Additive Effect of Botanical Insecticide and Entomopathogenic Fungi on Pest Mortality and the Behavioral Response of Its Natural Enemy

 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Aphid Mortality Assay
2.1.1. Survival
2.1.2. Hyphal Growth on Insect Surface
2.1.3. Number of Offspring
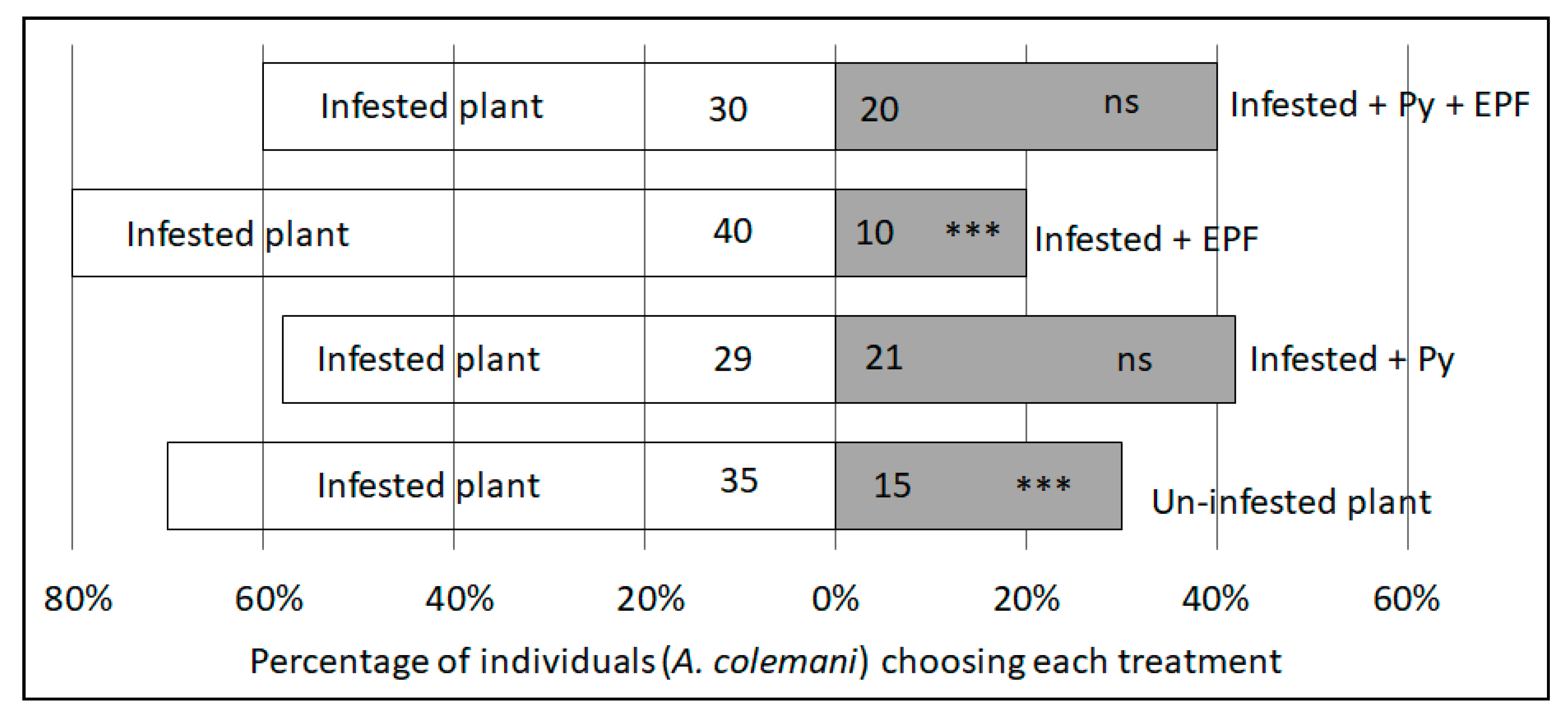
2.2. Parasitoid Dual-Choice Assays
3. Discussion
- Efficacy of pyrethrum and EPF would be enhanced when presented in combination.
- The biopesticides would affect parasitoid plant/host preference.
4. Materials and Methods
4.1. Insect Rearing
4.2. Entomopathogenic Fungi
4.3. Preperation of Pyrethrum
4.4. Mortality Assays
4.5. Parasitoid Choice Assays
4.6. Statistical Analyses
4.6.1. Survival
4.6.2. Visible Fungal Growth
4.6.3. Number of Offspring
4.6.4. Parasitoid Dual-Choice Assays
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Belmain, S.R.; Stevenson, P.C. Ethnobotanicals in Ghana: Reviving and modernising an age-old practise. Pestic Outlook 2001, 12, 233–238. [Google Scholar]
- Tembo, Y.; Mkindi, A.G.; Mkenda, P.A.; Mpumi, N.; Mwanauta, R.; Stevenson, P.C.; Ndakidemi, P.A.; Belmain, S.R. Pesticidal Plant Extracts Improve Yield and Reduce Insect Pests on Legume Crops Without Harming Beneficial Arthropods. Front. Plant Sci. 2018, 9, 1425. [Google Scholar] [CrossRef] [PubMed]
- Mkenda, P.; Mwanauta, R.; Stevenson, P.C.; Ndakidemi, P.; Mtei, K.; Belmain, S.R. Extracts from Field Margin Weeds Provide Economically Viable and Environmentally Benign Pest Control Compared to Synthetic Pesticides. PLoS ONE 2015, 10, 0143530. [Google Scholar] [CrossRef] [PubMed]
- Deng, A.L.; Ogendo, J.O.; Owuor, G.; Bett, P.K.; Omolo, E.O.; Mugisha-Kamatenesi, M.; Mihale, J.M. Factors determining the use of botanical insect pest control methods by small-holder farmers in the Lake Victoria basin Kenya. AJEST 2009, 3, 108–115. [Google Scholar]
- Grzywacz, D.; Stevenson, P.C.; Mushobozi, W.L.; Belmain, S.; Wilson, K. The use of indigenous ecological resources for pest control in Africa. Food Secur. 2014, 6, 71–86. [Google Scholar] [CrossRef]
- Cox, C. Pyrethrum/pyrethrin insecticide fact sheet. J. Pestic. Reform 2002, 22, 14–20. [Google Scholar]
- Schleier, J.J., III; Peterson, R.K.D. Pyrethrins and pyrethroid insecticides. In Green Trends in Insect Control. Lopez O.; Fernandez-Bolanos, J., Ed.; RSC: London, UK, 2011; Volume 3, pp. 94–131. [Google Scholar]
- Casida, J.E. Pyrethrum flowers and pyrethroid insecticides. Environ. Health Perspect. 1980, 34, 189–202. [Google Scholar] [CrossRef]
- Sola, P.; Mvumi, B.M.; Ogendo, J.O.; Mponda, O.; Kamanula, J.F.; Nyirenda, S.P.; Belmain, S.R.; Stevenson, P.C. Botanical pesticide production, trade and regulatory mechanisms in sub-Saharan Africa: Making a case for plant-based pesticidal products. Food Secur. 2014, 6, 369–384. [Google Scholar] [CrossRef]
- Gallo, M.; Formato, A.; Ianniello, D.; Andolfi, A.; Conte, E.; Ciaravolo, M.; Varchetta, V.; Naviglio, D. Supercritical fluid extraction of pyrethrins from pyrethrum flowers (Chrysanthemum cinerariifolium) compared to traditional maceration and cyclic pressurization extraction. J. Supercrit. Fluid 2017, 119, 104–112. [Google Scholar] [CrossRef]
- Chandler, D.; Bailey, A.; Tatchell, M.; Davidson, G.; Greaves, J.; Grant, W. The development, regulation and use of biopesticides for integrated pest management. Philos. Trans. R. Soc. B 2011, 366, 1987–1998. [Google Scholar] [CrossRef]
- Marrone, P. The Market and Potential for Biopesticides Biopesticides. ACS Symp. Ser. Am. Chem. Soc. 2014, 1172, 245–258. [Google Scholar]
- Damalas, C.A.; Koutroubas, S.D. Current Status and Recent Developments in Biopesticide Use. Agriculture 2018, 8, 13. [Google Scholar] [CrossRef]
- Chen, M.; Du, Y.; Zhu, G.; Takamatsu, G.; Ihara, M.; Matsuda, K.; Zhorov, B.; Dong, K. Action of six pyrethrins purified from the botanical insecticide pyrethrum on cockroach sodium channels expressed in Xenopus oocytes. Pestic. Biochem. Physiol. 2018, 151, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Davies, T.G.E.; Field, L.M.; Usherwood, P.N.R.; Williamson, M.S. DDT, pyrethrins, pyrethroids and insect sodium channels. IUBMB Life 2007, 59, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Oruonye, E.D.; Okrikata, E. Sustainable use of plant protection products in Nigeria and challenges. J. Plant Breed. Crop Sci. 2010, 2, 267–272. [Google Scholar]
- Mpumi, N.; Mtei, K.; Machunda, R.; Ndakidemi, P.A. The Toxicity, Persistence and Mode of Actions of Selected Botanical Pesticides in Africa against Insect Pests in Common Beans, P. vulgaris: A Review. Am. J. Plant Sci. 2016, 7, 138–151. [Google Scholar] [CrossRef]
- Karani, A.O.; Ndakidemi, P.A.; Mbega, E.R. Botanical Pesticides in Management of Common Bean Pests: Importance and Possibilities for Adoption by Small-scale Farmers in Africa. J. Appl. Life Sci. 2017, 12, 1–10. [Google Scholar] [CrossRef]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef]
- Inglis, D.G.; Gottel, M.S.; Butt, T.M.; Strasser, H. Use of Hyphomycetous Fungi for Managing Insect Pests. In Fungi as Biocontrol Agents: Progress, Problems and Potential; Butt, T.M., Jackson, C., Magan, N., Eds.; CAB International: Wallingford, UK, 2001; pp. 23–69. [Google Scholar]
- Sinha, K.K.; Choudhary, A.K.; Priyanka, K. Chapter 15-Entomopathogenic Fungi. In Ecofriendly Pest Management for Food Security; Omkar, Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 475–505. [Google Scholar]
- Steinkraus, D.C. Factors affecting transmission of fungal pathogens of aphids. J. Invertebr. Pathol. 2006, 92, 1–41. [Google Scholar] [CrossRef]
- Meyling, N.; Eilenberg, J. Ecology of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in temperate agroecosystems: Potential for conservation biological control. Biol. Control. 2007, 43, 145–155. [Google Scholar] [CrossRef]
- Islam, M.T.; Omar, D.; Latif, M.A.; Morshed, M.M. The integrated use of entomopathogenic fungus, Beauveria bassiana with botanical insecticide, neem against Bemisia tabaci on eggplant. Afr. J. Microbiol. Res. 2011, 5, 3409–3413. [Google Scholar]
- Islam, M.T.; Omar, D. Combined effect of Beauveria bassiana with neem on virulence of insect in case of two application approaches. J. Anim. Plant Sci. 2012, 22, 77–82. [Google Scholar]
- Ribeiro, L.P.; Blume, E.; Bogorni, P.C.; Dequech, S.T.B.; Brand, S.C.; Junges, E. Compatibility of Beauveria bassiana commercial isolate with botanical insecticides utilized in organic crops in southern Brazil. Biol. Agric. Hortic. 2012, 28, 223–240. [Google Scholar] [CrossRef]
- Shoukat, R.F.; Freed, S.; Ahmad, K.W. Evaluation of binary mixtures of entomogenous fungi and botanicals on biological parameters of Culex pipiens (Diptera: Culicidae) under laboratory and field conditions. Int. J. Mosq. Res. 2016, 3, 17–24. [Google Scholar]
- Ludwig, S.W.; Oetting, R.D. Susceptibility of Natural Enemies to Infection by Beauveria bassiana and Impact of Insecticides on Ipheseius degenerans (Acari: Phytoseiidae). J. Agric. Urban Entomol. 2001, 18, 169–178. [Google Scholar]
- Shipp, J.L.; Zhang, Y.; Hunt, D.W.A.; Ferguson, G. Influence of Humidity and Greenhouse Microclimate on the Efficacy of Beauveria bassiana (Balsamo) for Control of Greenhouse Arthropod Pests. Environ. Entomol. 2003, 32, 1154–1163. [Google Scholar] [CrossRef]
- Rashkia, M.; Kharazi-pakdel, A.; Allahyari, H.; van Alphen, J.J.M. Interactions among the entomopathogenic fungus, Beauveria bassiana (Ascomycota: Hypocreales), the parasitoid, Aphidius matricariae (Hymenoptera: Braconidae), and its host, Myzus persicae (Homoptera: Aphididae). Biol. Control 2009, 50, 324–328. [Google Scholar] [CrossRef]
- Martins, I.C.F.; Silva, R.J.; Alencar, J.R.D.C.C.; Silva, K.P.; Cividanes, F.J.; Duarte, R.T.; Agostini, L.T.; Polanczyk, R.A. Interactions Between the Entomopathogenic Fungi Beauveria bassiana (Ascomycota: Hypocreales) and the Aphid Parasitoid Diaeretiella rapae (Hymenoptera: Braconidae) on Myzus persicae (Hemiptera: Aphididae). J. Econ. Entomol. 2014, 107, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, M.M.; Hatcher, P.E. Combining entomopathogenic fungi and parasitoids to control the green peach aphid Myzus persicae. Biol. Control 2017, 110, 44–55. [Google Scholar] [CrossRef]
- Ali, S.; Farooqi, M.A.; Sajjad, A. Compatibility of entomopathogenic fungi and botanical extracts against the wheat aphid, Sitobion avenae (Fab.) (Hemiptera: Aphididae). Egypt. J. Biol. Pest Control 2018, 28, 97. [Google Scholar] [CrossRef]
- Otieno, J.A.; Pallmann, P.; Poehling, H.M. Additive and synergistic interactions amongst Orius laevigatus (Heteroptera: Anthocoridae), entomopathogens and azadirachtin for controlling western flower thrips (Thysanoptera: Thripidae). BioControl 2017, 62, 85–95. [Google Scholar] [CrossRef]
- Johnson, D.J.; Goettel, M.S. Reduction of grasshopper populations following field application of the fungus Beauveria bassiana. Biocontrol Sci. Technol. 1992, 3, 165–175. [Google Scholar] [CrossRef]
- Jaros-Su, J.; Groden, E.; Zhang, J. Effects of Selected Fungicides and the Timing of Fungicide Application on Beauveria bassiana-Induced Mortality of the Colorado Potato Beetle (Coleoptera: Chrysomelidae). BioControl 1999, 15, 259–269. [Google Scholar]
- Benjamin, M.A.; Zhioua, E.; Ostfeld, R.S. Laboratory and Field Evaluation of the Entomopathogenic Fungus Metarhizium anisopliae (Deuteromycetes) for Controlling Questing Adult Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 2002, 39, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Peng, G.; Wang, W.; Yin, Y.; Zeng, D.; Xia, Y. Field trials of Metarhizium anisopliae var. acridum (Ascomycota: Hypocreales) against oriental migratory locusts, Locusta migratoria manilensis (Meyen) in Northern China. Crop. Prot. 2008, 27, 1244–1250. [Google Scholar] [CrossRef]
- Ormond, E.L.; Alison, P.M.T.; Pell, J.K.; Freeman, S.N.; Roy, H.E. Avoidance of a generalist entomopathogenic fungus by the ladybird, Coccinella septempunctata. FEMS Microbiol. Ecol. 2011, 77, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Meyling, N.V.; Pell, J.K. Detection and avoidance of an entomopathogenic fungus by a generalist insect predator. Ecol. Entomol. 2006, 31, 162–171. [Google Scholar] [CrossRef]
- Lord, J.C. Response of the wasp, Cephalonomia tarsalis (Hymenoptera: Bethylidae) to Beauveria bassiana (Hyphomycetes: Moniliales) as free conidia or infection in its host, the saw-toothed grain beetle, Oryzaephilus surinamensis (Coleoptera: Silvanidae). Biol. Control 2001, 21, 300–304. [Google Scholar] [CrossRef]
- Rajapakse, R.H.S. Effect of host age, parasitoid age, and temperature on interspecific competition between Chelonus insularis Cresso, Cotesia marginiventris Cresson and Microplitis manilae Ashmead. Int. J. Trop. Insect Sci. 1992, 13, 87–94. [Google Scholar] [CrossRef]
- Silva-Torres, C.S.A.; Barros, R.; Torres, J.B. Effect of age, photoperiod and host availability on the parasitism behaviour of Oomyzus sokolowskii Kurdjumov (Hymenoptera: Eulophidae). Neotrop. Entomol. 2009, 38, 512–519. [Google Scholar] [CrossRef]
- Pizzol, J.; Desneux, N.; Wajnberg, E.; Thiéry, D. Parasitoid and host egg ages have independent impact on various biological traits in a Trichogramma species. J. Pest Sci. 2012, 85, 489–496. [Google Scholar] [CrossRef]
- Guerrieri, E.; Poppy, G.M.; Powell, W.; Tremblay, W.; Pennacchio, F. Induction and Systemic Release of Herbivore-Induced Plant Volatiles Mediating In-Flight Orientation of Aphidius ervi. J. Chem. Ecol. 1999, 25, 1247–1261. [Google Scholar] [CrossRef]
- Crawley, M.J. The R Book, 2nd ed.; Wiley: Chichester, UK, 2013; pp. 869–892. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model.; Springer: New York, NY, USA, 2000; pp. 1–350. [Google Scholar]
- Therneau, T. A Package for Survival Analysis in S. version 2.38. 2015. Available online: https://CRAN.R-project.org/package=survival. (accessed on 25 December 2019).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Length, R. emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.3.3. 2019. Available online: https://cran.r-project.org/web/packages/emmeans/index.html (accessed on 25 December 2019).




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Grandon, G.M.; Harte, S.J.; Ewany, J.; Bray, D.; Stevenson, P.C. Additive Effect of Botanical Insecticide and Entomopathogenic Fungi on Pest Mortality and the Behavioral Response of Its Natural Enemy. Plants 2020, 9, 173. https://doi.org/10.3390/plants9020173
Fernández-Grandon GM, Harte SJ, Ewany J, Bray D, Stevenson PC. Additive Effect of Botanical Insecticide and Entomopathogenic Fungi on Pest Mortality and the Behavioral Response of Its Natural Enemy. Plants. 2020; 9(2):173. https://doi.org/10.3390/plants9020173
Chicago/Turabian StyleFernández-Grandon, G. Mandela, Steven J. Harte, Jaspher Ewany, Daniel Bray, and Philip C. Stevenson. 2020. "Additive Effect of Botanical Insecticide and Entomopathogenic Fungi on Pest Mortality and the Behavioral Response of Its Natural Enemy" Plants 9, no. 2: 173. https://doi.org/10.3390/plants9020173
APA StyleFernández-Grandon, G. M., Harte, S. J., Ewany, J., Bray, D., & Stevenson, P. C. (2020). Additive Effect of Botanical Insecticide and Entomopathogenic Fungi on Pest Mortality and the Behavioral Response of Its Natural Enemy. Plants, 9(2), 173. https://doi.org/10.3390/plants9020173