Some Mechanisms Modulating the Root Growth of Various Wheat Species under Osmotic-Stress Conditions

 ,
,  ,
,
Abstract

1. Introduction
2. Results
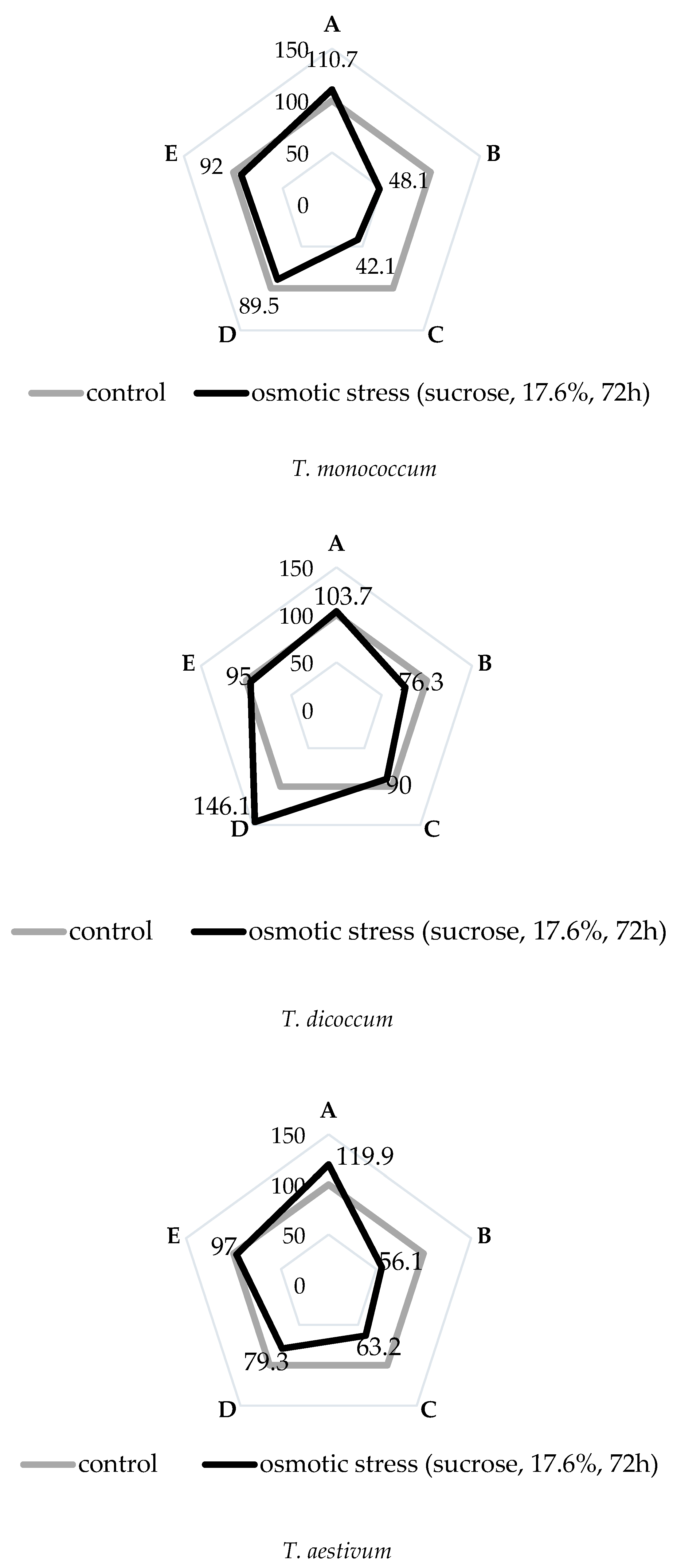
2.1. Morphophysiological Reactions of Primary Roots of Different Wheat Types to Osmotic Stress
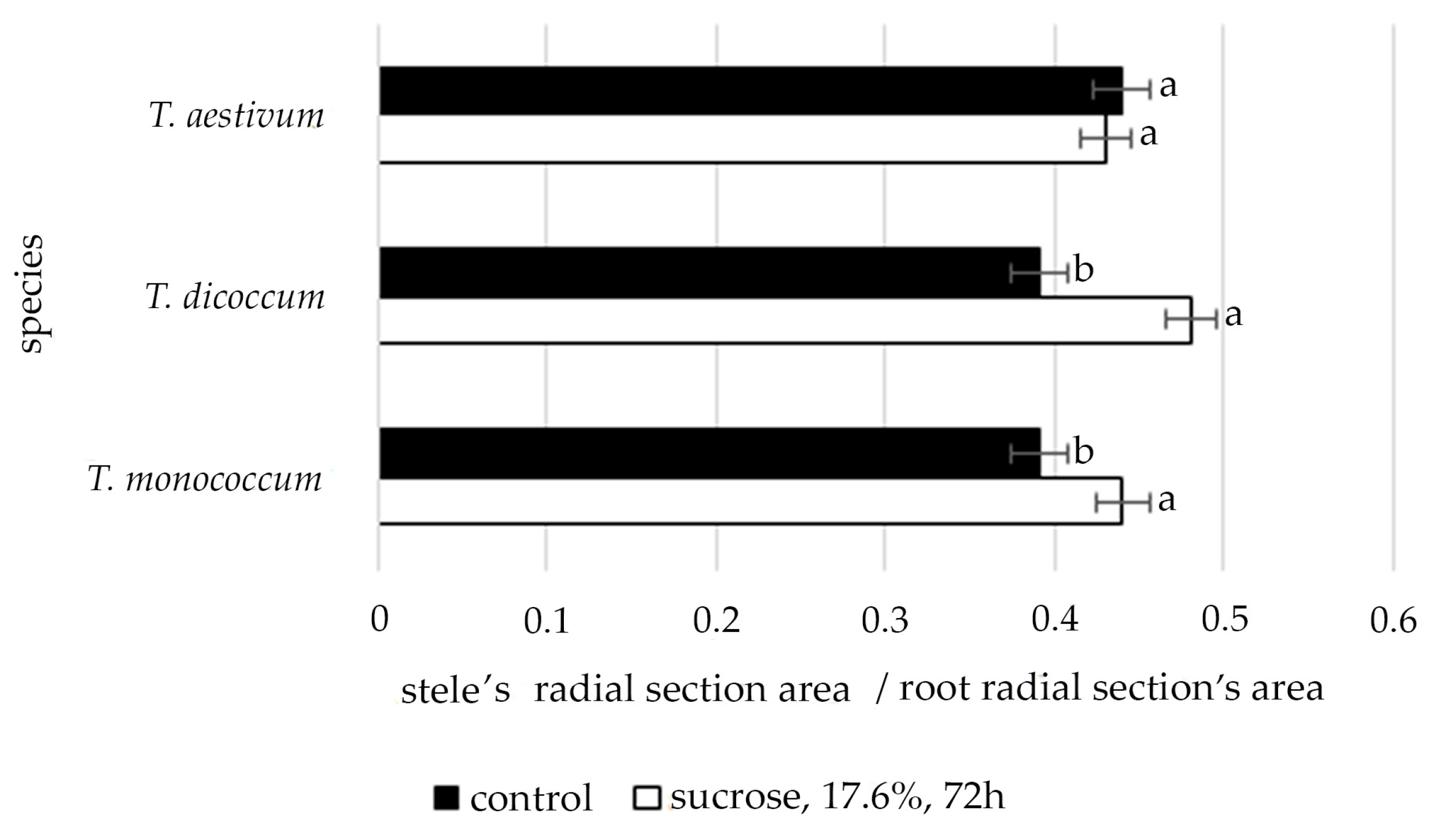
2.2. Changes in the Elements of the Anatomical Structure of Different Wheat Species’ Primary Roots during Osmotic Stress
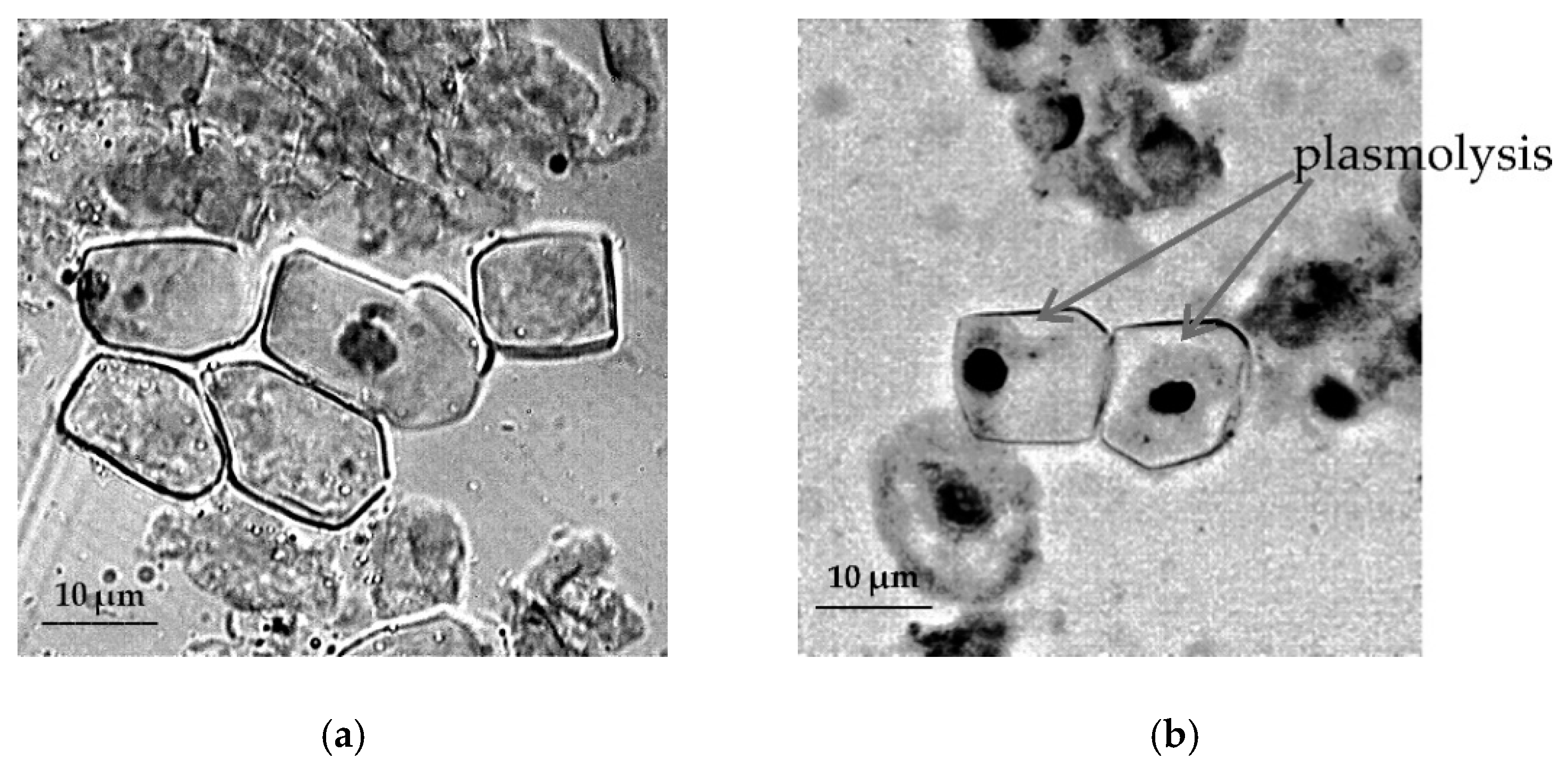
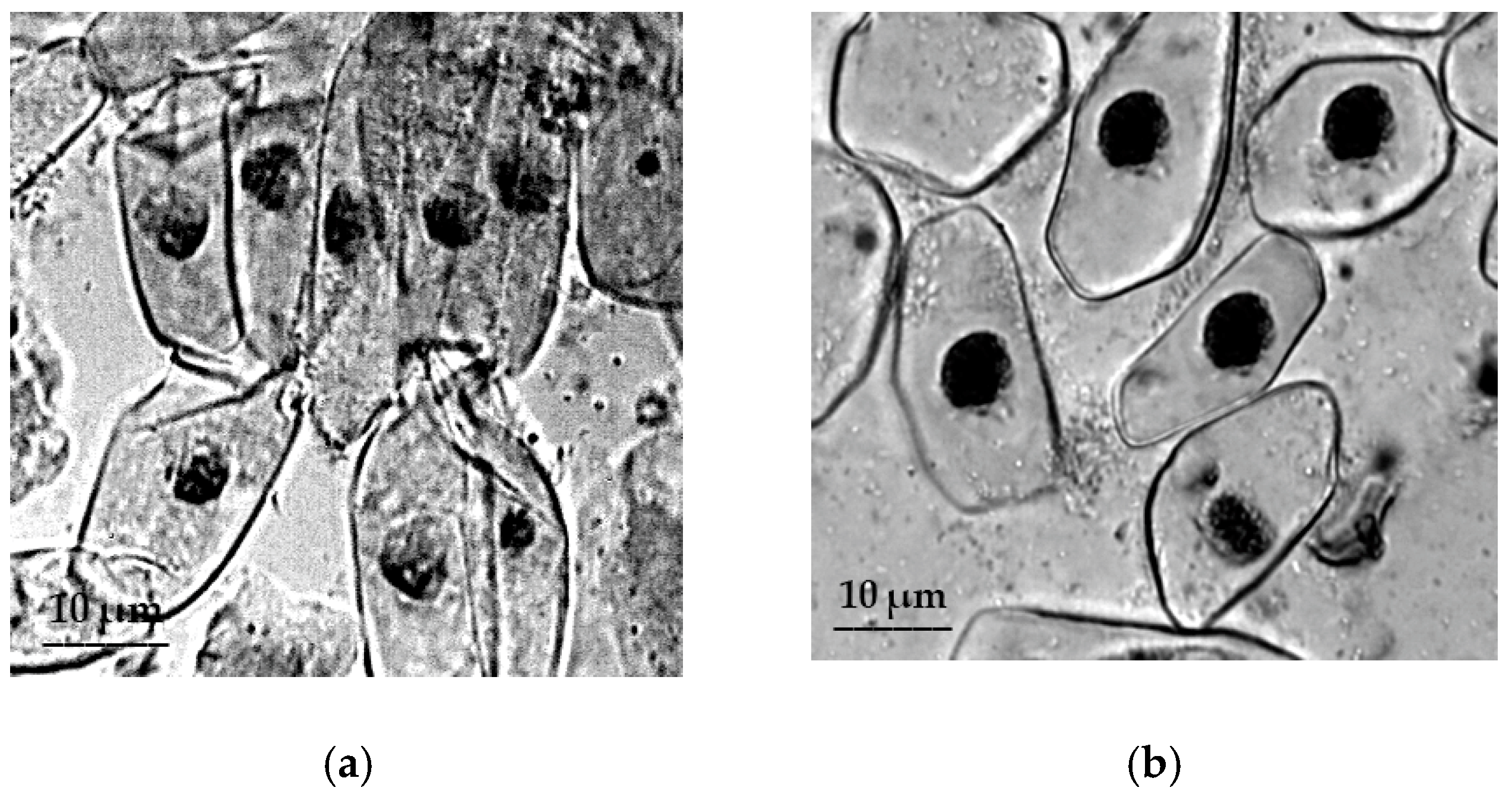
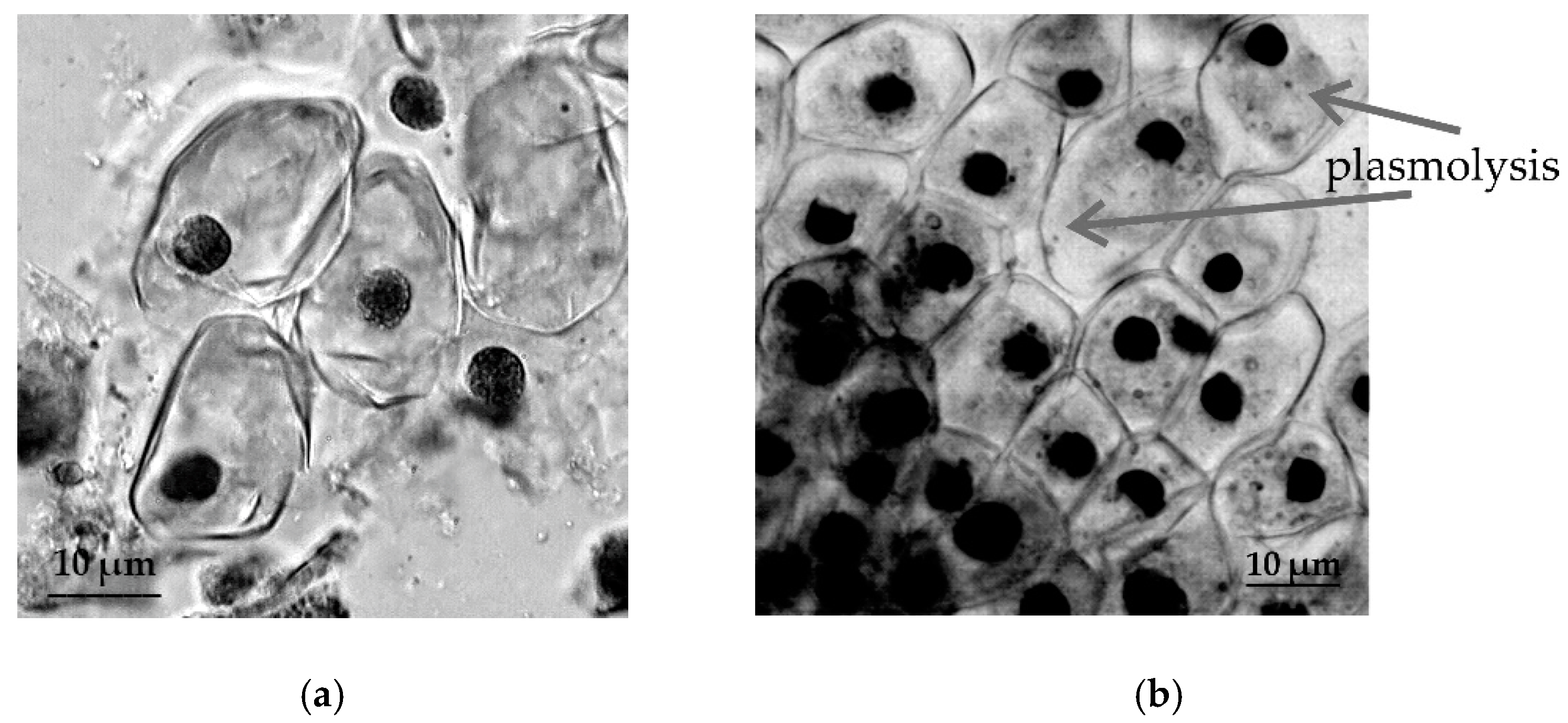
2.3. Cytological Reactions of Primary Roots of Different Types of Wheat to Osmotic Stress
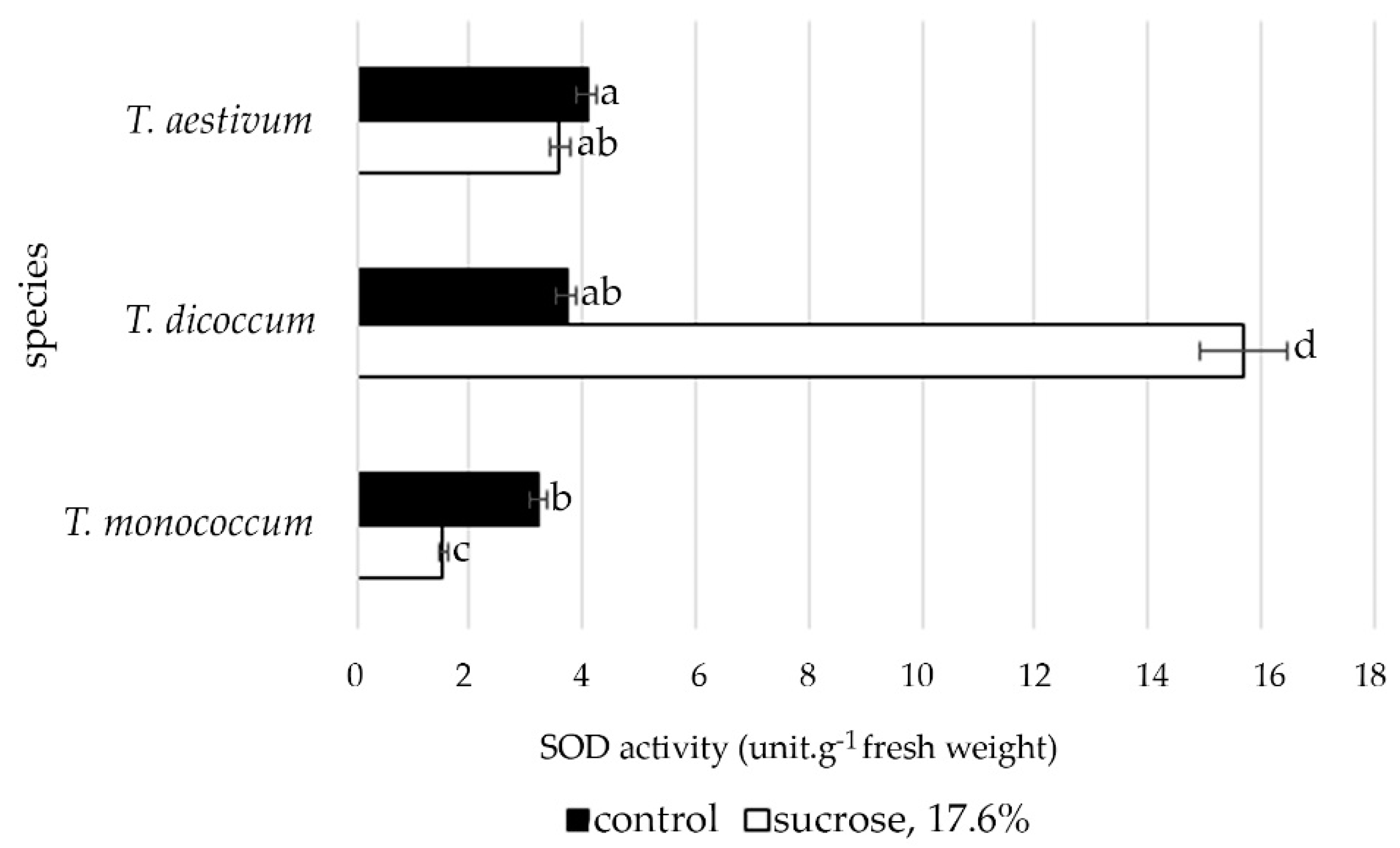
2.4. Changes in Superoxide Dismutase Activity in Primary Roots of Different Wheat Species during Osmotic Stress
3. Discussion
4. Materials and Methods
4.1. Analysis of Physiological Parameters of Drought Tolerance
4.2. Anatomical Analysis of Primary Roots
4.3. Cytological Analysis of Primary Roots
4.4. Analysis of Superoxide Dismutase (SOD) Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate trends and global crop production since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, M.; Soolanayakanahally, R.; Ogawa, S.; Uga, Y.; Selvaraj, M.G.; Kagale, S. Drought response in wheat: Key genes and regulatory mechanisms controlling root system architecture and transpiration efficiency. Front. Chem. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Atkin, O.; Macherel, D. The crucial role of plant mitochondria in orchestrating drought tolerance. Ann. Bot. 2009, 103, 581–597. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Maximova, E.; Fuggi, A.; Carillo, P. Durum Wheat Roots Adapt to Salinity Remodeling the Cellular Content of Nitrogen Metabolites and Sucrose. Front. Plant Sci. 2017, 7, 2035. [Google Scholar] [CrossRef] [PubMed]
- Bramley, H.; Turner, N.C.; Turner, D.W.; Tyerman, S.D. Roles of morphology, anatomy, and aquaporins in determining contrasting hydraulic behavior of roots. Plant Physiol. 2009, 150, 348–364. [Google Scholar] [CrossRef]
- Glyn Bengough, A.; McKenzie, B.M.; Hallett, P.D.; Valentine, T.A. Root elongation, water stress, and mechanical impedance: A review of limiting stresses and beneficial root tip traits. J. Exp. Bot. 2011, 62, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Aggarwal, P.; Robbins, N.E., II; Sturrock, C.J.; Thompson, M.C.; Tan, H.Q.; Tham, C.; Duan, L.; Rodriguez, P.L.; Vernoux, T.; et al. Plant roots use a patterning mechanism to position lateral root branches toward available water. Proc. Natl. Acad. Sci. USA 2014, 111, 9319–9324. [Google Scholar] [CrossRef]
- Lovelli, S.; Perniola, M.; Di Tommaso, T.; Bochicchio, R.; Amato, M. Specific root length and diameter of hydroponically-grown tomato plants under salinity. J. Agron. 2012, 11, 101–106. [Google Scholar] [CrossRef]
- Khan, M.A.; Gemenet, D.C.; Villordon, A. Root System Architecture and Abiotic Stress Tolerance: Current Knowledge in Root and Tuber Crops. Front. Plant Sci. 2016, 7, 1584. [Google Scholar] [CrossRef]
- Woodrow, P.; Ciarmiello, L.F.; Annunziata, M.G.; Pacifico, S.; Iannuzzi, F.; Mirto, A.; D’Amelia, L.; Dell’Aversana, E.; Piccolella, S.; Fuggi, A.; et al. Durum wheat seedling responses to simultaneous high light and salinity involve a fine reconfiguration of amino acids and carbohydrate metabolism. Physiol. Plant. 2017, 159, 290–312. [Google Scholar] [CrossRef]
- Torrey, J.G. Physiology of Root Elongation. Ann. Rev. Plant Physiol. 1956, 7, 237–266. [Google Scholar] [CrossRef]
- West, G.; Inzé, D.; Beemster, G.T.S. Cell Cycle Modulation in the Response of the Primary Root of Arabidopsis to Salt Stress. Plant Physiol. 2004, 135, 1050–1058. [Google Scholar] [CrossRef] [PubMed]
- Aroca, R.; Porcel, R.; Ruis-Lozano, J.M. Regulation of Root Water Uptake Under Drought Stress Conditions. J. Exp. Bot. 2011, 63, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Villordon, A.Q.; Firon, N.; Clark, C.A.; Smit, A. Manipulating Root System Architecture in Sweetpotato for Global Food Security: Progress Prospects and Applications. figshare. Available online: https://figshare.com/articles/Manipulating_Root_System_Architecture_in_Sweetpotato_for_Global_Food_Security_Progress_Prospects_and_Applications/1462655 (accessed on 27 March 2020).
- Serraj, R.; Sinclair, T.R. Osmolyte Accumulation: Can It Really Help Increase Crop Yield under Drought Conditions? Plant Cell Environ. 2002, 25, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P.; Wojciechowski, T. Opportunities and challenges in the subsoil: Pathways to deeper rooted crops. J. Exp. Bot. 2015, 66, 2199–2210. [Google Scholar] [CrossRef] [PubMed]
- Waines, J.W.; Endaie, B. Domestication and Crop Physiology: Roots of Green Revolution Wheat. Ann. Bot. 2007, 63, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Giehl, R.F.; Gruber, B.D.; von Wirén, N. It’s time to make changes: Modulation of root system architecture by nutrient signals. J. Exp. Bot. 2014, 65, 769–778. [Google Scholar] [CrossRef]
- Paez-Garcia, A.; Motes, C.M.; Scheible, W.; Chen, R.; Blancaflor, E.B.; Montero, M.J. Root traits and phenotyping strategies for plant improvement. Plants 2015, 4, 334–355. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, L.; Li, N.; Zhang, Q. Effect of Root Moisture Content and Diameter on Root Tensile Properties. PLoS ONE 2016, 11, e0151791. [Google Scholar] [CrossRef]
- Wasson, A.P.; Richards, R.A.; Chatrath, R.; Misra, S.C.; Prasad, S.V.; Rebetzke, G.J.; Kirkegaard, J.A.; Christopher, J.; Watt, M. Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J. Exp. Bot. 2012, 63, 3485–3498. [Google Scholar] [CrossRef]
- Nie, M.; Lu, M.; Bell, J.; Raut, S.; Pendall, E. Altered root traits due to elevated CO2: A meta-analysis. Glob. Ecol. Biogeogr. 2013, 22, 1095–1105. [Google Scholar] [CrossRef]
- Carrillo, Y.; Dijkstra, F.A.; LeCain, D.; Morgan, J.A.; Blumenthal, D.; Waldron, S.; Pendall, E. Disentangling root responses to climate change in a semiarid grassland. Oecologia 2014, 175, 699–711. [Google Scholar] [CrossRef] [PubMed]
- Suseela, V.; Tharayil, N.; Pendall, E.; Rao, A.M. Warming and elevated CO2 alter the suberin chemistry in roots of photosynthetically divergent grass species. AoB Plants 2017, 9, plx041. [Google Scholar] [CrossRef]
- Wu, J.; Guo, Y. An integrated method for quantifying root architecture of field-grown maize. Ann. Bot. 2014, 114, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Clark, L.J.; Price, A.H.; Steele, K.A.; Whalley, W.R. Evidence from near-isogenic lines that root penetration increases with root diameter and bending stiffness in rice. Func. Plant Biol. 2008, 35, 1163–1171. [Google Scholar] [CrossRef]
- Jaramillo, R.E.; Nord, E.A.; Chimungu, J.G.; Brown, K.M.; Lynch, J.P. Root cortical burden influences drought tolerance in maize. Ann. Bot. 2013, 112, 429–437. [Google Scholar] [CrossRef]
- Genet, M.; Stokes, A.; Salin, F.; Mickovski, S.B.; Fourcaud, T.; Dumail, J.F.; van Beek, R. The influence of cellulose content on tensile strength in tree roots. Plant Soil 2005, 278, 1–9. [Google Scholar] [CrossRef]
- Wu, Q.; Pagès, L.; Wu, J. Relationships between root diameter, root length and root branching along lateral roots in adult, field-grown maize. Ann. Bot. 2016, 117, 379–390. [Google Scholar] [CrossRef]
- Terletskaya, N.; Duisenbayeva, U.; Rysbekova, A.; Kurmanbayeva, M.; Blavachinskaya, I. Architectural traits in response to salinity of wheat primary roots. Acta Physiol Plant. 2019, 41, 157. [Google Scholar] [CrossRef]
- Kreszies, T.; Schreiber, L.; Ranathunge, K. Suberized transport barriers in Arabidopsis, barley and rice roots: From the model plant to crop species. J. Plant Physiol. 2018, 227, 75–83. [Google Scholar] [CrossRef]
- Enstone, D.E.; Peterson, C.A.; Ma, F. Root endodermis and exodermis: Structure, function, and responses to the environment. J. Plant Growth Regul. 2003, 21, 335–351. [Google Scholar] [CrossRef]
- Schreiber, L. Transport barriers made of cutin, suberin and associated waxes. Trends Plant Sci. 2010, 15, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Kotula, L.; Schreiber, L.; Colmer, T.D.; Nakazono, M. Anatomical and biochemical characterisation of a barrier to radial O2 loss in adventitious roots of two contrasting Hordeum marinum accessions. Func. Plant Biol. 2017, 44, 845. [Google Scholar] [CrossRef] [PubMed]
- Baxter, I.; Hosmani, P.S.; Rus, A.; Lahner, B.; Borevitz, J.O.; Muthukumar, B.; Mickelbart, M.V.; Schreiber, L.; Franke, R.B.; Salt, D.E. Root suberin forms an extracellular barrier that affects water relations and mineral nutrition in Arabidopsis. PLoS Genet. 2009, 5, e1000492. [Google Scholar] [CrossRef] [PubMed]
- Stevović, S.; Mikovilović, V.S.; Ćalić-Dragosavac, D. Environmental impact on morphological and anatomical structure of Tansy. AJB 2010, 9, 2413–2421. [Google Scholar] [CrossRef]
- Ceccoli, G.; Ramos, J.C.; Ortega, L.I.; Acosta, J.M.; Perreta, M.G. Salinity induced anatomical and morphological changes in Chloris gayana Kunth roots. Biocell 2011, 35, 9–17. [Google Scholar] [CrossRef]
- De Bauw, P.; Vandamme, E.; Lupembe, A.; Mwakasege, L.; Senthilkumar, K.; Dramé, K.N.; Merckx, R. Anatomical root responses of rice to combined phosphorus and water stress—Relations to tolerance and breeding opportunities. Func. Plant Biol. 2019, 46, 1009–1022. [Google Scholar] [CrossRef]
- Ouyang, W.; Yin, X.; Yang, J.; Struik, P.C. Comparisons with wheat reveal root anatomical and histochemical constraints of rice under water-deficit stress. Plant Soil 2020. [Google Scholar] [CrossRef]
- Zwieniecki, M.A.; Secchi, F. Threats to xylem hydraulic function of trees under ‘new climate normal’ conditions. Plant Cell Environ. 2015, 38, 1713–1724. [Google Scholar] [CrossRef]
- Steudle, E. Water uptake by roots: Effects of water deficit. J. Exp. Bot. 2000, 51, 1531–1542. [Google Scholar] [CrossRef]
- Zhu, Y.H.; Weiner, J.; Yu, M.X.; Li, F.M. Evolutionary Agroecology: Trends in root architecture during wheat breeding. Evol. Appl. 2019, 12, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Malokostova, E.I. Main Directions of Spring Wheat Breeding for Drought Resistance. Zemledelie 2018, 3, 37–39. (In Russian) [Google Scholar] [CrossRef]
- Prosyannikova, E.B.; Rodchenko, O.P. Histogenesis of Root Conducting System. Sci. Notes ZabGSPU (Nat. Sci.) 2009, 1, 91–94. [Google Scholar]
- Shimazaki, Y.; Ookawa, T.; Hirasawa, T. The root tip and accelerating region suppress elongation of the decelerating region without any effects on cell turgor in primary roots of maize under water stress. Plant Physiol. 2005, 139, 458–465. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Carden, D.E.; Walker, D.J.; Flowers, T.J.; Miller, A.J. Signal cell measurements of the contributions of cytosolic Na+ and K+ to salt tolerance. Plant Physiol. 2003, 131, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Adem, G.; Roy, S.J.; Zhou, M.; Bowman, J.P.; Shabala, S. Evaluating contribution of ionic, osmotic and oxidative stress components towards salinity tolerance in barley. BMC Plant Biol. 2014, 14, 113. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zhang, W.; Li, B.; Wang, Y.; Li, K.; Sodmergen; Han, C.; Zhang, Y.; Li, X. An endoplasmic reticulum response pathway mediates programmed cell death of root tip induced by water stress in Arabidopsis. New Phytol. 2010, 186, 681–695. [Google Scholar] [CrossRef] [PubMed]
- Gahoonia, T.S.; Nielsen, N.E.; Joshi, P.A.; Jahoor, A.A. Root hairless barley mutant for elucidating genetic of root hairs and phosphorus uptake. Plant Soil 2001, 235, 211–219. [Google Scholar] [CrossRef]
- Tanaka, N.; Kato, M.; Tomioka, R.; Kurata, R.; Fukao, Y.; Aoyama, T.; Maeshima, M. Characteristics of a root hair-less line of Arabidopsis thaliana under physiological stresses. J. Exp. Bot. 2014, 65, 1497–1512. [Google Scholar] [CrossRef]
- Carol, R.J.; Dolan, L. The role of reactive oxygen species in cell growth: Lessons from root hairs. J. Exp. Bot. 2006, 57, 1829–1834. [Google Scholar] [CrossRef]
- Lee, S.H.; Singh, A.P.; Chung, G.C. Rapid accumulation of hydrogen peroxide in cucumber roots due to exposure to low temperature appears to mediate decreases in water transport. J. Exp. Bot. 2004, 55, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Dubey, R.S. Involvement of oxidative stress and role of antioxidative defense system in growing rice seedlings exposed to toxic concentrations of aluminum. Plant Cell Rep. 2007, 26, 2027–2038. [Google Scholar] [CrossRef] [PubMed]
- Tyburski, J.B.; Dunajska, K.; Tretyn, A. Reactive oxygen species localization in roots of Arabidopsis thaliana seedlings grown under phosphate deficiency. Plant Grow. Reg. 2009, 59, 27–36. [Google Scholar] [CrossRef]
- Garg, N.; Manchanda, G. ROS generation in plants: Boon or bane? Plant Biosyst. 2009, 143, 81–96. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.; et al. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 2003, 422, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.S.; Van Breusegem, F.; Stone, J.M.; Denev, I.; Laloi, C. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. Bioessays 2006, 28, 1091–1101. [Google Scholar] [CrossRef]
- Libik-Konieczny, M.; Kozieradzka-Kiszkurno, M.; Michalec-Warzecha, Ż.; Miszalski, Z.; Bizan, J.; Konieczny, R. Influence of anti- and prooxidants on rhizogenesis from hypocotyls of Mesembryanthemum crystallinum L. cultured in vitro. Acta Physiol. Plant. 2017, 39, 166. [Google Scholar] [CrossRef]
- Abozeid, A.; Ying, Z.; Lin, Y.; Liu, J.; Zhang, Z.; Tang, Z. Ethylene Improves Root System Development under Cadmium Stressby Modulating Superoxide AnionConcentration in Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 253. [Google Scholar] [CrossRef]
- Sánchez-Calderón, L.; Ibarra-Cortés, M.E.; Zepeda-Jazo, I. Root Development and Abiotic Stress Adaptation; Vahdati, K., Leslie, C., Eds.; InTech: London, UK, 2013; pp. 135–168. [Google Scholar] [CrossRef]
- Demidchik, V.; Shabala, S.N.; Coutts, K.B.; Tester, M.A.; Davies, J.M. Free oxygen radicals regulate plasma membrane Ca2+- and K+-permeable channels in plant root cells. J Cell Sci. 2003, 116, 81–88. [Google Scholar] [CrossRef]
- Passardi, F.; Tognolli, M.; De Meyer, M.; Penel, C.; Dunand, C. Two cell wall associated peroxidases from Arabidopsis influence root elongation. Planta 2006, 223, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Terletskaya, N.V.; Shcherban, A.B.; Nesterov, M.A.; Perfil’ev, R.N.; Salina, E.A.; Altayeva, N.A.; Blavachinskaya, I.V. Drought Stress Tolerance and Photosynthetic Activity of Alloplasmic Lines, T. dicoccum x T. aestivum. Int. J. Mol. Sci. 2020, 21, 3356. [Google Scholar] [CrossRef] [PubMed]
- Methodical guidance. In Diagnostics Stability of Plant Resistance to Stress; Udovenko, G.V., Ed.; VIR: Leningrad, Russia, 1988; 89p. [Google Scholar]
- Barykina, R.P. Guide on Botanical Microtechique; Base and Methods; MSU: Moscow, Russia, 2004; 312p. [Google Scholar]
- Pausheva, Z.P. Workshop on cytology (Praktikum po tsitologii); Kolos: Moskow, Russia, 1970. [Google Scholar]
- Van Rossun, M.W.P.C.; Alberda, M.; Van Der Plas, L.H.W. Role of oxidative damage in tulip bulb scale micropropagation. Plant Sci. 1997, 130, 207–216. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, J. Superoxide Dismutase: Improved Assays and an Assay Applicable to Acrylamide Gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condi-tions | Parameters | |||||
|---|---|---|---|---|---|---|
| Thickness of the Epiblem, μm | Thickness of the Exoderm, μm | Thickness of the Cortical Parenchyma, μm | Thickness of the Endoderm, μm | Diameter of the Central Cylinder (Stele), μm | Diameter of the Root, μm | |
| T. monococcum | ||||||
| control | 14.8 ± 0.7 | 13.9 ± 0.4 | 74.9 ± 1.7 | 18.6 ± 0.6 | 191.4 ± 2.2 | 435.8 ± 7.1 |
| sucrose, 17.6% | 14.1 ± 0.7 | 27.7 ± 0.2 * | 85.1 ± 1.9 * | 18.6 ± 0.5 | 187.6 ± 1.4 | 478.5 ± 2.2 * |
| % to control | 95 | 199 | 113 | 100 | 98 | 110 |
| T. dicoccum | ||||||
| control | 19.4 ± 0.2 | 16.7 ± 0.8 | 67.3 ± 0.4 | 12.8 ± 0.6 | 217.5 ± 3.5 | 449.9 ± 5.2 |
| sucrose, 17.6% | 13.7 ± 0.1 * | 30.4 ± 0.6 * | 80.0 ± 0.7 * | 26.4 ± 0.8 * | 192.9 ± 1.1 * | 493.9 ± 0.6 * |
| % to control | 71 | 182 | 119 | 206 | 89 | 110 |
| T. aestivum | ||||||
| control | 16.6 ± 0.8 | 28.5 ± 0.9 | 70.1 ± 1.2 | 19.7 ± 0.5 | 205.8 ± 3.6 | 475.5 ± 9.2 |
| sucrose, 17.6% | 18.5 ± 0.1 * | 25.6 ± 0.8 * | 75.9 ± 0.5 * | 24.5 ± 0.3 * | 234.0 ± 8.9 * | 523.0 ± 9.8 * |
| % to control | 112 | 90 | 108 | 124 | 114 | 110 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terletskaya, N.V.; Lee, T.E.; Altayeva, N.A.; Kudrina, N.O.; Blavachinskaya, I.V.; Erezhetova, U. Some Mechanisms Modulating the Root Growth of Various Wheat Species under Osmotic-Stress Conditions. Plants 2020, 9, 1545. https://doi.org/10.3390/plants9111545
Terletskaya NV, Lee TE, Altayeva NA, Kudrina NO, Blavachinskaya IV, Erezhetova U. Some Mechanisms Modulating the Root Growth of Various Wheat Species under Osmotic-Stress Conditions. Plants. 2020; 9(11):1545. https://doi.org/10.3390/plants9111545
Chicago/Turabian StyleTerletskaya, Nina V., Tamara E. Lee, Nazira A. Altayeva, Nataliya O. Kudrina, Irina V. Blavachinskaya, and Ulzhan Erezhetova. 2020. "Some Mechanisms Modulating the Root Growth of Various Wheat Species under Osmotic-Stress Conditions" Plants 9, no. 11: 1545. https://doi.org/10.3390/plants9111545
APA StyleTerletskaya, N. V., Lee, T. E., Altayeva, N. A., Kudrina, N. O., Blavachinskaya, I. V., & Erezhetova, U. (2020). Some Mechanisms Modulating the Root Growth of Various Wheat Species under Osmotic-Stress Conditions. Plants, 9(11), 1545. https://doi.org/10.3390/plants9111545