
Potential Impacts of Climate Change on the Richness and Distribution of Endemic Anurans from the Montane Cloud Forest of Mexico

 , and
, and
Abstract
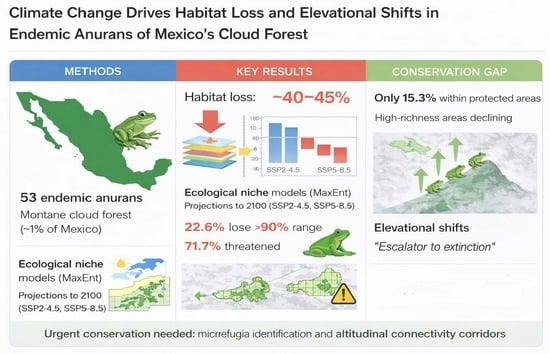
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Species Records and Occurrence Data
2.3. Environmental Data and Determination of the Accessible Area (M)
2.4. Species Distribution Modeling
2.5. Richness, Turnover, and Vulnerability
2.6. Diversity Indices and Assemblage Structure Analysis
2.7. Assessment of Species Representativeness in Protected Natural Areas
3. Results
3.1. Species and Model Performance
3.2. Changes in Assemblage Diversity Structure
3.3. Species Vulnerability and Changes in Richness
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CF | Cloud Forest |
References
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef]
- Pandit, J.; Sharma, A.K. A comprehensive review of climate change’s imprint on ecosystems. J. Water Clim. Change 2023, 14, 4273–4284. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef]
- Urban, M.C. Accelerating extinction risk from climate change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef] [PubMed]
- Duellman, W.E.; Trueb, L. Biology of Amphibians; The Johns Hopkins University Press: Baltimore, MD, USA, 1994. [Google Scholar]
- Wells, K.D. The Ecology and Behavior of Amphibians; University of Chicago Press: Chicago, IL, USA, 2007. [Google Scholar]
- Blaustein, A.R.; Walls, S.C.; Bancroft, B.A.; Lawler, J.J.; Searle, C.L.; Gervasi, S.S. Direct and indirect effects of climate change on amphibian populations. Diversity 2010, 2, 281–313. [Google Scholar] [CrossRef]
- Duarte, H.; Tejedo, M.; Katzenberger, M.; Marangoni, F.; Baldo, D.; Beltrán, J.F.; Álvarez-Villanueva, A.; Nicieza, A.G. Can amphibians take the heat? Vulnerability to climate warming in subtropical and temperate larval amphibian communities. Glob. Change Biol. 2012, 18, 412–421. [Google Scholar] [CrossRef]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef]
- Hof, C.; Araújo, M.B.; Jetz, W.; Rahbek, C. Additive threats from pathogens, climate and land-use change for global amphibian diversity. Nature 2011, 480, 516–519. [Google Scholar] [CrossRef]
- Li, Y.; Cohen, J.M.; Rohr, J.R. Review and synthesis of the effects of climate change on amphibians. Integr. Zool. 2013, 8, 145–161. [Google Scholar] [CrossRef] [PubMed]
- Blaustein, A.R.; Belden, L.K.; Olson, D.H.; Green, D.M.; Root, T.L.; Kiesecker, J.M. Amphibian breeding and climate change. Conserv. Biol. 2003, 17, 1454–1461. [Google Scholar] [CrossRef]
- Catenazzi, A. State of the world’s amphibians. Annu. Rev. Environ. Resour. 2015, 40, 91–119. [Google Scholar] [CrossRef]
- University of California. AmphibiaWeb; University of California: Berkeley, CA, USA, 2024; Available online: https://amphibiaweb.org (accessed on 5 January 2026).
- Balderas-Valdivia, C.J.; González-Hernández, A. Inventario de la herpetofauna de México 2021. Herpetol. Mex. 2021, 2, 10–71. [Google Scholar] [CrossRef]
- Ramírez-Bautista, A.; Torres-Hernández, L.A.; Cruz-Elizalde, R.; Berriozabal-Islas, C.; Hernández-Salinas, U.; Wilson, L.D.; Leyte-Manrique, A.; Morales-Martínez, D.M. An updated list of the Mexican herpetofauna: With a summary of historical and contemporary studies. ZooKeys 2023, 1166, 287–306. [Google Scholar] [CrossRef]
- Abarca-Alvarado, J.G. Endangered amphibians: Threats and effective conservation strategies. Biocenosis 2021, 32, 133–145. [Google Scholar] [CrossRef]
- Pottier, P.; Kearney, M.R.; Wu, N.C.; Gunderson, A.R.; Rej, J.E.; Rivera-Villanueva, A.N.; Pollo, P.; Burke, S.; Drobniak, S.M.; Nakagawa, S. Vulnerability of amphibians to global warming. Nature 2025, 639, 954–961. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Raven, P.H. Vertebrates on the brink as indicators of biological annihilation and the sixth mass extinction. Proc. Natl. Acad. Sci. USA 2020, 117, 13596–13602. [Google Scholar] [CrossRef] [PubMed]
- Wake, D.B.; Vredenburg, V.T. Are we in the midst of the sixth mass extinction? A view from the world of amphibians. Proc. Natl. Acad. Sci. USA 2008, 105, 11466–11473. [Google Scholar] [CrossRef]
- Barnosky, A.D.; Matzke, N.; Tomiya, S.; Wogan, G.O.U.; Swartz, B.; Quental, T.B.; Marshall, C.; McGuire, J.L.; Lindsey, E.L.; Maguire, K.C.; et al. Approaching a state shift in Earth’s biosphere. Nature 2012, 486, 52–58. [Google Scholar] [CrossRef]
- Araújo, M.B.; Thuiller, W.; Pearson, R.G. Climate warming and the decline of amphibians and reptiles in Europe. J. Biogeogr. 2006, 33, 1712–1728. [Google Scholar] [CrossRef]
- García, A.; Ortega-Huerta, M.A.; Martínez-Meyer, E. Potential distributional changes and conservation priorities of endemic amphibians in western Mexico as a result of climate change. Environ. Conserv. 2013, 47, 1–12. [Google Scholar] [CrossRef]
- Ochoa-Ochoa, L.; Rodríguez, P.; Mora, F.; Flores-Villela, O.; Whittaker, R. Climate change and amphibian diversity patterns in Mexico. Biol. Conserv. 2012, 150, 94–102. [Google Scholar] [CrossRef]
- Estrada-Porrúa, F.; Zavala-Hidalgo, J.; Martínez-Arroyo, A.; Raga, G.; Gay-García, C. Estado y Perspectivas del Cambio Climático en México: Un Punto de Partida; Programa de Investigación en Cambio Climático, Universidad Nacional Autónoma de México: Mexico City, Mexico, 2023; Available online: https://cambioclimatico.unam.mx/estado-y-perspectivas-del-cambio-climatico-en-mexico/ (accessed on 19 February 2026).
- Toledo-Aceves, T.; Meave, J.A.; González-Espinosa, M.; Ramírez-Marcial, N. Tropical montane cloud forests: Current threats and opportunities for their conservation and sustainable management in Mexico. J. Environ. Manag. 2011, 92, 974–981. [Google Scholar] [CrossRef] [PubMed]
- Ponce-Reyes, R.; Reynoso-Rosales, V.H.; Watson, J.E.M.; VanDerWal, J.; Fuller, R.A.; Pressey, R.L.; Possingham, H.P. Vulnerability of cloud forest reserves in Mexico to climate change. Nat. Clim. Change 2012, 2, 448–452. [Google Scholar] [CrossRef]
- Gual-Díaz, M.; Rendón-Correa, A. (Eds.) Bosques Mesófilos de Montaña de México: Diversidad, Ecología y Manejo; CONABIO: Mexico City, Mexico, 2014.
- Flores-Villela, O.; Gerez, P. Biodiversidad y Conservación en México: Vertebrados, Vegetación y Uso del Suelo; UNAM-CONABIO: Mexico City, Mexico, 1994. [Google Scholar]
- Almazán-Núñez, R.C.; Alvarez-Alvarez, E.A.; Ruiz-Gutiérrez, F.; Toribio-Jiménez, S. Biological survey of a cloud forest in southwestern Mexico: Plants, amphibians, reptiles, birds, and mammals. Biota Neotrop. 2018, 18, e20170444. [Google Scholar] [CrossRef]
- Cruz-Elizalde, R.; Ochoa-Ochoa, L.M.; Flores-Villela, O.A.; Velasco, J.A. Taxonomic distinctiveness and phylogenetic variability of amphibians and reptiles in the cloud forest of Mexico. Community Ecol. 2022, 23, 87–102. [Google Scholar] [CrossRef]
- NOM-059-SEMARNAT-2010; Norma Oficial Mexicana, Protección Ambiental—Especies Nativas de México de Flora y Fauna Silvestres—Categorías de Riesgo y Especificaciones para su Inclusión, Exclusión o Cambio—Lista de Especies en Riesgo. Diario Oficial de la Federación; SEMARNAT: Mexico City, Mexico, 2010.
- IUCN. The IUCN Red List of Threatened Species; Version 2022-2; IUCN: Gland, Switzerland, 2022; Available online: https://www.iucnredlist.org (accessed on 3 January 2026).
- Cruz-Elizalde, R.; Ochoa-Ochoa, L.M.; Flores-Villela, O.A. La importancia de los endemismos de vertebrados en el bosque mesófilo de montaña en México. Cuad. Biodivers. 2022, 63, 30–39. [Google Scholar] [CrossRef]
- Chen, I.C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Lawler, J.J.; Shafer, S.L.; Bancroft, B.A.; Blaustein, A.R. Projected climate impacts for the amphibians of the Western Hemisphere. Conserv. Biol. 2010, 24, 38–47. [Google Scholar] [CrossRef]
- Foden, W.B.; Butchart, S.H.M.; Stuart, S.N.; Vié, J.C.; Akçakaya, H.R.; Angulo, A.; DeVantier, L.M.; Gutsche, A.; Turak, E.; Cao, L.; et al. Identifying the world’s most climate change vulnerable species: A systematic trait-based assessment of all birds, amphibians and corals. PLoS ONE 2013, 8, e65427. [Google Scholar] [CrossRef]
- Ballesteros-Barrera, C.; Tapia-Pérez, O.; Zárate-Hernández, R.; Leyte-Manrique, A.; Martínez-Bernal, A.; Vargas-Miranda, B.; Sigala-Rodríguez, J.J. The potential effect of climate change on the distribution of endemic anurans from Mexico’s tropical dry forest. Diversity 2022, 14, 650. [Google Scholar] [CrossRef]
- Rzedowski, J. Vegetación de México; Limusa: Mexico City, Mexico, 1978. [Google Scholar]
- Challenger, A. Utilización y Conservación de los Ecosistemas Terrestres de México: Pasado, Presente y Futuro; CONABIO-UNAM-Agrupación Sierra Madre: Mexico City, Mexico, 1998. [Google Scholar]
- González-Espinosa, M.; Meave, J.A.; Lorea-Hernández, F.G.; Ibarra-Manríquez, G.; Newton, A.C. The Red List of Mexican Cloud Forest Trees; Fauna & Flora International: Cambridge, UK, 2011. [Google Scholar]
- Montiel Canales, G.; Goyenechea Mayer-Goyenechea, I. Amphibian areas of endemism: A conservation priority in the threatened Mexican cloud forest. Vertebr. Zool. 2022, 72, 235–244. [Google Scholar] [CrossRef]
- Brown, J.L. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2016. [Google Scholar]
- Saha, A.; Rahman, S.; Alam, S. Modeling current and future potential distributions of desert locust Schistocerca gregaria under climate change scenarios using MaxEnt. J. Asia-Pac. Biodivers. 2021, 14, 399–409. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Vasconcelos, T.S.; Rodríguez, M.Á.; Hawkins, B.A. Species distribution modelling as a macroecological tool: A case study using New World amphibians. Ecography 2012, 35, 539–548. [Google Scholar] [CrossRef]
- Barve, N.; Barve, V.; Jiménez-Valverde, A.; Lira-Noriega, A.; Maher, S.P.; Peterson, A.T.; Soberón, J.; Villalobos, F. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Model. 2011, 222, 1810–1819. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological Niches and Geographic Distributions; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: https://www.R-project.org/ (accessed on 7 November 2025).
- Shcheglovitova, M.; Anderson, R.P. Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecol. Model. 2013, 269, 9–17. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Rzedowski, J. Vegetación Potencial. IV.8.2. In Atlas Nacional de México; Vol. II, Escala 1:4,000,000; Instituto de Geografía, UNAM: Mexico City, Mexico, 1990; Available online: http://www.conabio.gob.mx/informacion/gis/?vns=gis_root/usv/otras/vpr4mgw (accessed on 1 January 2026).
- Hijmans, R.J. Terra: Spatial Data Analysis, R Package Version 1.7-29; R Foundation for Statistical Computing: Vienna, Austria, 2023. Available online: https://CRAN.R-project.org/package=terra (accessed on 1 January 2026).
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- CONANP (Comisión Nacional de Áreas Naturales Protegidas). Áreas Naturales Protegidas Federales de México, Septiembre 2024 [Shapefile]; CONANP: Mexico City, Mexico, 2024. Available online: http://geoportal.conabio.gob.mx/metadatos/doc/html/anpsp2024gw.html (accessed on 1 January 2026).
- Pebesma, E. Simple features for R: Standardized support for spatial vector data. R J. 2018, 10, 439–446. [Google Scholar] [CrossRef]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Ochoa-Ochoa, L.; Flores-Villela, O. Áreas de Diversidad y Endemismo de la Herpetofauna Mexicana; UNAM-CONABIO: Mexico City, Mexico, 2006. [Google Scholar]
- Luedtke, J.A.; Chanson, J.; Neam, K.; Hobin, L.; Maciel, A.O.; Catenazzi, A.; Borzée, A.; Hamidy, A.; Aowphol, A.; Auth, D.; et al. Ongoing declines for the world’s amphibians in the face of emerging threats. Nature 2023, 622, 308–314. [Google Scholar] [CrossRef]
- Frías-Álvarez, P.; Zúñiga-Vega, J.J.; Flores-Villela, O. A general assessment of the conservation status and decline of amphibians in Mexico. Biol. Conserv. 2010, 19, 3699–3742. [Google Scholar] [CrossRef]
- Flores-Villela, O.; Canseco-Márquez, L. Nuevas especies y cambios taxonómicos para la herpetofauna de México. Acta Zool. Mex. (n.s.) 2004, 20, 115–144. [Google Scholar] [CrossRef]
- Jiménez-García, D.; Peterson, A.T. Climate change impact on the distribution of amphibians in Mexico. Rev. Mex. Biodivers. 2019, 90, e902841. [Google Scholar] [CrossRef]
- Yang, W.; Ma, K.; Kreft, H. Geographical sampling bias in a large distributional database and its effects on species richness–environment models. J. Biogeogr. 2013, 40, 1415–1426. [Google Scholar] [CrossRef]
- Quintero-Vallejo, D.E.; Ochoa-Ochoa, L.M. Priorización y distribución de los anfibios en las áreas naturales protegidas de México. Rev. Mex. Biodivers. 2022, 93, e933939. [Google Scholar] [CrossRef]
- Parra-Olea, G.; Flores-Villela, O.; Mendoza-Almeralla, C. Biodiversidad de anfibios en México. Rev. Mex. Biodivers. 2014, 85, S460–S466. [Google Scholar] [CrossRef]
- Rovito, S.M.; Parra-Olea, G.; Recuero, E.; Wake, D.B. Diversification and biogeographical history of Neotropical plethodontid salamanders. Zool. J. Linn. Soc. 2015, 175, 167–188. [Google Scholar] [CrossRef]
- Ochoa-Ochoa, L.M.; Campbell, J.A.; Flores-Villela, O.A. Patterns of richness and endemism of the Mexican herpetofauna, a matter of spatial scale? Biol. J. Linn. Soc. 2014, 111, 305–316. [Google Scholar] [CrossRef]
- Hijmans, R.; Graham, C. The ability of climate envelope models to predict the effect of climate change on species distributions. Glob. Change Biol. 2006, 12, 2272–2281. [Google Scholar] [CrossRef]
- Bolom-Huet, R.; Caloca-Peña, L.; Falconi-Briones, F.A.; Sunny, A. Climate-driven erosion of richness and evolutionary diversity and conservation shortfall in a salamander hotspot. Biodivers. Conserv. 2026, 35, 1–29. [Google Scholar] [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef]
- Ramírez-Barahona, S.; Cuervo-Robayo, Á.P.; Feeley, K.J.; Ortiz-Rodríguez, A.E.; Vásquez-Aguilar, A.A.; Ornelas, J.F.; Rodríguez-Correa, H. Upslope plant species shifts in Mesoamerican cloud forests driven by climate and land use change. Science 2025, 387, 1058–1063. [Google Scholar] [CrossRef] [PubMed]
- Olden, J.D.; Poff, N.L.; Douglas, M.R.; Douglas, M.E.; Fausch, K.D. Ecological and evolutionary consequences of biotic homogenization. Trends Ecol. Evol. 2004, 19, 18–24. [Google Scholar] [CrossRef]
- Clavel, J.; Julliard, R.; Devictor, V. Worldwide decline of specialist species: Toward a global functional homogenization? Front. Ecol. Environ. 2011, 9, 222–228. [Google Scholar] [CrossRef]
- Colwell, R.K.; Brehm, G.; Cardelús, C.L.; Gilman, A.C.; Longino, J.T. Global warming, elevational range shifts, and lowland biotic attrition in the wet tropics. Science 2002, 322, 258–261. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; Ferreira de Siqueira, M.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef] [PubMed]
- Menéndez-Guerrero, P.A.; Green, D.M.; Davies, T.J. Climate change and the future of Neotropical anurans biodiversity. Ecography 2020, 43, 222–235. [Google Scholar] [CrossRef]
- Guo, F.; Lenoir, J.; Bonebrake, T.C. Land-use change interacts with climate to determine elevational species redistribution. Nat. Commun. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- Smith, M.A.; Green, D.M. Dispersal and the metapopulation paradigm in amphibian ecology and conservation: Are all amphibian populations metapopulations? Ecography 2005, 28, 110–128. [Google Scholar] [CrossRef]
- Díaz-García, J.M.; Oropeza-Sánchez, M.T.; Aguilar-López, J.L. Servicios ecosistémicos de los anfibios en México: Un análisis de diversidad, distribución y conservación. Etnobiología 2019, 17, 49–60. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Bioclimatic Variables |
|---|---|
| Bio 1 | Annual mean temperature (°C) |
| Bio 2 | Mean diurnal range (mean of monthly (max temp–min temp)) (°C) |
| Bio 3 | Isothermality ((BIO2/BIO7) × 100) (°C) |
| Bio 4 | Temperature seasonality (standard deviation × 100) (°C) |
| Bio 5 | Maximum temperature of warmest month (°C) |
| Bio 6 | Minimum temperature of coldest month (°C) |
| Bio 7 | Temperature Annual Range (BIO5-BIO6) |
| Bio 8 | Mean temperature of wettest quarter (°C) |
| Bio 9 | Mean temperature of driest quarter (°C) |
| Bio 10 | Mean temperature of warmest quarter (°C) |
| Bio 11 | Mean temperature of coldest quarter (°C) |
| Bio 12 | Annual precipitation (mm) |
| Bio 13 | Precipitation of wettest month (mm) |
| Bio 14 | Precipitation of driest month (mm) |
| Bio 15 | Precipitation seasonality (standard deviation × 100) |
| Bio 16 | Precipitation of wettest quarter (mm) |
| Bio 17 | Precipitation of driest quarter (mm) |
| Bio 18 | Precipitation of warmest quarter (mm) |
| Bio 19 | Precipitation of coldest quarter (mm) |
| Index | Present | SSP2-4.5 | SSP5-8.5 | ΔSSP2-4.5 | ΔSSP5-8.5 |
|---|---|---|---|---|---|
| Alpha diversity | |||||
| Species richness (S) | 53 | 52 | 50 | −1 | −3 |
| Shannon (H′) | 3.1049 | 2.8827 | 2.8300 | −0.2222 | −0.2749 |
| Simpson (1 − D) | 0.9133 | 0.8845 | 0.8816 | −0.0288 | −0.0317 |
| Pielou’s evenness (J) | 0.7820 | 0.7296 | 0.7234 | −0.0524 | −0.0586 |
| Dominance (max pi) | 0.2379 | 0.2964 | 0.2990 | +0.0585 | +0.0611 |
| Habitat stability | |||||
| Mean stability index | 1.000 | 0.597 | 0.545 | −0.403 | −0.455 |
| Beta diversity (Baselga decomposition) | |||||
| βSOR (Sørensen dissimilarity) | — | 0.0095 | 0.0291 | — | — |
| βSIM (turnover component) | — | 0.0000 | 0.0000 | — | — |
| βNES (nestedness component) | — | 0.0095 | 0.0291 | — | — |
| Rarity-weighted vulnerability | |||||
| Rarity-weighted vulnerability index | — | 0.8186 | 0.8451 | — | — |
| Protected Natural Area | State(s) | Current Richness | SSP2-4.5 | SSP5-8.5 | ΔSSP2-4.5 | ΔSSP5-8.5 |
|---|---|---|---|---|---|---|
| Richness loss expected | ||||||
| Río Necaxa | Puebla | 6.09 | 5.35 | 4.72 | −0.74 | −1.37 |
| La Sepultura | Chiapas | 4.17 | 3.59 | 3.05 | −0.58 | −1.12 |
| La Concordia | Chiapas | 3.66 | 2.76 | 2.74 | −0.90 | −0.92 |
| Los Tuxtlas | Veracruz | 3.62 | 2.79 | 2.44 | −0.83 | −1.18 |
| Richness gain expected | ||||||
| Sierra de Manantlán | Jalisco Colima | 2.76 | — | 3.00 | — | +0.24 |
| Bosencheve | México Michoacán | 2.58 | 3.00 | 3.00 | +0.42 | +0.42 |
| Valle de Bravo | México | — | — | — | +0.76 | +0.82 |
| Mariposa Monarca | México Michoacán | 2.03 | 2.66 | 2.75 | +0.63 | +0.72 |
| Complete loss of climatic suitability | ||||||
| Montes Azules | Chiapas | — | 0 | 0 | Complete loss | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Ballesteros-Barrera, C.; Tapia-Pérez, O.; Leyte-Manrique, A.; Martínez-Bernal, A.; Zárate-Hernández, R.; Vargas-Miranda, B.; Martínez-Coronel, M.; Ortiz-Burgos, S. Potential Impacts of Climate Change on the Richness and Distribution of Endemic Anurans from the Montane Cloud Forest of Mexico. Climate 2026, 14, 114. https://doi.org/10.3390/cli14060114
Ballesteros-Barrera C, Tapia-Pérez O, Leyte-Manrique A, Martínez-Bernal A, Zárate-Hernández R, Vargas-Miranda B, Martínez-Coronel M, Ortiz-Burgos S. Potential Impacts of Climate Change on the Richness and Distribution of Endemic Anurans from the Montane Cloud Forest of Mexico. Climate. 2026; 14(6):114. https://doi.org/10.3390/cli14060114
Chicago/Turabian StyleBallesteros-Barrera, Claudia, Oscar Tapia-Pérez, Adrián Leyte-Manrique, Angélica Martínez-Bernal, Rocío Zárate-Hernández, Bárbara Vargas-Miranda, Matías Martínez-Coronel, and Selene Ortiz-Burgos. 2026. "Potential Impacts of Climate Change on the Richness and Distribution of Endemic Anurans from the Montane Cloud Forest of Mexico" Climate 14, no. 6: 114. https://doi.org/10.3390/cli14060114
APA StyleBallesteros-Barrera, C., Tapia-Pérez, O., Leyte-Manrique, A., Martínez-Bernal, A., Zárate-Hernández, R., Vargas-Miranda, B., Martínez-Coronel, M., & Ortiz-Burgos, S. (2026). Potential Impacts of Climate Change on the Richness and Distribution of Endemic Anurans from the Montane Cloud Forest of Mexico. Climate, 14(6), 114. https://doi.org/10.3390/cli14060114