
- Falloon, R.E.; Merz, U.; Butler, R.C.; Curtin, D.; Lister, R.A.; Thomas, S.M. Root infection of potato by Spongospora subterranea: Knowledge review and evidence for decreased plant productivity. Plant Pathol. 2016, 65, 422–434. [Google Scholar] [CrossRef]
- Yu, X.; Tegg, R.S.; Eyles, A.; Wilson, A.J.; Wilson, C.R. Development and validation of a novel rapid in-vitro assay for resistance of potato cultivars to Spongospora subterranea zoospore root attachment. Plant Pathol. 2022. Submitted and in the revision. [Google Scholar] [CrossRef]
- Deacon, J. Behavioural responses of fungal zoospores. Microbiol. Sci. 1988, 5, 249–252. [Google Scholar]
- Raftoyannis, Y.; Dick, M.W. Effects of plant culture method, plant age, zoospore concentration and temperature on zoospore encystment of Phytophthora and Pythium species on plant roots. Arch. Phytopathol. Plant Prot. 2006, 39, 69–77. [Google Scholar] [CrossRef]
- Amponsah, J.; Tegg, R.S.; Thangavel, T.; Wilson, C.R. Moments of weaknesses—Exploiting vulnerabilities between germination and encystment in the Phytomyxea. Biol. Rev. 2021, 96, 1603–1615. [Google Scholar] [CrossRef]
- Keskin, B.; Fuchs, W.H. Der infektionsvorgang bei Polymyxa betae. Arch Mikrobiol 1969, 68, 218–226. [Google Scholar]
- Merz, U. Infectivity, inoculum density and germination of Spongospora subterranea resting spores: A solution-culture test system. EPPO Bull. 1989, 19, 585–592. [Google Scholar] [CrossRef]
- Merz, U. Powdery scab of potato—Occurrence, life cycle and epidemiology. Am. J. Potato Res. 2008, 85, 241–246. [Google Scholar] [CrossRef]
- Callow, J.A.; Estradagarcia, T.; Green, J.R. Recognition of non-self: The causation and avoidance of disease. Ann. Bot. 1987, 60, 3–14. [Google Scholar] [CrossRef]
- Sarkar, D.; Sun, Y.; Tayagui, A.; Adams, R.; Garrill, A.; Nock, V. Microfluidic platform to study electric field based root targeting by pathogenic zoospores. In Proceedings of the IEEE Symposium on Mass Storage Systems and Technologies, Tokyo, Japan, 9–13 January 2022; pp. 884–887. [Google Scholar]
- Estradagarcia, T.; Ray, T.C.; Green, J.R.; Callow, J.A.; Kennedy, J.F. Encystment of Pythium aphanidermatum zoospores is induced by root mucilage polysaccharides, pectin and a monoclonal antibody to a surface antigen. J. Exp. Bot. 1990, 41, 693–699. [Google Scholar] [CrossRef]
- Hinch, J.M.; Clarke, A.E. Adhesion of fungal zoospores to root surfaces is mediated by carbohydrate determinants of the root slime. Physiol. Plant Pathol. 1980, 16, 303–307. [Google Scholar]
- Longman, D.; Callow, J.A. Specific saccharide residues are involved in the recognition of plant-root surfaces by zoospores of Pythium aphanidermatum. Physiol. Mol. Plant Pathol. 1987, 30, 139–150. [Google Scholar] [CrossRef]
- Irving, H.R.; Grant, B.R. The effects of pectin and plant root surface carbohydrates on encystment and development of Phytophthora cinnamomi zoospores. J. Gen. Microbiol. 1984, 130, 1015–1018. [Google Scholar] [CrossRef]
- Grant, B.R.; Irving, H.R.; Radda, M. The effect of pectin and related-compounds on encystment and germination of Phytophthora palmivora zoospores. J. Gen. Microbiol. 1985, 131, 669–676. [Google Scholar] [CrossRef]
- Hardham, A.R.; Suzaki, E. Encystment of zoospores of the fungus, Phytophthora cinnamomi, is induced by specific lectin and monoclonal antibody binding to the cell surface. Protoplasma 1986, 133, 165–173. [Google Scholar]
- Hardham, A.R. Lectin and antibody labelling of surface components of spores of Phytophthora cinnamomi. Aust. J. Plant Pathol. 1989, 133, 165–173. [Google Scholar]
- Dill, B.C.; Fuller, M.S. Amino acid immobilization of fungal motile cells. Arch. Fur Mikrobiol. 1971, 78, 92–98. [Google Scholar] [CrossRef]
- Grant, B.R.; Griffith, J.M.; Irving, H.R. A model to explain ion-induced differentiation in zoospores of Phytophthora palmivora. Exp. Mycol. 1986, 10, 89–98. [Google Scholar] [CrossRef]
- Byrt, P.N.; Irving, H.R.; Grant, B.R. The effect of cations on zoospores of the fungus Phytophthora cinnamomi. J. Gen. Microbiol. 1982, 128, 1189–1198. [Google Scholar] [CrossRef]
- Byrt, P.N.; Irving, H.R.; Grant, B.R. The Effect of Organic-Compounds on the Encystment, Viability and Germination of Zoospores of Phytophthora-Cinnamomi. J. Gen. Microbiol. 1982, 128, 2343–2351. [Google Scholar]
- Shaw, B.; Hoch, H. Ions regulate spore attachment, germination, and fungal growth. In Biology of the Fungal Cell; Springer: Berlin/Heidelberg, Germany, 2007; pp. 219–236. [Google Scholar]
- Jones, M.J.; Epstein, L. Adhesion of Nectria haematococca macroconidia. Physiol. Mol. Plant Pathol. 1989, 35, 453–461. [Google Scholar] [CrossRef]
- Jones, M.J.; Epstein, L. Adhesion of macroconidia to the plant surface and virulence of Nectria haematococca. Appl. Environ. Microbiol. 1990, 56, 3772–3778. [Google Scholar] [CrossRef]
- Jones, S.W.; Donaldson, S.P.; Deacon, J.W. Behavior of zoospores and zoospore cysts in relation to root infection by Pythium aphanidermatum. New Phytol. 1991, 117, 289–301. [Google Scholar] [CrossRef]
- Donaldson, S.P.; Deacon, J.W. Differential encystment of zoospores of Pythium species by saccharides in relation to establishment on roots. Physiol. Mol. Plant Pathol. 1993, 42, 177–184. [Google Scholar] [CrossRef]
- Downer, A.J.; Menge, J.A.; Pond, E. Effects of cellulytic enzymes on Phytophthora cinnamomi. Phytopathology 2001, 91, 839–846. [Google Scholar] [CrossRef]
- Dunn, A.R.; Smart, C.D. Interactions of Phytophthora capsici with resistant and susceptible pepper roots and stems. Phytopathology 2015, 105, 1355–1361. [Google Scholar]
- Yu, X.; Wilson, R.; Balotf, S.; Tegg, R.S.; Eyles, A.; Wilson, C.R. Comparative proteomic analysis of potato roots from resistant and susceptible cultivars to Spongospora subterranea zoospore root attachment in vitro. Molecules 2022, 27, 6024. [Google Scholar] [CrossRef]
- Balotf, S.; Wilson, R.; Nichols, D.S.; Tegg, R.S.; Wilson, C.R. Multi-omics reveals mechanisms of resistance to potato root infection by Spongospora subterranea. Sci. Rep. 2022, 12, 10804. [Google Scholar] [CrossRef]
- Falloon, R.E.; Genet, R.A.; Wallace, A.R.; Butler, R.C. Susceptibility of potato (Solanum tuberosum) cultivars to powdery scab (caused by Spongospora subterranea f. sp subterranea), and relationships between tuber and root infection. Australas. Plant Pathol. 2003, 32, 377–385. [Google Scholar] [CrossRef]
- Sigma-Aldrich. Technical Bulletin (PNGase F P7367). Available online: https://www.sigmaaldrich.com/deepweb/assets/sigmaaldrich/product/documents/418/348/p7367bul.pdf (accessed on 1 May 2022).
- Byrt, C.S.; Cahyanegara, R.; Grof, C.P. Plant carbohydrate binding module enhances activity of hybrid microbial cellulase enzyme. Front. Plant Sci. 2012, 3, 254. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Q.; Yu, X.; Bao, C.; Wang, L.; Li, M.; Gan, J.H.; Qu, D.; Ma, J.B.; Chen, L. Identification and characterization of a novel prokaryotic peptide n-glycosidase from Elizabethkingia meningoseptica. J. Biol. Chem. 2015, 290, 7452–7462. [Google Scholar] [CrossRef]
- Vorwerk, S.; Somerville, S.; Somerville, C. The role of plant cell wall polysaccharide composition in disease resistance. Trends Plant Sci. 2004, 9, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Castilleux, R.; Plancot, B.; Ropitaux, M.; Carreras, A.; Leprince, J.; Boulogne, I.; Follet-Gueye, M.-L.; Popper, Z.A.; Driouich, A.; Vicré, M. Cell wall extensins in root–microbe interactions and root secretions. J. Exp. Bot. 2018, 69, 4235–4247. [Google Scholar] [PubMed]
- Mitchell, R.T.; Deacon, J.W. Differential (Host-specific) accumulation of zoospores of Phthium on roots of graminaceous and non-graminaceous plants. New Phytol. 1986, 102, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Gubler, F.; Hardham, A.R. Secretion of adhesive material during encystment of Phytophthora cinnamomi zoospores, characterized by immunogold labeling with monoclonal-antibodies to components of peripheral vesicles. J. Cell Sci. 1988, 90, 225–235. [Google Scholar]
- Koller, A.; Washburn, M.P.; Lange, B.M.; Andon, N.L.; Deciu, C.; Haynes, P.A.; Hays, L.; Schieltz, D.; Ulaszek, R.; Wei, J.; et al. Proteomic survey of metabolic pathways in rice. Proc. Natl. Acad. Sci. United States Am. 2002, 99, 11969–11974. [Google Scholar] [CrossRef]
- Wang, H.X.; Fan, W.J.; Li, H.; Yang, J.; Huang, J.R.; Zhang, P. Functional Characterization of Dihydroflavonol-4-Reductase in Anthocyanin Biosynthesis of Purple Sweet Potato Underlies the Direct Evidence of Anthocyanins Function against Abiotic Stresses. PLoS ONE 2013, 8, e78484. [Google Scholar] [CrossRef]
- Mahmood, Q.; Ahmad, R.; Kwak, S.-S.; Rashid, A.; Anjum, N.A. Ascorbate and glutathione: Protectors of plants in oxidative stress. In Ascorbate-Glutathione Pathway and Stress Tolerance in Plants; Springer: Berlin/Heidelberg, Germany, 2010; pp. 209–229. [Google Scholar]
- Zechmann, B.; Müller, M. Subcellular compartmentation of glutathione in dicotyledonous plants. Protoplasma 2010, 246, 15–24. [Google Scholar]
- Pei, D.; Ma, H.; Zhang, Y.; Ma, Y.; Wang, W.; Geng, H.; Wu, J.; Li, C. Virus-induced gene silencing of a putative glutathione S-transferase gene compromised Ol-1-mediated resistance against powdery mildew in tomato. Plant Mol. Biol. Report. 2011, 29, 972–978. [Google Scholar] [CrossRef]
- Zhao, J.; Buchwaldt, L.; Rimmer, S.R.; Sharpe, A.; McGregor, L.; Bekkaoui, D.; Hegedus, D. Patterns of differential gene expression in Brassica napus cultivars infected with Sclerotinia sclerotiorum. Mol. Plant Pathol. 2009, 10, 635–649. [Google Scholar] [PubMed]
- Islam, M.S.; Choudhury, M.; Majlish, A.-N.K.; Islam, T.; Ghosh, A. Comprehensive genome-wide analysis of glutathione S-transferase gene family in potato (Solanum tuberosum L.) and their expression profiling in various anatomical tissues and perturbation conditions. Gene 2018, 639, 149–162. [Google Scholar] [PubMed]
- Ithal, N.; Recknor, J.; Nettleton, D.; Maier, T.; Baum, T.J.; Mitchum, M.G. Developmental transcript profiling of cyst nematode feeding cells in soybean roots. Mol. Plant-Microbe Interact. 2007, 20, 510–525. [Google Scholar] [PubMed]
- He, Z.G.; De Buck, J. Localization of proteins in the cell wall of Mycobacterium avium subsp paratuberculosis K10 by proteomic analysis. Proteome Sci. 2010, 8, 21. [Google Scholar] [CrossRef] [PubMed]
- Zahir, M.; Fogliano, V.; Capuano, E. Effect of soybean processing on cell wall porosity and protein digestibility. Food Funct. 2020, 11, 285–296. [Google Scholar]
- Nessler, C.L.; Allen, R.D.; Galewsky, S. Identification and characterization of latex-specific proteins in opium poppy. Plant Physiol. 1985, 79, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Nessler, C.L.; Kurz, W.G.; Pelcher, L.E. Isolation and analysis of the major latex protein genes of opium poppy. Plant Mol. Biol. 1990, 15, 951–953. [Google Scholar] [PubMed]
- Suyama, T.; Yamada, K.; Mori, H.; Takeno, K.; Yamaki, S. Cloning cDNAs for genes preferentially expressed during fruit growth in cucumber. J. Am. Soc. Hortic. Sci. 1999, 124, 136–139. [Google Scholar] [CrossRef]
- Ruperti, B.; Bonghi, C.; Ziliotto, F.; Pagni, S.; Rasori, A.; Varotto, S.; Tonutti, P.; Giovannoni, J.J.; Ramina, A. Characterization of a major latex protein (MLP) gene down-regulated by ethylene during peach fruitlet abscission. Plant Sci. 2002, 163, 265–272. [Google Scholar] [CrossRef]
- Inui, H.; Sawada, M.; Goto, J.; Yamazaki, K.; Kodama, N.; Tsuruta, H.; Eun, H. A major latex-like protein is a key factor in crop contamination by persistent organic pollutants. Plant Physiol. 2013, 161, 2128–2135. [Google Scholar] [PubMed]
- Zhu, Y.; Wu, N.; Song, W.; Yin, G.; Qin, Y.; Yan, Y.; Hu, Y. Soybean (Glycine max) expansin gene superfamily origins: Segmental and tandem duplication events followed by divergent selection among subfamilies. BMC Plant Biol. 2014, 14, 93. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Li, R.; Shen, W.; Jiao, S.; Zhang, J.; Xu, W. Genome-wide evolutionary characterization and expression analyses of major latex protein (MLP) family genes in Vitis vinifera. Mol. Genet. Genom. 2018, 293, 1061–1075. [Google Scholar] [CrossRef]
- Lytle, B.L.; Song, J.; Norberto, B.; Peterson, F.C.; Johnson, K.A.; Bingman, C.A.; Phillips Jr, G.N.; Volkman, B.F. Structures of two Arabidopsis thaliana major latex proteins represent novel helix-grip folds. Proteins 2009, 76, 237. [Google Scholar]
- Dai, N.; Kandel-Kfir, M.; Petreikov, M.; Hanael, R.; Levin, I.; Ricard, B.; Rothan, C.; Schaffer, A.A.; Granot, D. The tomato hexokinase LeHXK1 cloning, mapping, expression pattern and phylogenetic relationships. Plant Sci. 2002, 163, 581–590. [Google Scholar] [CrossRef]
- Park, C.J.; Kim, K.J.; Shin, R.; Park, J.M.; Shin, Y.C.; Paek, K.H. Pathogenesis-related protein 10 isolated from hot pepper functions as a ribonuclease in an antiviral pathway. Plant J. 2004, 37, 186–198. [Google Scholar] [PubMed]
- He, S.; Yuan, G.; Bian, S.; Han, X.; Liu, K.; Cong, P.; Zhang, C. Major latex protein MdMLP423 negatively regulates defense against fungal infections in apple. Int. J. Mol. Sci. 2020, 21, 1879. [Google Scholar] [CrossRef] [PubMed]
- Sergeant, K.; Printz, B.; Guerriero, G.; Renaut, J.; Lutts, S.; Hausman, J.-F. The dynamics of the cell wall proteome of developing alfalfa stems. Biology 2019, 8, 60. [Google Scholar] [CrossRef]
- Nguema-Ona, E.; Vicré-Gibouin, M.; Gotté, M.; Plancot, B.; Lerouge, P.; Bardor, M.; Driouich, A. Cell wall O-glycoproteins and N-glycoproteins: Aspects of biosynthesis and function. Front. Plant Sci. 2014, 5, 499. [Google Scholar] [CrossRef]
- Doxey, A.C.; Yaish, M.W.; Moffatt, B.A.; Griffith, M.; McConkey, B.J. Functional divergence in the Arabidopsis β-1, 3-glucanase gene family inferred by phylogenetic reconstruction of expression states. Mol. Biol. Evol. 2007, 24, 1045–1055. [Google Scholar] [CrossRef]
- Shinshi, H.; Wenzler, H.; Neuhaus, J.-M.; Felix, G.; Hofsteenge, J.; Meins Jr, F. Evidence for N-and C-terminal processing of a plant defense-related enzyme: Primary structure of tobacco prepro-β-1, 3-glucanase. Proc. Natl. Acad. Sci. 1988, 85, 5541–5545. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Yang, Y. Disease resistance and abiotic stress tolerance in rice are inversely modulated by an abscisic acid–inducible mitogen-activated protein kinase. Plant Cell 2003, 15, 745–759. [Google Scholar] [PubMed]
- Harris, J.M. Abscisic acid: Hidden architect of root system structure. Plants 2015, 4, 548–572. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.; Wilson, R.; Eyles, A.; Balotf, S.; Tegg, R.S.; Wilson, C.R. Enzymatic Investigation of Spongospora subterranea Zoospore Attachment to Roots of Potato Cultivars Resistant or Susceptible to Powdery Scab Disease. Proteomes 2023, 11, 7. https://doi.org/10.3390/proteomes11010007
Yu X, Wilson R, Eyles A, Balotf S, Tegg RS, Wilson CR. Enzymatic Investigation of Spongospora subterranea Zoospore Attachment to Roots of Potato Cultivars Resistant or Susceptible to Powdery Scab Disease. Proteomes. 2023; 11(1):7. https://doi.org/10.3390/proteomes11010007
Chicago/Turabian StyleYu, Xian, Richard Wilson, Alieta Eyles, Sadegh Balotf, Robert Stephen Tegg, and Calum Rae Wilson. 2023. "Enzymatic Investigation of Spongospora subterranea Zoospore Attachment to Roots of Potato Cultivars Resistant or Susceptible to Powdery Scab Disease" Proteomes 11, no. 1: 7. https://doi.org/10.3390/proteomes11010007
APA StyleYu, X., Wilson, R., Eyles, A., Balotf, S., Tegg, R. S., & Wilson, C. R. (2023). Enzymatic Investigation of Spongospora subterranea Zoospore Attachment to Roots of Potato Cultivars Resistant or Susceptible to Powdery Scab Disease. Proteomes, 11(1), 7. https://doi.org/10.3390/proteomes11010007