Physical Dormancy Release in Medicago truncatula Seeds Is Related to Environmental Variations

 , ,
, ,  , , and
, , and 
Abstract
1. Introduction
2. Results
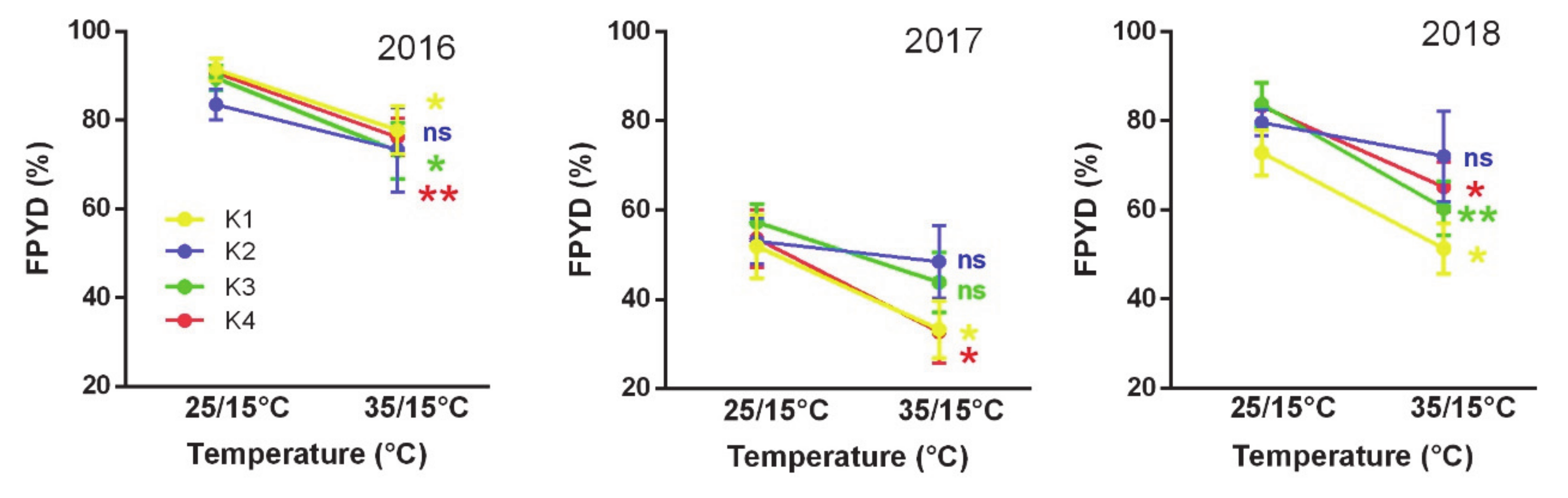
2.1. Responses of Dormancy Traits of Medicago Accessions to Experimental Temperature Treatments
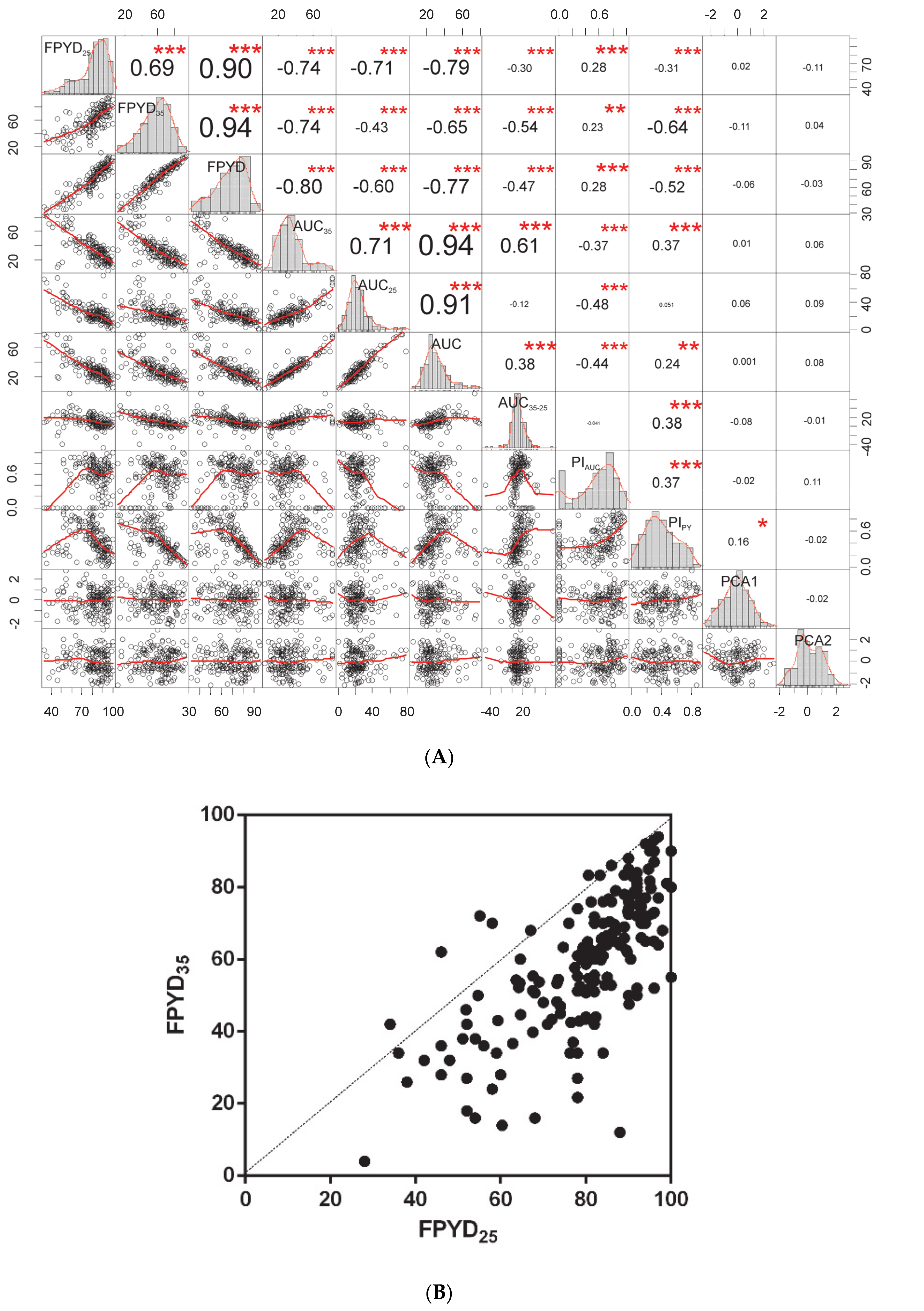
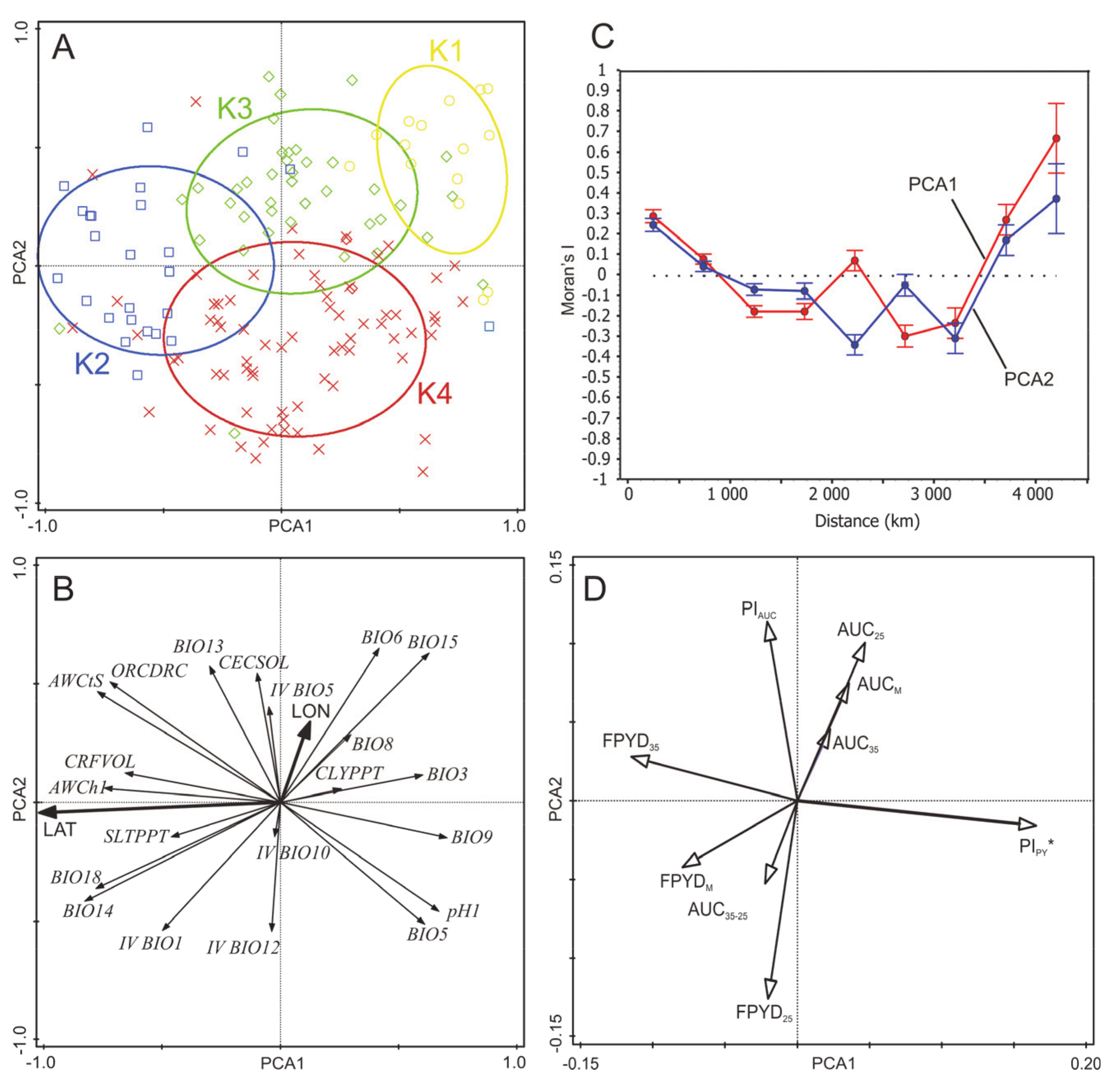
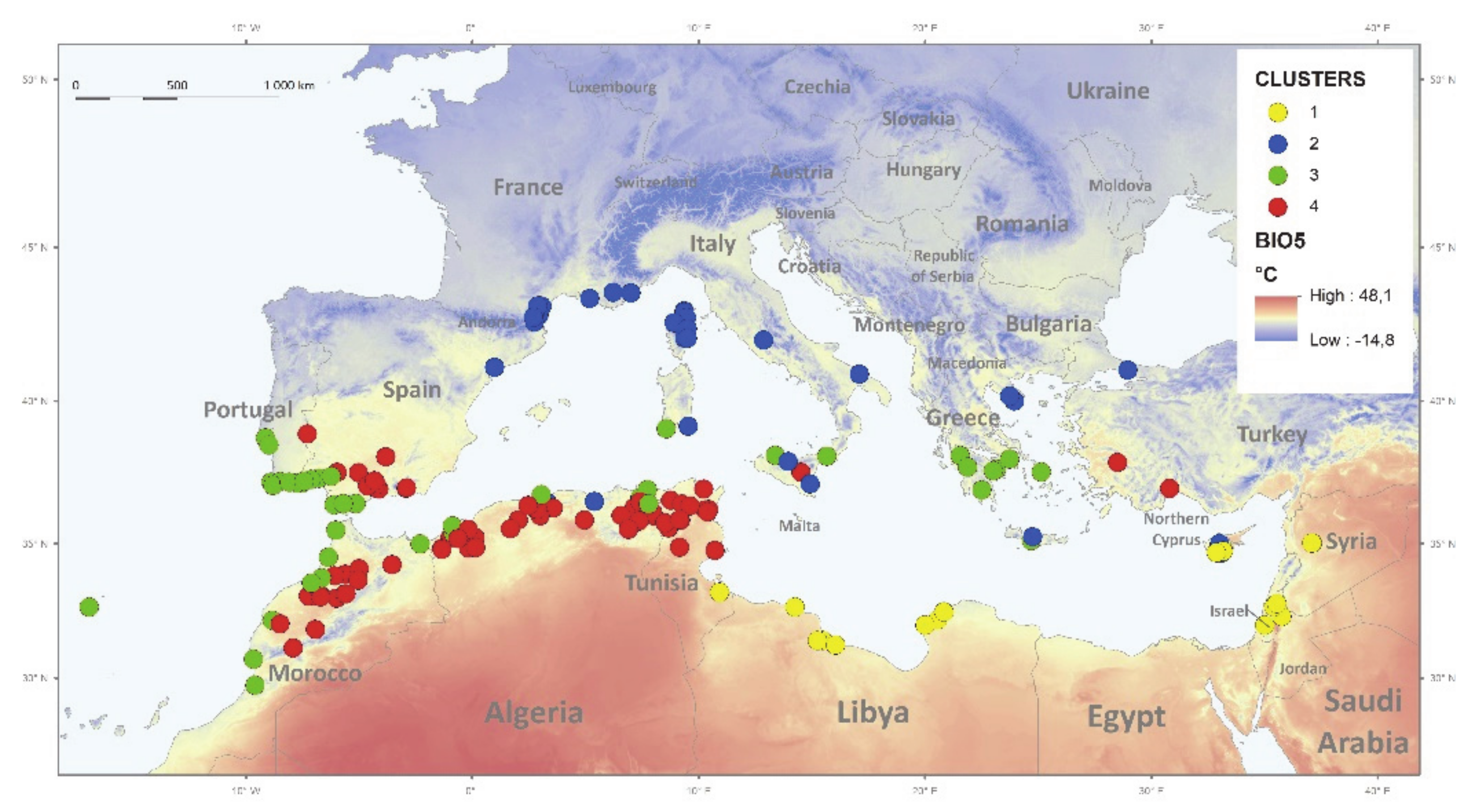
2.2. Associations of Environmental Gradients with Dormancy Traits of Medicago Accessions
2.3. Association Analysis of Dormancy Traits
3. Discussion
3.1. Association between Seed Dormancy Traits and the Environment
3.2. Potential Shortcomings of the Study
3.3. Genetic Basis of Seed Dormancy Release in Legumes
4. Materials and Methods
4.1. Plant Material
4.2. Seed Dormancy Release Experiments
4.3. Evaluation of Dormancy and Germination Traits
4.4. Extraction of Environmental Variables and Spatial Accuracy
4.5. Climate and Soil Characteristics of Localities of Studied Accessions
4.6. Testing of Relationships Among Dormancy Traits, Geography and Environmental Variables
4.7. Phenotypic Plasticity by Macro-Environmental Clusters
4.8. Genome-Wide Association Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PY | physical dormancy |
| FPYD | final PY dormancy at 25/15 °C and 35/15 °C (FPYD25, FPYD35) |
| AUC | area under curve representing germination pattern (AUC25, AUC35) |
| PIAUC | phenotypic plasticity index of the germination pattern |
| PIPY | phenotypic plasticity index of final PY dormancy |
| IV | index of variability |
| FPYDM | means of FPYD coefficients estimated for the two temperature treatments |
| AUCM | means of AUC coefficients estimated for the two temperature treatments |
References
- Tognetti, P.M.; Mazia, N.; Ibáñez, G. Seed local adaptation and seedling plasticity account for Gleditsia triacanthos tree invasion across biomes. Ann. Bot. 2019, 124, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Valladares, F.; Sanchez-Gomez, D.; Zavala, M.A. Quantitative estimation of phenotypic plasticity: Bridging the gap between the evolutionary concept and its ecological applications. J. Ecol. 2006, 94, 1103–1116. [Google Scholar] [CrossRef]
- Forsman, A. Rethinking phenotypic plasticity and its consequences for individuals, populations and species. Heredity 2015, 115, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Clausen, J. Stages in the Evolution of Plant Species; Cornell University Press: Ithaca, NY, USA, 1951. [Google Scholar]
- Ungerer, M.; Johnson, L.; Herman, M. Ecological genomics: Understanding gene and genome function in the natural environment. Heredity 2008, 100, 178–183. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.J.; Mathesius, U.; van Kleunen, M. Plant phenotypic plasticity in a changing climate. Trends. Plant. Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef]
- Richards, C.L.; Bossdorf, O.; Muth, N.Z.; Gurevitch, J.; Pigliucci, M. Jack of all trades, master of some? On the role of phenotypic plasticity in plant invasions. Ecol. Lett. 2006, 9, 981–993. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Seeds: Ecology, Biogeography and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Willis, C.G.; Baskin, C.C.; Baskin, J.M.; Auld, J.R.; Venable, D.L.; Cavender-Bares, J.; Donohue, K.; Rubio de Casas, R. NESCent Germination Working Group. The evolution of seed dormancy: Environmental cues, evolutionary hubs, and diversification of the seed plants. New. Phytol. 2014, 203, 300–309. [Google Scholar] [CrossRef]
- Hradilová, I.; Duchoslav, M.; Brus, J.; Pechanec, V.; Hýbl, M.; Kopecký, P.; Smržová, L.; Štefelová, N.; Vaclávek, T.; Bariotakis, M.; et al. Variation in wild pea (Pisum sativum subsp. elatius) seed dormancy and its relationship to the environment and seed coat traits. PeerJ 2019, 7, e6263. [Google Scholar]
- Norman, H.C.; Cocks, P.C.; Galwey, N.W. Hardseededness in annual clovers: Variation between populations from wet and dry environments. Aus. J. Agric. Sci. 2002, 53, 821–829. [Google Scholar] [CrossRef]
- Rosenberg, M.S.; Anderson, C.D. PASSaGE: Pattern Analysis, Spatial Statistics, and Geographic Exegesis. Version 2. Method. Ecol. Evol. 2011, 2, 229–232. [Google Scholar] [CrossRef]
- Smýkal, P.; Vernoud, V.; Blair, M.W.; Soukup, A.; Thompson, R.D. The role of the testa during development and in establishment of dormancy of the legume seed. Front. Plant. Sci. 2014, 5, 351. [Google Scholar] [PubMed]
- Sultan, S.E. Phenotypic plasticity for plant development, function and life history. Trends. Plant. Sci. 2000, 5, 537–542. [Google Scholar] [CrossRef]
- Templeton, A.R.; Levin, D.A. Evolutionary consequences of seed pools. Am. Nat. 1979, 114, 232–249. [Google Scholar] [CrossRef]
- Evans, M.E.K.; Ferrière, R.; Kane, M.J.; Venable, D.L. Bet hedging via seed banking in desert evening primroses (Oenothera, Onagraceae): Demographic evidence from natural populations. Am. Nat. 2007, 169, 184–194. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Venable, D.L. Bet hedging in a guild of desert annuals. Ecology 2007, 88, 1086–1090. [Google Scholar] [CrossRef] [PubMed]
- Duncan, C.; Schultz, N.L.; Good, M.K.; Lewandrowski, W.; Cook, S. The risk-takers and -avoiders: Germination sensitivity to water stress in an arid zone with unpredictable rainfall. AOB Plants 2019. [Google Scholar] [CrossRef]
- Clauss, M.J.; Venable, D.L. Seed germination in desert annuals: An empirical test of adaptive bet-hedging. Am. Nat. 2000, 155, 168–186. [Google Scholar] [CrossRef]
- Van Klinken, R.D.; Flack, L.K.; Pettit, W. Wet-season Dormancy Release in Seed Banks of a Tropical Leguminous Shrub is Determined by Wet Heat. Ann. Bot. 2006, 98, 875–883. [Google Scholar] [CrossRef]
- Batlla, D.; Benech-Arnold, R.L. A framework for the interpretation of temperature effects on dormancy and germination in seed populations showing dormancy. Seed Sci. Res. 2015, 25, 147–158. [Google Scholar] [CrossRef]
- Ellis, R.H.; Simon, G.; Covell, S. The influence of temperature on seed germination rate in grain legumes. III. A comparison of five faba bean genotypes at constant temperatures using a new screening method. J. Exp. Bot. 1987, 38, 1033–1043. [Google Scholar] [CrossRef]
- Renzi, J.P.; Chantre, G.R.; Cantamutto, M.A. Vicia villosa ssp. villosa Roth field emergence model in a semiarid agroecosystem. Grass. Forage. Sci. 2018, 73, 146–158. [Google Scholar]
- Rosbakh, S.; Poschlod, P. Initial temperature of seed germination as related to species occurrence along a temperature gradient. Funct. Ecol. 2015, 29, 5–14. [Google Scholar] [CrossRef]
- Hudson, A.R.; Ayre, D.J.; Ooi, M.K.J. Physical dormancy in a changing climate. Seed Sci. Res. 2015, 25, 66–81. [Google Scholar] [CrossRef]
- Rubio de Casas, R.; Willis, C.G.; Pearse, W.D.; Baskin, C.C.; Baskin, J.M.; Cavender-Bares, J. Global biogeography of seed dormancy is determined by seasonality and seed size: A case study in the legumes. New. Phytol. 2017, 214, 1527–1536. [Google Scholar] [CrossRef]
- Quinlivan, B.J. The effect of constant and fluctuating temperatures on the permeability of the hard seeds of some legume species. Aust. J. Agric. Res. 1961, 12, 1009–1022. [Google Scholar] [CrossRef]
- Quinlivan, B.J. The relationship between temperature fluctuations and the softening of hard seeds of some legume species. Aust. J. Agric. Res. 1966, 17, 625–631. [Google Scholar] [CrossRef]
- Quinlivan, B.J.; Millington, A.J. The effect of a Mediterranean summer environment on the permeability of hard seeds of subterranean clover. Aust. J. Agric. Res. 1962, 13, 377–387. [Google Scholar] [CrossRef]
- Berger, J.D.; Shrestha, D.; Ludwig, C. Reproductive Strategies in Mediterranean Legumes: Trade-Offs between Phenology, Seed Size and Vigor within and between Wild and Domesticated Lupinus Species Collected along Aridity Gradients. Front. Plant. Sci. 2017, 8, 548. [Google Scholar] [CrossRef]
- Ronfort, J.; Bataillon, T.; Santoni, S.; Delalande, M.; David, J.L.; Prosperi, J.-M. Microsatellite diversity and broad scale geographic structure in a model legume: Building a set of nested core collection for studying naturally occurring variation in Medicago truncatula. BMC Plant Biol. 2006, 6, 28. [Google Scholar] [CrossRef]
- Gallardo, K.; Firnhaber, C.; Zuber, H.; Héricher, D.; Belghazi, M.; Henry, C.; Küster, H.; Thompson, R. A combined proteome and transcriptome analysis of developing Medicago truncatula seeds: Evidence for metabolic specialization of maternal and filial tissues. Mol. Cell. Proteom. 2007, 6, 2165–2179. [Google Scholar] [CrossRef]
- Bolingue, W.; Vu, B.L.; Leprince, O.; Buitink, J. Characterization of dormancy behaviour in seeds of the model legume Medicago truncatula. Seed. Sci. Res. 2010, 20, 97–107. [Google Scholar] [CrossRef]
- Faria, J.M.R.; Buitink, J.; van Lammeren, A.A.M.; Hilhors, H.W.M. Changes in DNA and microtubules during loss and re-establishment of desiccation tolerance in germinating Medicago truncatula seeds. J. Exp. Bot. 2005, 56, 2119–2130. [Google Scholar] [CrossRef] [PubMed]
- Vu, W.T.; Chang, P.L.; Moriuchi, K.S.; Friesen, M.L. Genetic variation of transgenerational plasticity of offspring germination in response to salinity stress and the seed transcriptome of Medicago truncatula. BMC Evol. Biol. 2015, 15, 59. [Google Scholar] [CrossRef] [PubMed]
- Stanton-Geddes, J.; Paape, T.; Epstein, B.; Briskine, R.; Yoder, J.; Mudge, J.; Bharti, A.K.; Farmer, A.D.; Zhou, P.; Denny, R.; et al. Candidate genes and genetic architecture of symbiotic and agronomic traits revealed by whole-genome, sequence-based association genetics in Medicago truncatula. PLoS ONE 2013, 8, e65688. [Google Scholar] [CrossRef] [PubMed]
- Burgarella, C.; Chantret, N.; Gay, L.; Prosperi, J.-M.; Bonhomme, M.; Tiffin, P.; Young, N.D.; Ronfort, J. Adaptation to climate through flowering phenology: A case study in Medicago truncatula. Mol. Ecol. 2016, 25, 3397–3415. [Google Scholar] [CrossRef]
- Julier, B.; Huguet, T.; Chardon, F.; Ayadi, R.; Pierre, J.-B.; Prosperi, J.-M.; Barre, P.; Huyghe, C. Identification of quantitative trait loci influencing aerial morphogenesis in the model legume Medicago truncatula. Appl. Genet. 2007, 114, 1391–1406. [Google Scholar] [CrossRef]
- Thompson, J.D. Plant Evolution in the Mediterranean; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Finch-Savage, W.E.; Footitt, S. Seed dormancy cycling and the regulation of dormancy mechanisms to time germination in variable field environments. J. Exp. Bot. 2017, 68, 843–856. [Google Scholar] [CrossRef]
- Tribouillois, H.; Dürr, C.; Demilly, D.; Wagner, M.-H.; Justes, E. Determination of Germination Response to Temperature and Water Potential for a Wide Range of Cover Crop Species and Related Functional Groups. PLoS ONE 2016, 11, e0161185. [Google Scholar] [CrossRef]
- Hu, X.W.; Fan, Y.; Baskin, C.C.; Baskin, J.M.; Wang, Y.R. Comparison of the effects of temperature and water potential on seed germination of Fabaceae species from desert and subalpine grassland. Am. J. Bot. 2015, 102, 649–660. [Google Scholar] [CrossRef]
- Long, R.L.; Gorecki, M.J.; Renton, M.; Scott, J.K.; Colville, L.; Goggin, D.E.; Commander, L.E.; Westcott, D.A.; Cherry, H.; Finch-Savage, W.E. The ecophysiology of seed persistence: A mechanistic view of the journey to germination or demise. Biol. Rev. Camb. Philos. Soc. 2015, 90, 31–59. [Google Scholar] [CrossRef]
- Ferreras, A.E.; Marcora, P.I.; Venier, M.P.; Funes, G. Different strategies for breaking physical seed dormancy in field conditions in two fruit morphs of Vachellia caven (Fabaceae). Seed. Sci. Res. 2018, 28, 8–15. [Google Scholar] [CrossRef]
- Debieu, M.; Tang, C.; Stich, B.; Sikosek, T.; Effgen, S.; Josephs, E.; Schmitt, J.; Nordborg, M.; Koornneef, M.; Meaux, J. Co-Variation between Seed Dormancy, Growth Rate and Flowering Time Changes with Latitude in Arabidopsis thaliana. PLoS ONE 2013, 8, e61075. [Google Scholar] [CrossRef] [PubMed]
- Postma, F.M.; Ågren, J. Maternal environment affects the genetic basis of seed dormancy in Arabidopsis thaliana. Mol. Ecol. 2015, 24, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Lampei, C.; Metz, J.; Tielbörger, K. Clinal population divergence in an adaptive parental environmental effect that adjusts seed banking. New. Phytol. 2017, 214, 1230–1244. [Google Scholar] [CrossRef]
- Wagmann, K.; Hautekèete, N.-C.; Piquot, Y.; Meunier, C.; Schmitt, S.E.; Van Dijk, H. Seed dormancy distribution: Explanatory ecological factors. Ann. Bot. 2012, 110, 1205–1219. [Google Scholar] [CrossRef]
- Burghardt, L.T.; Metcalf, C.J.E.; Donohue, K. A cline in seed dormancy helps conserve the environment experienced during reproduction across the range of Arabidopsis thaliana. Am. J. Bot. 2016, 103, 47–59. [Google Scholar] [CrossRef]
- Cochrane, A. Multi-year sampling provides insight into the bet-hedging capacity of the soil-stored seed reserve of a threatened Acacia species from Western Australia. Plant. Ecol. 2019, 220, 241–253. [Google Scholar] [CrossRef]
- Ten Brink, H.; Gremer, J.R.; Kokko, H. Optimal germination timing in unpredictable environments: The importance of dormancy for both among- and within-season variation. Ecol. Lett. 2020, 23, 620–630. [Google Scholar] [CrossRef]
- Taylor, G.B. Hardseededness in Mediterranean annual pasture legumes in Australia: A review. Aust. J. Agric. Res. 2005, 56, 645–661. [Google Scholar] [CrossRef]
- Jaganathan, G.K.; Dalrymple, S.E.; Liu, B. Towards an Understanding of Factors Controlling Seed Bank Composition and Longevity in the Alpine Environment. Bot. Rev. 2015, 81, 70–103. [Google Scholar] [CrossRef]
- Tozer, M.G.; Ooi, M.K.J. Humidity-regulated dormancy onset in the Fabaceae: A conceptual model and its ecological implications for the Australian wattle Acacia saligna. Ann. Bot. 2014, 114, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Penfield, S.; MacGregor, D.R. Effects of environmental variation during seed production on seed dormancy and germination. J. Exp. Bot. 2017, 68, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Stine, P.A.; Hunsaker, C.T. An introduction to uncertainty issues for spatial data used in ecological applications. In Spatial Uncertainty in Ecology; Springer: New York, NY, USA, 2001; pp. 91–107. [Google Scholar]
- Brus, J.; Pechanec, V.; Machar, I. Depiction of uncertainty in the visually interpreted land cover data. Ecol. Inf. 2018, 47, 10–13. [Google Scholar] [CrossRef]
- Buisson, L.; Thuiller, W.; Casajus, N.; Lek, S.; Grenouillet, G. Uncertainty in ensemble forecasting of species distribution. Glob. Chang. Biol. 2010, 16, 1145–1157. [Google Scholar] [CrossRef]
- Meyer, C.; Weigelt, P.; Kreft, H. Multidimensional biases, gaps and uncertainties in global plant occurrence information. Ecol. Lett. 2016, 19, 992–1006. [Google Scholar] [CrossRef] [PubMed]
- Kerdaffrec, E.; Nordborg, M. The maternal environment interacts with genetic variation in regulating seed dormancy in Swedish Arabidopsis thaliana. PLoS ONE 2017, 12, e0190242. [Google Scholar] [CrossRef] [PubMed]
- Dias, P.M.B.; Brunel-Muguet, S.; Dürr, C.; Huguet, T.; Demilly, D.; Wagner, M.-H.; Teulat-Merah, B. QTL analysis of seed germination and pre-emergence growth at extreme temperatures in Medicago truncatula. Appl. Genet. 2011, 122, 429–444. [Google Scholar] [CrossRef]
- Kang, Y.; Sakiroglu, M.; Krom, N.; Stanton-Geddes, J.; Wang, M.; Lee, Y.-C.; Young, N.D.; Udvardi, M. Genome-wide association of drought-related and biomass traits with HapMap SNPs in Medicago truncatula. Plant Cell Environ. 2015, 38, 1997–2011. [Google Scholar] [CrossRef]
- Francoz, E.; Lepiniec, L.; North, H.M. Seed coats as an alternative molecular factory: Thinking outside the box. Plant. Reprod. 2018, 31, 327–342. [Google Scholar] [CrossRef]
- Hradilová, I.; Trněný, O.; Válková, M.; Cechová, M.; Janská, A.; Prokešová, L.; Aamir, K.; Krezdorn, N.; Rotter, B.; Winter, P.; Varshney, R.K.; et al. A combined comparative transcriptomic, metabolomic, and anatomical analyses of two key domestication traits: Pod dehiscence and seed dormancy in pea (Pisum. sp.). Front. Plant. Sci. 2017, 8, 542e. [Google Scholar]
- Leubner-Metzger, G. Functions and regulation of β-1,3-glucanases during seed germination, dormancy release and after-ripening. Seed. Sci. Res. 2003, 13, 17–34. [Google Scholar] [CrossRef]
- Leubner-Metzger, G.; Frundt, C.; Vogeli-Lange, R.; Meins, F. Class I [beta]-1,3-Glucanases in the Endosperm of Tobacco during Germination. Plant. Physiol. 1995, 109, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Nighojkar, A.; Srivastava, S.; Kumar, A. Pectin methylesterase from germinating Vigna. sinensis. seeds. Plant. Sci. 1994, 103, 115–120. [Google Scholar] [CrossRef]
- Sitrit, Y.; Hadfield, K.A.; Bennett, A.B.; Bradford, K.J.; Downie, A.B. Expression of a Polygalacturonase Associated with Tomato Seed Germination. Plant. Physiol. 1999, 121, 419–428. [Google Scholar] [CrossRef]
- Buckeridge, M.S.; Hutcheon, I.S.; Reid, J.S.G. The Role of Exo-(1→4)-β-galactanase in the Mobilization of Polysaccharides from the Cotyledon Cell Walls of Lupinus. angustifolius. Following Germination. Ann. Bot. 2005, 96, 435–444. [Google Scholar] [CrossRef]
- Hancock, J.T.; Neill, S.J. Nitric Oxide: Its Generation and Interactions with Other Reactive Signaling Compounds. Plants 2019, 8, 41. [Google Scholar] [CrossRef]
- Müller, K.; Linkies, A.; Vreeburg, R.A.M.; Fry, S.C.; Krieger-Liszkay, A.; Leubner-Metzger, G. In vivo cell wall loosening by hydroxyl radicals during cress seed germination and elongation growth. Plant. Physiol. 2009, 150, 1855–1865. [Google Scholar] [CrossRef]
- Jeevan Kumar, S.P.; Rajendra Prasad, S.; Banerjee, R.; Thammineni, C. Seed birth to death: Dual functions of reactive oxygen species in seed physiology. Ann. Bot. 2015, 116, 663–668. [Google Scholar] [CrossRef]
- Raviv, B.; Godwin, J.; Granot, G.; Grafi, G. The Dead Can Nurture: Novel Insights into the Function of Dead Organs Enclosing Embryos. Int. J. Mol. Sci. 2018, 19, 2455. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy; Springer: New York, NY, USA, 2013. [Google Scholar]
- Novo-Uzal, E.; Fernández-Pérez, F.; Herrero, J.; Gutiérrez, J.; Gómez-Ros, L.V.; Bernal, M.Á.; Díaz, J.; Cuello, J.; Pomar, F.; Pedreño, M.Á. From Zinnia to Arabidopsis: Approaching the involvement of peroxidases in lignification. J. Exp. Bot. 2013, 64, 3499–3518. [Google Scholar] [CrossRef]
- Pollard, A.T. Seeds vs fungi: An enzymatic battle in the soil seedbank. Seed. Sci. Res. 2018, 28, 197–214. [Google Scholar] [CrossRef]
- Graeber, K.; Nakabayashi, K.; Miatton, E.; Leubner-Metzger, G.; Soppe, W.J. Molecular mechanisms of seed dormancy. Plant. Cell. Environ. 2012, 35, 1769–86. [Google Scholar] [CrossRef] [PubMed]
- Yoder, J.B.; Stanton-Geddes, J.; Zhou, P.; Briskine, R.; Young, N.D.; Tiffin, P. Genomic signature of adaptation to climate in Medicago truncatula. Genetics 2014, 196, 1263–1275. [Google Scholar] [CrossRef] [PubMed]
- De Boor, C. A Practical Guide to Splines; Springer: New York, NY, USA, 1978. [Google Scholar]
- Machalová, J. Optimal interpolating and optimal smoothing spline. J. Elect. Eng. 2002, 5312, 79–82. [Google Scholar]
- Kader, M.A. A Comparison of Seed Germination Calculation Formulae and the Associated Interpretation of Resulting Data. J. Proc. R. Soc. New. South. Wales. 2005, 138, 65–75. [Google Scholar]
- Talská, R.; Machalová, J.; Smýkal, P.; Hron, K. A comparison of seed germination coefficients using functional regression. Appl. Plant. Sci. 2020, (in press).
- ESRI. Available online: https://pro.arcgis.com (accessed on 5 October 2019).
- WorldClim. Available online: http://worldclim.org (accessed on 5 October 2019).
- Naimi, B. Uncertainty Analysis for Species Distribution Models. Version 1.1-18. 2017. Available online: http://r-gis.net (accessed on 5 October 2019).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Cobon, D.H.; Kouadio, L.; Mushtaq, S.; Jarvis, C.; Carter, J.; Stone, G.; Davis, P. Evaluating the shifts in rainfall and pasture-growth variabilities across the pastoral zone of Australia during 1910–2010. Crop. Pasture. Sci. 2019, 70, 634–647. [Google Scholar] [CrossRef]
- Vega, G.C.; Pertierra, L.R.; Olalla-Tarraga, M.A. MERRAclim, a high-resolution global dataset of remotely sensed bioclimatic variables for ecological modelling. Sci. Data 2018, 5, 170078. [Google Scholar] [CrossRef]
- ISRIC. Available online: https://www.isric.org/explore/soilgrids (accessed on 5 October 2019).
- Hengl, T.; de Mendes, J.J.; Heuvelink, G.B.M.; Ruiperez Gonzalez, M.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B.; et al. SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef]
- Peterson, B.G. Econometric Tools for Performance and Risk Analysis. 2019. Available online: https://github.com/braverock/PerformanceAnalytics (accessed on 10 October 2019).
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2012; p. 990. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, version 5.0; Microcomputer Power: Ithaca, NY, USA, 2012; p. 496. [Google Scholar]
- Dutilleul, P.; Clifford, P.; Richardson, S.; Hemon, D. Modifying the t test for assessing the correlation between two spatial processes. Biometrics 1993, 49, 305–314. [Google Scholar] [CrossRef]
- Rangel, T.F.; Diniz-Filho, J.A.F.; Bini, L.M. SAM: A comprehensive application for Spatial Analysis in Macroecology. Ecography 2010, 33, 46–50. [Google Scholar] [CrossRef]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Version 2013; Grupo InfoStat, FCA; Universidad Nacional de Córdoba: Córdoba, Argentina, 2013. [Google Scholar]
- Yu, J.; Buckler, E.S. Genetic association mapping and genome organization of maize. Curr. Opin. Biotech. 2006, 17, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative Usage of Fixed and Random Effect Models for Powerful and Efficient Genome-Wide Association Studies. PLoS Genet. 2016, 16, e1005767. [Google Scholar] [CrossRef] [PubMed]
- Pecrix, Y.; Staton, S.E.; Sallet, E.; Lelandais-Brière, C.; Moreau, S.; Carrère, S.; Zahm, M.; Kreplak, J.; Mayjonade, B.; Carine Satgé, C.; et al. Whole-genome landscape of Medicago. truncatula. symbiotic genes. Nat. Plants. 2018, 4, 1017–1025. [Google Scholar] [CrossRef]
- Bonhomme, M.; André, O.; Badis, Y.; Ronfort, J.; Burgarella, C.; Chantret, N.; Miteul, H.; Hajri, A.; Baranger, A.; Tiffin, P.; et al. High-density genome-wide association mapping implicates an F-box encoding gene in Medicago truncatula resistance to Aphanomyces euteiches. New. Phytol. 2014, 201, 1328–1342. [Google Scholar]
- Medicago Truncatula A17 r5.0 Genome Portal. 2019. Available online: https://medicago.toulouse.inra.fr/MtrunA17r5.0-ANR (accessed on 15 December 2019).
- Branca, A.; Paape, T.D.; Zhou, P.; Briskine, R.; Farmer, A.D.; Mudge, J.; Ben, C.; Denny, R.; Sadowsky, M.J.; Ronfort, J.; et al. Whole-genome nucleotide diversity, recombination, and linkage disequilibrium in the model legume Medicago truncatula. Proc. Nat. Acad. Sci. USA 2011, 108, E864–E870. [Google Scholar]
- Fu, F.; Zhang, W.; Li, Y.Y.; Wang, H.L. Establishment of the model system between phytochemicals and gene expression profiles in Macrosclereid cells of Medicago truncatula. Sci. Rep. 2017, 7, 2580. [Google Scholar] [CrossRef]
- Verdier, J.; Dessaint, F.; Schneider, C.; Abirached-Darmency, M. A combined histology and transcriptome analysis unravels novel questions on Medicago truncatula seed coat. J. Exp. Bot. 2013, 64, 459–470. [Google Scholar] [CrossRef]
- Bar. University of Toronto. Available online: http://bar.utoronto.ca/efpmedicago/cgi-bin/efpWeb.cgi (accessed on 15 December 2019).
- Gentzbittel, L.; Ben, C.; Mazurier, M.; Shin, M.-G.; Lorenz, T.; Rickauer, M.; Marjoram, P.; Nuzhdin, S.V.; Tatarinova, T.V. WhoGEM: An admixture-based prediction machine accurately predicts quantitative functional traits in plants. Genome. Biol. 2019, 20, 106. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chrom | Position of QTN | P-value | Candidate gene ID within +/- 10kb interval (Mtv5) | Gene ID v4 | Gene annotation | Gene description |
|---|---|---|---|---|---|---|
| 1 | 34656783 | 4.25 × 10−11 | MtrunA17Chr1g0184131 | Medtr1g070110 | Hyoscyamine 6-dioxygenase | secondary metabolism.flavonoids.dihydroflavonols |
| 1 | 49585766 | 1.36 × 10−7 | MtrunA17Chr1g0204011 | Medtr1g101830 | Peroxidase | misc.peroxidases |
| 1 | 50761951 | 2.79 × 10−7 | MtrunA17Chr1g0205571 | Medtr1g104590 | Primary-amine oxidase | misc.oxidases - copper, flavone etc |
| 1 | 50761951 | 4.23 × 10−8 | MtrunA17Chr1g0205541 | Medtr1g104550 | Primary-amine oxidase | misc.oxidases - copper, flavone etc |
| 2 | 10895258 | 5.60 × 10−9 | MtrunA17Chr2g0291751 | Medtr2g028980 | Peroxidase | misc.peroxidases |
| 2 | 12184616 | 5.76 × 10−9 | MtrunA17Chr2g0293411 | Medtr2g031920 | Ent-kaurenoic acid oxidase 2 | hormone metabolism.gibberelin.synthesis-degradation.ent-kaurenoic acid hydroxylase/oxygenase |
| 4 | 53502844 | 5.22 × 10−8 | MtrunA17Chr4g0061311 | Medtr4g109360 | UDP-glucose 6-dehydrogenase | cell wall.precursor synthesis.UDP-Glc dehydrogenase (UGD) |
| 4 | 53502844 | 3.62 × 10−8 | MtrunA17Chr4g0061381 | Medtr4g109470 | Flavonoid 3’-monooxygenase | secondary metabolism.flavonoids.dihydroflavonols.flavonoid 3-monooxygenase |
| 4 | 61245281 | 2.56 × 10−7 | MtrunA17Chr4g0072091 | Medtr4g127670 | Peroxidase | misc.peroxidases |
| 5 | 4768980 | 2.23 × 10−12 | MtrunA17Chr5g0400421 | Medtr5g014250 | Beta-amyrin 11-oxidase-like | hormone metabolism.gibberelin.synthesis-degradation.ent-kaurenoic acid hydroxylase/oxygenase |
| 5 | 33045218 | 2.41 × 10−7 | MtrunA17Chr5g0432331 | Medtr5g074710 | Peroxidase | misc.peroxidases |
| 5 | 33045218 | 2.41 × 10−7 | MtrunA17Chr5g0432361 | Medtr5g074770 | Peroxidase | misc.peroxidases |
| 6 | 28332981 | 4.21 × 10−10 | MtrunA17Chr6g0474391 | Medtr6g464620 | Gibberellin 3-beta-dioxygenase | hormone metabolism.gibberelin.synthesis-degradation.GA20 oxidase |
| 6 | 34330903 | 7.87 × 10−6 | MtrunA17Chr6g0479681 | Medtr6g072490 | Cytokinin hydroxylase-like | misc.cytochrome P450 |
| 8 | 6442861 | 1.76 × 10−9 | MtrunA17Chr8g0343001 | Medtr8g018650 | Seed linoleate 9S-lipoxygenase | hormone metabolism.jasmonate.synthesis-degradation.lipoxygenase |
| 8 | 48706047 | 1.32 × 10−9 | MtrunA17Chr8g0391921 | Medtr8g105630 | Glutathione peroxidase | redox.ascorbate and glutathione.glutathione |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Renzi, J.P.; Duchoslav, M.; Brus, J.; Hradilová, I.; Pechanec, V.; Václavek, T.; Machalová, J.; Hron, K.; Verdier, J.; Smýkal, P. Physical Dormancy Release in Medicago truncatula Seeds Is Related to Environmental Variations. Plants 2020, 9, 503. https://doi.org/10.3390/plants9040503
Renzi JP, Duchoslav M, Brus J, Hradilová I, Pechanec V, Václavek T, Machalová J, Hron K, Verdier J, Smýkal P. Physical Dormancy Release in Medicago truncatula Seeds Is Related to Environmental Variations. Plants. 2020; 9(4):503. https://doi.org/10.3390/plants9040503
Chicago/Turabian StyleRenzi, Juan Pablo, Martin Duchoslav, Jan Brus, Iveta Hradilová, Vilém Pechanec, Tadeáš Václavek, Jitka Machalová, Karel Hron, Jerome Verdier, and Petr Smýkal. 2020. "Physical Dormancy Release in Medicago truncatula Seeds Is Related to Environmental Variations" Plants 9, no. 4: 503. https://doi.org/10.3390/plants9040503
APA StyleRenzi, J. P., Duchoslav, M., Brus, J., Hradilová, I., Pechanec, V., Václavek, T., Machalová, J., Hron, K., Verdier, J., & Smýkal, P. (2020). Physical Dormancy Release in Medicago truncatula Seeds Is Related to Environmental Variations. Plants, 9(4), 503. https://doi.org/10.3390/plants9040503