Allelochemical Activity of Eugenol-Derived Coumarins on Lactuca sativa L.

 , ,
, ,  ,
,
Abstract

1. Introduction
2. Results
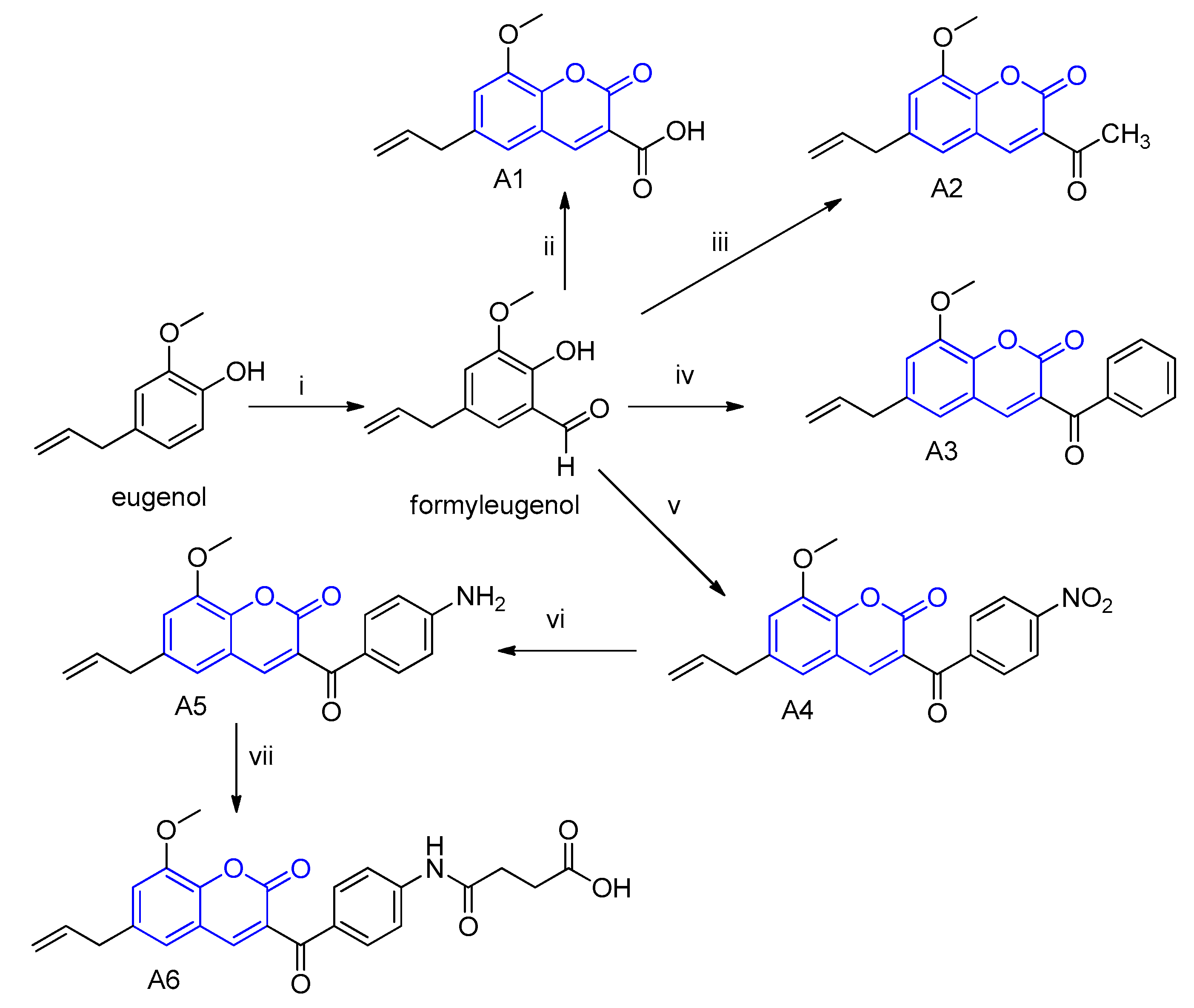
2.1. Synthesis and Characterization of the Eugenol-Derived Coumarins
2.2. Phytotoxicity Assay
2.3. Cytogenotoxicity Assay
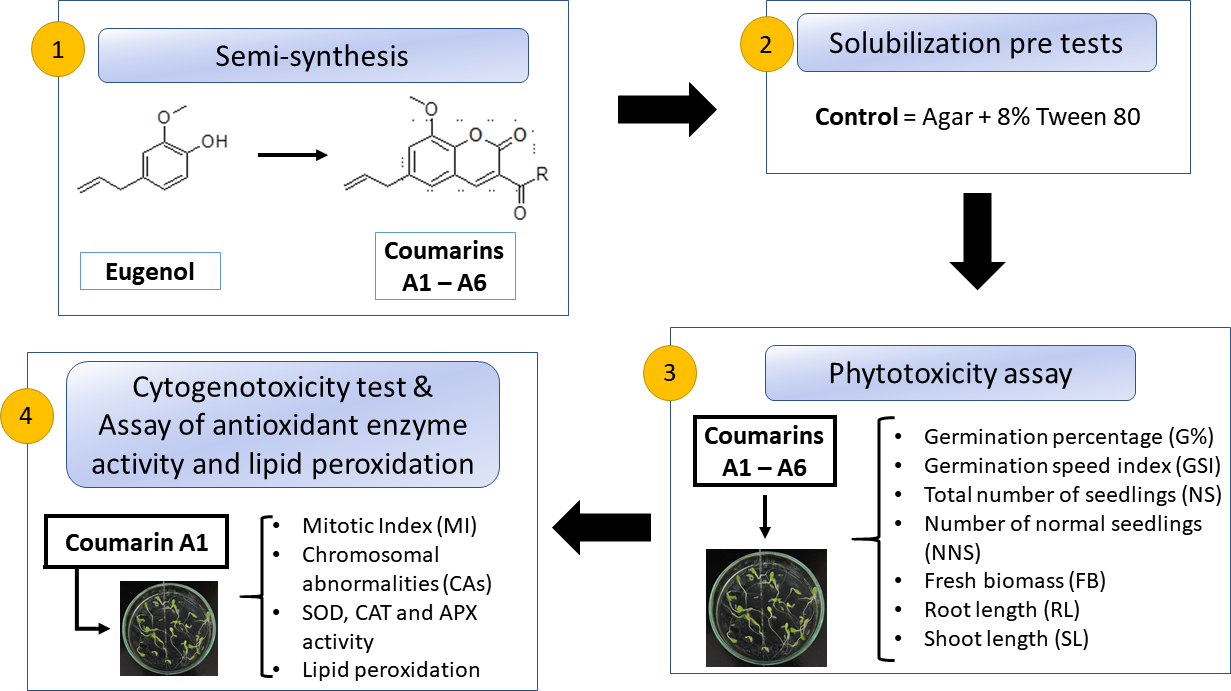
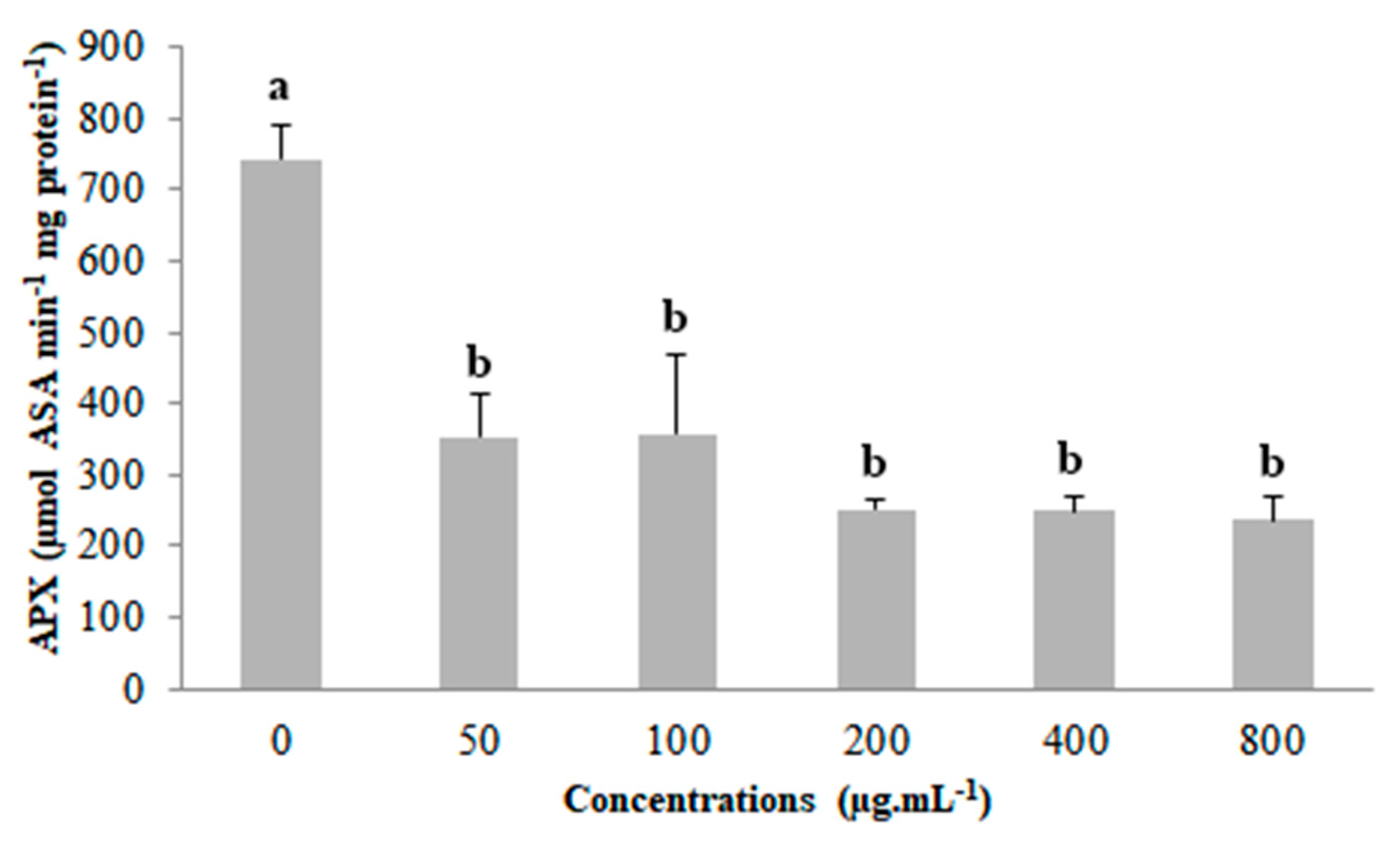
2.4. Assay of Antioxidant Enzyme Activity and Lipid Peroxidation
3. Discussion
4. Materials and Methods
4.1. Synthesis and Characterization of the Eugenol-Derived Coumarins A1–A6
4.2. Phytotoxicity Assay
4.3. Cytogenotoxicity Test
4.4. Assay of Antioxidant Enzyme Activity and Lipid Peroxidation
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Anese, S.; Jatobá, L.J.; Grisi, P.U.; Gualtieri, S.C.J.; Santos, M.F.C.; Berlinck, R.G.S. Bioherbicidal activity of drimane sesquiterpenes from Drimys brasiliensis Miers roots. Ind. Crops Prod. 2015, 74, 28–35. [Google Scholar] [CrossRef]
- Cordeau, S.; Triolet, M.; Wayman, S.; Steinberg, C.; Guillemin, J.P. Bioherbicides: Dead in the water? A review of the existing products for integrated weed management. Crop Prot. 2016, 87, 44–49. [Google Scholar] [CrossRef]
- Pes, M.P.; Mazutti, M.A.; Almeida, T.C.; Curioletti, L.E.; Melo, A.A.; Guedes, J.V.C.; Kuhn, R.C. Bioherbicide based on Diaporthe sp. secondary metabolites in the control of three tough weeds. Afr. J. Agric. Res. 2016, 11, 4242–4249. [Google Scholar] [CrossRef]
- Kuster, R.M.; Rocha, L.M. Cumarinas, cromonas e xantonas. In Farmacognosia: Da planta ao medicamento, 5th ed.; Simões, C.O.S., Schenkel, E.P., Gosmann, G., Mello, J.C.P., Mentz, L.A., Petrovick, P.R., Eds.; UFSC: Florianópolis, Brazil, 2007; pp. 537–556. [Google Scholar]
- Razavi, S.M. Plant coumarins as allelopathic agents. Int. J. Biol. Chem. 2011, 5, 86–90. [Google Scholar] [CrossRef]
- Wu, C.X.; Zhao, G.Q.; Liu, D.L.; Liu, S.J.; Gun, X.X.; Tang, Q. Discovery and weed inhibition effects of coumarin as the predominant allelochemical of yellow sweetclover (Melilotus officinalis). Int. J. Agric. Biol. 2016, 18, 168–175. [Google Scholar] [CrossRef]
- Dias, M.P.; Nozari, R.M.; Santarém, E.R. Herbicidal activity of natural compounds from Baccharis spp. on the germination and seedlings growth of Lactuca sativa and Bidens pilosa. Allelopath. J. 2017, 42, 21–35. [Google Scholar] [CrossRef]
- Araniti, F.; Mancuso, R.; Lupini, A.; Giofrè, S.; Sunseri, F.; Gabriele, B.; Abenavoli, M. Phytotoxic Potential and Biological Activity of Three Synthetic Coumarin Derivatives as New Natural-Like Herbicides. Molecules 2015, 20, 17883–17902. [Google Scholar] [CrossRef] [PubMed]
- Klenkar, J.; Molnar, M. Natural and synthetic coumarins as potential anticancer agents. J. Chem. Pharm. Res. 2015, 7, 1223–1238. [Google Scholar]
- Neelgundmath, M.; Dinesh, K.R.; Mohan, C.D.; Li, F.; Dai, X.; Siveen, K.S.; Paricharak, S.; Mason, D.J.; Fuchs, J.E.; Sethi, G.; et al. Novel synthetic coumarins that targets NF-κB in Hepatocellular carcinoma. Bioorg. Med. Chem. Lett. 2015, 25, 893–897. [Google Scholar] [CrossRef]
- Farley, C.M.; Dibwe, D.F.; Ueda, J.Y.; Hall, E.A.; Awale, S.; Magolan, J. Evaluation of synthetic coumarins for antiausterity cytotoxicity against pancreatic cancers. Bioorg. Med. Chem. Lett. 2016, 26, 1471–1474. [Google Scholar] [CrossRef]
- Ueda, C.; Tse, D.C. Stability of Catechol Modified Carbon Electrodes for Electrocatalysis of Dihudronicotinamide Adenine Dinucleotide and Ascorbic Acid. Anal. Chem. 1982, 54, 850–856. [Google Scholar] [CrossRef]
- Ciszewski, A.; Milczarek, G. Preparation and General Properties of Chemically Modified Electrodes Based on Electrosynthesized Thin Polymeric Films Derived from Eugenol. Electroanalysis 2001, 13, 860–867. [Google Scholar] [CrossRef]
- Kishore, D.; Kannan, S. Double bond migration of eugenol to isoeugenol over as-synthesized hydrotalcites and their modified forms. Appl. Catal. A Gen. 2004, 270, 227–235. [Google Scholar] [CrossRef]
- Gusman, G.S.; Bittencourt, A.H.C.; Vestena, S. Allelopathy of Baccharis dracunculifolia DC. on the germination and development of cultivated species. Acta Sci. Biol. Sci. 2008, 30, 119–125. [Google Scholar]
- Lupini, A.; Sorgona, A.; Miller, A.J.; Abenavoli, M.R. Short-term effects of coumarin along the maize primary root axis. Plant Signal. Behav. 2010, 5, 1395–1400. [Google Scholar] [CrossRef] [PubMed]
- Lupini, A.; Araniti, F.; Sunseri, F.; Abenavoli, M.R. Coumarin interacts with auxin polar transport to modify root system architecture in Arabidopsis thaliana. Plant Growth Regul. 2014, 74, 23–31. [Google Scholar] [CrossRef]
- Leme, D.M.; Marin-Morales, M.A. Allium cepa test in environmental monitoring: A review on its application. Mutat. Res. Mutat. Res. 2009, 682, 71–81. [Google Scholar] [CrossRef]
- Pesnya, D.S.; Romanovsky, A.V.; Serov, D.A.; Poddubnaya, N.Y. Genotoxic effects of Heracleum sosnowskyi in the Allium cepa test. Caryologia 2017, 70, 55–61. [Google Scholar] [CrossRef]
- Iganci, J.R.V.; Bobrowski, V.L.; Heiden, G.; Stein, V.C.; Rocha, B.H.G. Effect of aquous extracts of the “boldo” species on the germination and mitotic index of Allium cepa L. Arq. Inst. Biol. (Sao Paulo) 2006, 73, 79–82. [Google Scholar]
- Araniti, F.; Scognamiglio, M.; Chambery, A.; Russo, R.; Esposito, A.; D’Abrosca, B.; Fiorentino, A.; Lupini, A.; Sunseri, F.; Abenavoli, M.R. Highlighting the effects of coumarin on adult plants of Arabidopsis thaliana (L.) Heynh. by an integrated -omic approach. J. Plant Physiol. 2017, 213, 30–41. [Google Scholar] [CrossRef]
- Simões, M.S.; Madail, R.H.; Barbosa, S.; Nogueira, M.D.L. Bioassay standardization for the detection of allelopathic compounds and environmental toxicants using lettuce. Biotemas 2013, 26, 29–36. [Google Scholar] [CrossRef]
- Moraes, R.M.; Santos Filho, P.R.; Carvalho, M.; Nogueira, M.D.L.; Barbosa, S. Effects of copper on physiological and cytological aspects in Lactuca sativa L. Rev. Bras. Biociências 2015, 13, 115–121. [Google Scholar]
- La Pietra, V.; Marinelli, L.; Cosconati, S.; Di Leva, F.S.; Nuti, E.; Santamaria, S.; Pugliesi, I.; Morelli, M.; Casalini, F.; Rossello, A.; et al. Identification of novel molecular scaffolds for the design of MMP-13 inhibitors: A first round of lead optimization. Eur. J. Med. Chem. 2012, 47, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Brancaglion, G.A.; Toyota, A.E.; Cardoso Machado, J.V.; Fernandes Júnior, A.Á.; Silveira, A.T.; Vilas Boas, D.F.; dos Santos, E.G.; Caldas, I.S.; Carvalho, D.T. In vitro and in vivo trypanocidal activities of 8-methoxy-3-(4-nitrobenzoyl)-6-propyl-2H-cromen-2-one, a new synthetic coumarin of low cytotoxicity against mammalian cells. Chem. Biol. Drug Des. 2018, 92, 1888–1898. [Google Scholar] [CrossRef] [PubMed]
- Suksungworn, R.; Srisombat, N.; Bapia, S.; Soun-Udom, M.; Sanevas, N.; Wongkantrakorn, N.; Kermanee, P.; Vajrodaya, S.; Duangsrisai, S. Coumarins from Haldina cordifolia lead to programmed cell death in giant mimosa: Potential bio-herbicides. Pakistan J. Bot. 2017, 49, 1173–1183. [Google Scholar]
- Saleh, A.M.; Madany, M.M.Y.; González, L. The Effect of Coumarin Application on Early Growth and Some Physiological Parameters in Faba Bean (Vicia faba L.). J. Plant Growth Regul. 2015, 34, 233–241. [Google Scholar] [CrossRef]
- Samajdar, S.; Mukherjee, S.; Das, P.P. Seed Germination Inhibitors: Molecular and Phytochemical Aspects. Int. J. Appl. Pharm. Sci. Res. 2018, 3, 12–23. [Google Scholar] [CrossRef]
- Haig, T.J.; Haig, T.J.; Seal, A.N.; Pratley, J.E.; An, M.; Wu, H. Lavender as a source of novel plant compounds for the development of a natural herbicide. J. Chem. Ecol. 2009, 35, 1129–1136. [Google Scholar] [CrossRef]
- Abenavoli, M.R.; Sorgonà, A.; Albano, S.; Cacco, G. Coumarin differentially affects the morphology of different root types of maize seedlings. J. Chem. Ecol. 2004, 30, 1871–1883. [Google Scholar] [CrossRef]
- Niro, E.; Marzaioli, R.; Crescenzo, S. De; D’Abrosca, B.; Castaldi, S.; Esposito, A.; Fiorentino, A.; Rutigliano, F.A. Effects of the allelochemical coumarin on plants and soil microbial community. Soil Biol. Biochem. 2016, 95, 30–39. [Google Scholar] [CrossRef]
- Schreiner, O.; Reed, H.S. The Toxic Action of Certain Organic Plant Constituents. Bot. Gaz. 1908, 45, 73–102. [Google Scholar] [CrossRef]
- de Andrade, H.M.; Bittencourt, A.H.C.; Vestena, S. Allelopathic potential of Cyperus rotundus L. upon cultivated species. Cienc. Agrotecnol. 2009, 33, 1984–1990. [Google Scholar] [CrossRef]
- Yan, Z.; Wang, D.; Cui, H.; Zhang, D.; Sun, Y.; Jin, H.; Li, X.; Yang, X.; Guo, H.; He, X.; et al. Phytotoxicity mechanisms of two coumarin allelochemicals from Stellera chamaejasme in lettuce seedlings. Acta Physiol. Plant. 2016, 38. [Google Scholar] [CrossRef]
- Yuksel, B.; Aksoy, O. Cytological effects of coumarin on the mitosis of Lens culinaris Medik. Fresenius Environ. Bull. 2017, 26, 6400–6407. [Google Scholar]
- Thakur, A.; Singla, R.; Jaitak, V. Coumarins as anticancer agents: A review on synthetic strategies, mechanism of action and SAR studies. Eur. J. Med. Chem. 2015, 101, 476–495. [Google Scholar] [CrossRef]
- dos Santos, S.C.; de Oliveira, U.A.; de Trindade, L.O.R.; Assis, M.D.O.; Campos, J.M.S.; Salgado, E.G.; Barbosa, S. Genotypes selection for plant bioassays using Lactuca sativa L. and Allium cepa L. Pakistan J. Bot. 2017, 49, 2201–2212. [Google Scholar]
- Inzé, D.; De Veylder, L. Cell division. In Biochemistry and Molecular Biology of Plants, 2nd ed.; Buchanan, B.B., Gruissem, W., Jones, R.L., Eds.; Wiley-Blackwell: West Sussex, UK, 2015; pp. 476–506. [Google Scholar]
- Graña, E.; Costas-Gil, A.; Longueira, S.; Celeiro, M.; Teijeira, M.; Reigosa, M.J.; Sánchez-Moreiras, A.M. Auxin-like effects of the natural coumarin scopoletin on Arabidopsis cell structure and morphology. J. Plant Physiol. 2017, 218, 45–55. [Google Scholar] [CrossRef]
- Houssain, M.I.; Qamar-Abbas, S.; Reigosa, M.J. Biological activities and novel applications of secondary metabolite coumarins. Planta Daninha 2017, 35, 1–13. [Google Scholar] [CrossRef]
- Yu, S.-M.; Hu, D.-H.; Zhang, J.-J. Umbelliferone exhibits anticancer activity via the induction of apoptosis and cell cycle arrest in HepG2 hepatocellular carcinoma cells. Mol. Med. Rep. 2015, 12, 3869–3873. [Google Scholar] [CrossRef]
- Ma, Y.M.; Zhou, Y.B.; Xie, C.M.; Chen, D.M.; Li, J. Novel microtubule-targeted agent 6-chloro-4-(methoxyphenyl) coumarin induces G2-M arrest and apoptosis in HeLa cells. Acta Pharmacol. Sin. 2012, 33, 407–417. [Google Scholar] [CrossRef]
- Chuang, J.-Y.; Huang, Y.-F.; Lu, H.-F.; Ho, H.-C.; Yang, J.-S.; Li, T.-M.; Chang, N.-W.; Chung, J.-G. Coumarin induces cell cycle arrest and apoptosis in human cervical cancer HeLa cells through a mitochondria- and caspase-3 dependent mechanism and NF-kappaB down-regulation. In Vivo (Brooklyn) 2007, 21, 1003–1010. [Google Scholar]
- El-Shora, H.M.; El-Gawad, A.M.A. Physiological and biochemical responses of Cucurbita pepo L. Mediated by Portulaca oleracea L. allelopathy. Fresenius Environ. Bull. 2015, 24, 386–393. [Google Scholar]
- Silveira, N.M.; de Oliveira, J.A.; Ribeiro, C.; Canatto, R.A.; Siman, L.; Cambraia, J.; Farnese, F. Nitric Oxide Attenuates Oxidative Stress Induced by Arsenic in Lettuce (Lactuca sativa) Leaves. Water Air Soil. Pollut. 2015, 226, 1–9. [Google Scholar] [CrossRef]
- Esringu, A.; Aksakal, O.; Tabay, D.; Kara, A.A. Effects of sodium nitroprusside (SNP) pretreatment on UV-B stress tolerance in lettuce (Lactuca sativa L.) seedlings. Environ. Sci. Pollut. Res. 2016, 23, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Yan, Z.; He, X.; Li, X.; Qin, B. Allelopathic effect of β-cembrenediol and its mode of action: Induced oxidative stress in lettuce seedlings. Emirates J. Food Agric. 2017, 29, 441–449. [Google Scholar] [CrossRef]
- Duke, S.O. Overview of herbicide mechanisms of action. Environ. Health Perspect. 1990, 87, 263–271. [Google Scholar] [CrossRef]
- Mallory-Smith, C.A.; Retzinger, E.J., Jr. Revised classification of herbicides by site of action for weed resistance management strategies. Weed Technol. 2003, 17, 605–619. [Google Scholar] [CrossRef]
- Reade, J.P.H.; Cobb, A.H. Herbicides: Modes of Action and Metabolism. In Weed Management Handbook, 9th ed.; Naylor, R.E.L., Ed.; Blackwell Science Ltd.: Oxford, UK, 2002; pp. 134–170. [Google Scholar]
- George, E.F. Plant Propagation by Tissue Culture; Springer: Dordrecht, The Netherlands, 2008. [Google Scholar]
- Chiapusio, G.; Sanchez, A.M.; Reigosa, M.J.; Gonzalez, L.; Pellissier, F. Do germination indices adequately reflect allelochemical effects on the germination process? J. Chem. Ecol. 1997, 23, 2445–2453. [Google Scholar] [CrossRef]
- Ribeiro, L.D.O.; Barbosa, S.; Balieiro, F.P.; Beijo, L.A.; Santos, B.R.; Gouvea, C.M.C.; Paiva, L.V. Phytotoxicity of Stryphnodendron adstringens (Mart.) Coville leaf extracts in lettuce bioassay. Rev. Bras. Biociências 2012, 10, 220–225. [Google Scholar]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases: I. Occurrence in Higher Plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Havir, E.A.; McHale, N.A. Biochemical and developmental characterization of multiple forms of catalase in tobacco leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal Lipid Peroxidation. In Methods in Enzymology; U.S. Department of Energy Office of Scientific and Technical Information: Oak Ridge, TN, USA, 1978; Volume 52, pp. 302–310. ISBN 9780121819521. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coumarin | Concentrations (µg mL−1) | |||||
|---|---|---|---|---|---|---|
| 0 | 50 | 100 | 200 | 400 | 800 | |
| A1 | 94.44% a | 91.11% a | 88.89% a | 92.22% a | 86.66% a | 71.11% b |
| A2 | 90.00% a | 91.11% a | 88.89% a | 93.33% a | 97.78% a | 91.11% a |
| A3 | 94.44% a | 96.67% a | 95.55% a | 95.55% a | 92.22% a | 96.67% a |
| A4 | 94.44% a | 84.44% a | 95.55% a | 93.33% a | 88.89% a | 93.33% a |
| A5 | 94.44% a | 94.44% a | 94.44% a | 92.22% a | 91.11% a | 94.44% a |
| A6 | 96.67% a | 93.33% a | 93.33% a | 90.00% a | 91.11% a | 87.77% a |
| Coumarin | Concentrations (µg mL−1) | |||||
|---|---|---|---|---|---|---|
| 0 | 50 | 100 | 200 | 400 | 800 | |
| A1 | 24.66 a | 22.66 a | 14.33 b | 18.50 b | 7.00 b | 0.00 d |
| A2 | 24.50 a | 24.00 a | 25.00 a | 25.50 a | 24.00 a | 15.33 b |
| A3 | 25.50 a | 24.66 a | 24.50 a | 25.50 a | 25.00 a | 7.50 c |
| A4 | 24.33 a | 19.00 b | 24.00 a | 25.33 a | 24.50 a | 18.66 b |
| A5 | 21.66 a | 16.33 b | 22.00 a | 21.66 a | 23.00 a | 23.00 a |
| A6 | 22.66 a | 22.33 a | 25.50 a | 15.00 b | 22.00 a | 1.33 d |
| Coumarin | Concentrations (µg mL−1) | |||||
|---|---|---|---|---|---|---|
| 0 | 50 | 100 | 200 | 400 | 800 | |
| A1 | 0.42 a | 0.27 a | 0.18 c | 0.18 c | 0.15 d | 0.14 c |
| A2 | 0.29 b | 0.36 a | 0.39 a | 0.44 a | 0.45 a | 0.21 b |
| A3 | 0.25 b | 0.25 a | 0.46 a | 0.33 b | 0.37 b | 0.38 a |
| A4 | 0.37 a | 0.24 a | 0.27 b | 0.39 a | 0.33 b | 0.24 b |
| A5 | 0.41 a | 0.29 a | 0.40 a | 0.32 b | 0.25 c | 0.32 a |
| A6 | 0.38 a | 0.32 a | 0.35 a | 0.34 b | 0.30 b | 0.30 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Govêa, K.P.; Pereira, R.S.T.; de Assis, M.D.O.; Alves, P.I.; Brancaglion, G.A.; Toyota, A.E.; Machado, J.V.C.; Carvalho, D.T.; de Souza, T.C.; Beijo, L.A.; et al. Allelochemical Activity of Eugenol-Derived Coumarins on Lactuca sativa L. Plants 2020, 9, 533. https://doi.org/10.3390/plants9040533
Govêa KP, Pereira RST, de Assis MDO, Alves PI, Brancaglion GA, Toyota AE, Machado JVC, Carvalho DT, de Souza TC, Beijo LA, et al. Allelochemical Activity of Eugenol-Derived Coumarins on Lactuca sativa L. Plants. 2020; 9(4):533. https://doi.org/10.3390/plants9040533
Chicago/Turabian StyleGovêa, Kamilla Pacheco, Rafaella Sueko Tomita Pereira, Mateus Donizetti Oliveira de Assis, Pâmela Ingrid Alves, Guilherme Andrade Brancaglion, André Eidi Toyota, José Vaz Cardoso Machado, Diogo Teixeira Carvalho, Thiago Corrêa de Souza, Luiz Alberto Beijo, and et al. 2020. "Allelochemical Activity of Eugenol-Derived Coumarins on Lactuca sativa L." Plants 9, no. 4: 533. https://doi.org/10.3390/plants9040533
APA StyleGovêa, K. P., Pereira, R. S. T., de Assis, M. D. O., Alves, P. I., Brancaglion, G. A., Toyota, A. E., Machado, J. V. C., Carvalho, D. T., de Souza, T. C., Beijo, L. A., Trindade, L. d. O. R., & Barbosa, S. (2020). Allelochemical Activity of Eugenol-Derived Coumarins on Lactuca sativa L. Plants, 9(4), 533. https://doi.org/10.3390/plants9040533