Inhibition of NO Biosynthetic Activities during Rehydration of Ramalina farinacea Lichen Thalli Provokes Increases in Lipid Peroxidation

 , , ,
, , ,  and
and
Abstract
1. Introduction
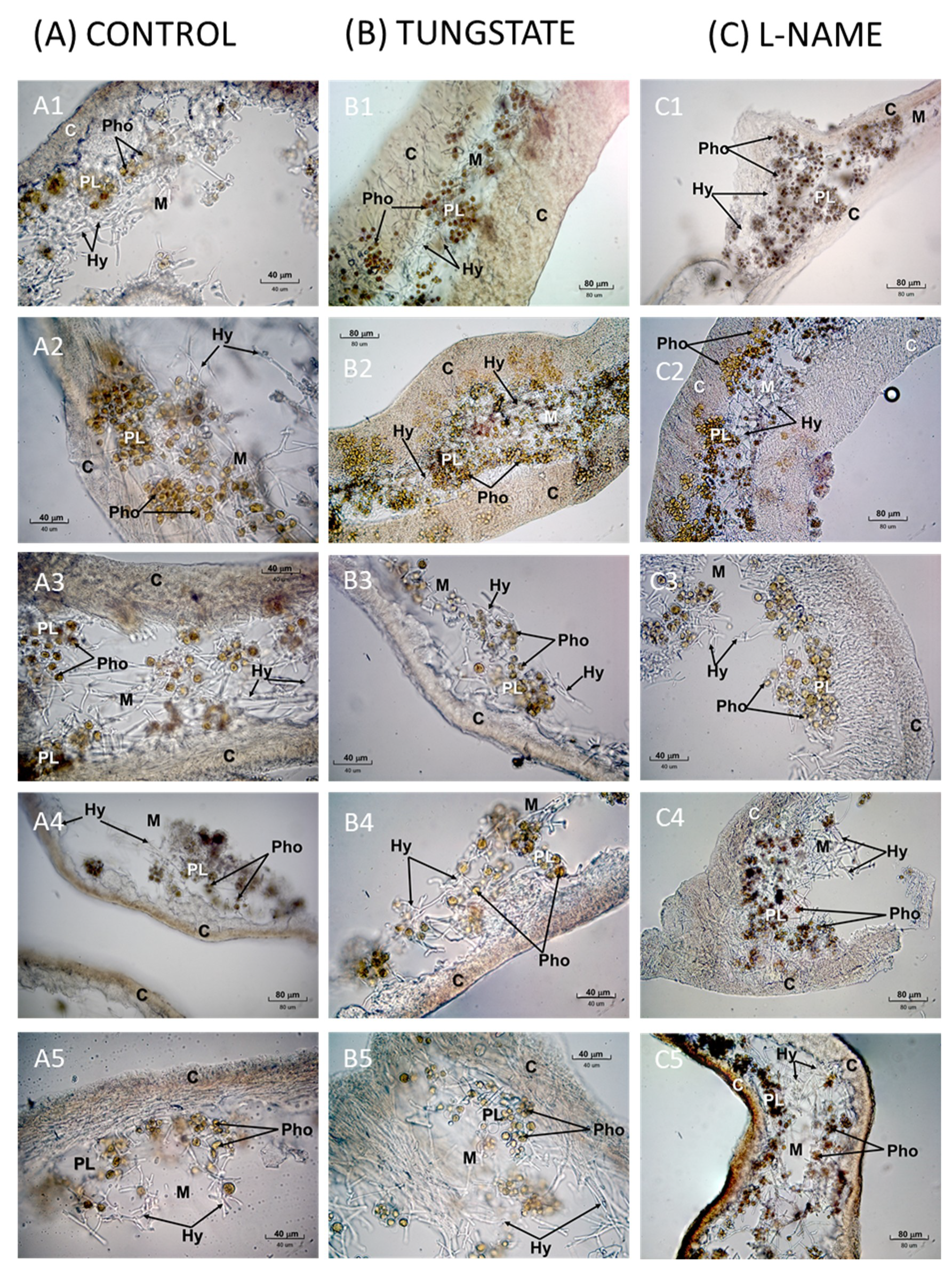
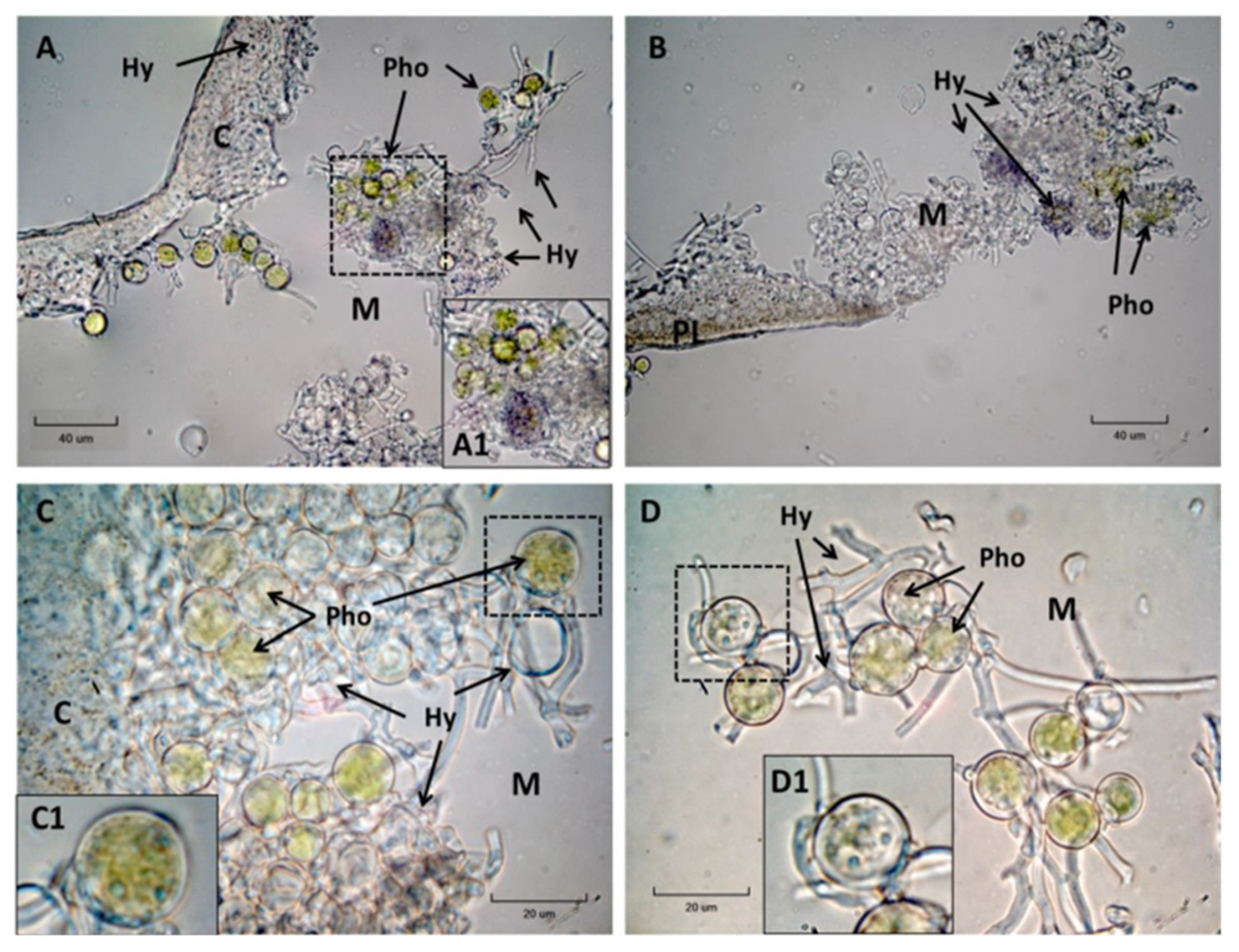
2. Results
2.1. Effects of NR Inhibition on Lipid Peroxidation during Lichen Rehydration
2.2. Effects of Nitric Oxide Synthase (NOS) Inhibition on Lipid Peroxidation during Lichen Rehydration
2.3. NO Endproducts
2.4. Diaphorase Activity
2.5. Specific Activity of NR
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Lichen Material
4.3. Treatments
4.4. Measurement of Lipid Peroxidation: MDA
4.5. NO Endproducts Determination
4.6. Diaphorase Activity
4.7. Specific Activity of NR
4.8. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BHT | 2,6-Di-tert-buthyl-4-methylphenol |
| BSA | Bovine serum albumin |
| DTT | Dithiothreitol |
| DW | Dry weight |
| etOH | Ethanol |
| EDTA | Ethylenediaminetetraacetic Acid |
| L-NAME | NG-nitro-L-arginine methyl ester |
| MDA | Malondialdehyde |
| NBT | Nitro blue tetrazolium |
| NOx | Nitric oxide oxidation end-products |
| NOS | Nitric oxide synthase |
| NR | Nitrate reductase |
| PBS | Phosphate buffered saline |
| ROS | Reactive oxygen species |
| TBA | 2-Thiobarbituric acid |
| TBARS | Thiobarbituric Acid Reactive Substances |
| TCA | Trichloroacetic acid |
| TEP | 1,1,3,3- Tetraexthoxypropane |
References
- Ahmadjian, V. The Lichen Symbiosis; John Wiley & Sons: New York, NY, USA, 1993; ISBN 0471578851. [Google Scholar]
- Izco, J.; Barreno, E.; Brugués, M.; Costa, M.; Devesa, J.A.; Fernández-González, F.; Llimona, X.; Valdés, B. Botánica; McGraw Hill Interamericana: Madrid, Spain, 1997; ISBN 8448601823. [Google Scholar]
- Kranner, I.; Beckett, R.; Hochman, A.; Nash, T.H. Desiccation-tolerance in lichens: A review. Bryologist 2008, 111, 576–593. [Google Scholar] [CrossRef]
- Kranner, I.; Cram, W.J.; Zorn, M.; Wornik, S.; Yoshimura, I.; Stabentheiner, E.; Pfeifhofer, H.W. Antioxidants and photoprotection in a lichen as compared with its isolated symbiotic partners. Proc. Natl. Acad. Sci. USA 2005, 102, 3141–3146. [Google Scholar] [CrossRef] [PubMed]
- McKersie, B.D.; Leshem, Y.Y. Stress and Stress Coping in Cultivated Plants; Kluwer Academic Publishers: Norwell, MA, USA, 1994. [Google Scholar]
- Halliwell, B.; Cross, C.E. Oxygen-derived species: Their relation to human disease and environmental stress. Environ. Health Perspect. 1994, 102, 5–12. [Google Scholar] [PubMed]
- Wilson, I.D.; Neill, S.J.; Hancock, J.T. Nitric oxide synthesis and signalling in plants. Plant. Cell Environ. 2008, 31, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Meilhoc, E.; Cam, Y.; Skapski, A.; Bruand, C. The Response to Nitric Oxide of the Nitrogen-Fixing Symbiont Sinorhizobium meliloti. Mol. Plant Microbe Interact. 2010, 748, 748–759. [Google Scholar] [CrossRef] [PubMed]
- Feelisch, M.; Martin, J.F. The early role of nitric oxide in evolution. Trends Ecol. Evol. 1995, 10, 496–499. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Vranová, E.; Inzé, D.; Van Breusegem, F. Signal transduction during oxidative stress. J. Exp. Botany 2002, 53, 1227–1236. [Google Scholar] [CrossRef]
- Millar, A.; Day, D. Nitric oxide inhibits the cytochrome oxidase but not the alternative oxidase of plant mitochondria. FEBS Lett. 1996, 398, 155–158. [Google Scholar] [CrossRef]
- Caro, A.; Puntarulo, S. Nitric oxide decreases superoxide anion generation by microsomes from soybean embryonic axes. Physiol. Plant. 1998, 104, 357–364. [Google Scholar] [CrossRef]
- Boveris, A.D.; Galatro, A.; Puntarulo, S. Effect of nitric oxide and plant antioxidants on microsomal content of lipid radicals. Biol. Res. 2000, 33, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Wendehenne, D.; Hancock, J.T. New frontiers in nitric oxide biology in plant Preface. Plant Sci. 2011, 181, 507–508. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Mori, M.; Pichtel, J.; Ahmad, A. Nitric Oxide in Plant Physiology; Whiley-Blackwell: Hoboken, NJ, USA, 2009. [Google Scholar]
- Gupta, K.J.; Fernie, A.R.; Kaiser, W.M.; van Dongen, J.T. On the origins of nitric oxide. Trends Plant Sci. 2011, 16, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Lobysheva, I.; Vanin, A.; Sineshchekov, O.; Govorunova, E. Phototaxis in Chlamydomonas reinhardtii is modulated by nitric oxide. Biofizika 1996, 41, 540–541. [Google Scholar]
- Mallick, N.; Mohn, F.H.; Soeder, C.J.; Grobbelaar, J.U. Ameliorative role of nitric oxide on H2O2 toxicity to a chlorophycean alga Scenedesmus obliquus. J. Gen. Appl. Microbiol. 2002, 48, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Feng, H.; Zhang, M.; Wang, X. Nitric oxide alleviates oxidative damage in the green alga Chlorella pyrenoidosa caused by UV-B radiation. Folia Microbiol. (Praha) 2003, 48, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Wilken, M.; Huchzermeyer, B. Suppression of mycelia formation by NO produced endogenously in Candida tropicalis. Eur. J. Cell Biol. 1999, 78, 209–213. [Google Scholar] [CrossRef]
- Maier, J.; Hecker, R.; Rockel, P.; Ninnemann, H. Role of nitric oxide synthase in the light-induced development of sporangiophores in Phycomyces blakesleeanus. Plant Physiol. 2001, 126, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Huang, C.; Chen, Q.; Zou, Y.; Zhang, J. Nitric oxide alleviates heat stress-induced oxidative damage in Pleurotus eryngii var. tuoliensis. Fungal Genet. Biol. 2012, 49, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Jeong, C.-S.; Choi, H.-S. Identification of Nitric Oxide Synthase in Flammulina velutipes. Mycologia 2000, 92, 1027–1032. [Google Scholar] [CrossRef]
- Catalá, M.; Gasulla, F.; Pradas del Real, A.E.; García-Breijo, F.; Reig-Armiñana, J.; Barreno, E. Fungal-associated NO is involved in the regulation of oxidative stress during rehydration in lichen symbiosis. BMC Microbiol. 2010, 10, 297. [Google Scholar] [CrossRef] [PubMed]
- Weissman, L.; Garty, J.; Hochman, A. Rehydration of the lichen Ramalina lacera results in production of reactive oxygen species and nitric oxide and a decrease in antioxidants. Appl. Environ. Microbiol. 2005, 71, 2121–2129. [Google Scholar] [CrossRef] [PubMed]
- Catalá, M.; Gasulla, F.; Pradas Del Real, A.E.; García-Breijo, F.; Reig-Armiñana, J.; Barreno, E. The organic air pollutant cumene hydroperoxide interferes with NO antioxidant role in rehydrating lichen. Environ. Pollut. 2013, 179, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Wendehenne, D.; Pugin, A.; Klessig, D.F.; Durner, J. Nitric oxide: Comparative synthesis and signaling in animal and plant cells. Trends Plant Sci. 2001, 6, 177–183. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide and the regulation of gene expression. Trends Cell Biol. 2001, 11, 66–75. [Google Scholar] [CrossRef]
- Yamasaki, H.; Sakihama, Y.; Takahashi, S. An alternative pathway for nitric oxide production in plants: New features of an old enzyme. Trends Plant Sci. 1999, 4, 128–129. [Google Scholar] [CrossRef]
- Berges, J. Miniview: Algal nitrate reductases. Eur. J. Phycol. 1997, 32, 3–8. [Google Scholar] [CrossRef]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, Á.; Ocaña-Calahorro, F.; Mariscal, V.; Carreras, A.; Barroso, J.B.; Galván, A.; Fernández, E. A dual system formed by the ARC and NR molybdoenzymes mediates nitrite-dependent NO production in Chlamydomonas. Plant Cell Environ. 2016, 39, 2097–2107. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B. Nitric oxide synthase-like activity in higher plants. Nitric Oxide Biol. Chem. 2017, 68, 5–6. [Google Scholar] [CrossRef]
- Foresi, N.; Correa-Aragunde, N.; Parisi, G.; Caló, G.; Salerno, G.; Lamattina, L. Characterization of a nitric oxide synthase from the plant kingdom: NO generation from the green alga Ostreococcus tauri is light irradiance and growth phase dependent. Plant Cell 2010, 22, 3816–3830. [Google Scholar] [CrossRef]
- Hichri, I.; Boscari, A.; Meilhoc, E.; Catalá, M.; Barreno, E.; Bruand, C.; Lanfranco, L.; Brouquisse, R. Nitric Oxide: A multitask Player in Plant–Microorganism Symbioses. In Gasotransmitters in Plants. The Rise of a New Paradigm in Cell Signaling; Lamattina, L., García-Mata, C., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 239–268. [Google Scholar]
- Moya, P.; Molins, A.; Martinez-Alberola, F.; Muggia, L.; Barreno, E. Unexpected associated microalgal diversity in the lichen Ramalina farinacea is uncovered by pyrosequencing analyses. PLoS ONE 2017, 12, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Catalá, M.; Gasulla, F.; Pradas del Real, A.E.; García-Breijo, F.J.; Reig-Armiñana, J.; Barreno, E. Nitric oxide is involved in oxidative stress during rehydration of Ramalina farinacea (L.) Ach. in the presence of the oxidative air pollutant cumene hydroperoxide. Biol. Lichens Symbiosis Ecol. Environ. Monit. Syst. Cyber Appl. 2010, 105, 87–92. [Google Scholar]
- Cueto, M.; Hernández-Perera, O.; Martín, R.; Bentura, M.L.; Rodrigo, J.; Lamas, S.; Golvano, M.P. Presence of nitric oxide synthase activity in roots and nodules of Lupinus albus. FEBS Lett. 1996, 398, 159–164. [Google Scholar] [CrossRef]
- Chow, F.; Capociama, F.V.; Faria, R.; de Oliveira, M.C. Characterization of nitrate reductase activity in vitro in Gracilaria caudata J. Agardh (Rhodophyta, Gracilariales). Rev. Bras. Bot. 2007, 30, 123–129. [Google Scholar] [CrossRef]
- Groß, F.; Durner, J.; Gaupels, F. Nitric oxide, antioxidants and prooxidants in plant defence responses. Front. Plant Sci. 2013, 4, 419. [Google Scholar] [CrossRef] [PubMed]
- Darley-Usmar, V.M.; Pate, R.P.; O’Donnell, V.B.; Freeman, B.A. Antioxidant Actions of Nitric Oxide. In Nitric Oxide: Biology and Pathology; Ignarro, L., Ed.; Academic Press: Los Ángeles, CA, USA, 2000; pp. 265–276. ISBN 978-0-12-370420-7. [Google Scholar]
- Xiong, J.; Fu, G.; Yang, Y.; Zhu, C.; Tao, L. Tungstate: Is it really a specific nitrate reductase inhibitor in plant nitric oxide research? J. Exp. Bot. 2012, 63, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Sakihama, Y.; Nakamura, S.; Yamasaki, H. Nitric oxide production mediated by nitrate reductase in the green alga Chlamydomonas reinhardtii: An alternative NO production pathway in photosynthetic organisms. Plant Cell Physiol. 2002, 43, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Mallick, N.; Rai, L.C.; Mohn, F.H.; Soeder, C.J. Studies on nitric oxide (NO) formation by the green alga Scenedesmus obliquus and the diazotrophic cyanobacterium Anabaena doliolum. Chemosphere 1999, 39, 1601–1610. [Google Scholar] [CrossRef]
- Medina-Andres, R.; Solano-Peralta, A.; Saucedo-Vazquez, J.P.; Napsucialy-Mendivil, S.; Pimentel-Cabrera, J.A.; Sosa-Torres, M.E.; Dubrovsky, J.G.; Lira-Ruan, V. The nitric oxide production in the moss Physcomitrella patens is mediated by nitrate reductase. PLoS ONE 2015, 10, e0119400. [Google Scholar] [CrossRef] [PubMed]
- Cánovas, D.; Marcos, J.F.; Marcos, A.T.; Strauss, J. Nitric oxide in fungi: Is there NO light at the end of the tunnel? Curr. Genet. 2016, 62, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Slot, J.C.; Hibbett, D.S. Horizontal transfer of a nitrate assimilation gene cluster and ecological transitions in fungi: A phylogenetic study. PLoS ONE 2007, 2, e1097. [Google Scholar] [CrossRef] [PubMed]
- Kopincová, J.; Púzserová, A.; Bernátová, I. L-NAME in the cardiovascular system – nitric oxide synthase activator? Pharmacol. Rep. 2012, 64, 511–520. [Google Scholar] [CrossRef]
- Gross, B.H.; Kreutz, K.J.; Osterberg, E.C.; McConnell, J.R.; Handley, M.; Wake, C.P.; Yalcin, K. Constraining recent lead pollution sources in the North Pacific using ice core stable lead isotopes. J. Geophys. Res. Atmos. 2012, 117, D16307. [Google Scholar] [CrossRef]
- Kim, D.; Yamaguchi, K.; Oda, T. Nitric oxide synthase-like enzyme mediated nitric oxide generation by harmful red tide phytoplankton, Chattonella marina. J. Plankton Res. 2006, 28, 613–620. [Google Scholar] [CrossRef]
- Valentovicová, K.; Halusková, L.; Huttová, J.; Mistrík, I.; Tamás, L. Effect of cadmium on diaphorase activity and nitric oxide production in barley root tips. J. Plant Physiol. 2010, 167, 10–14. [Google Scholar] [CrossRef]
- Roszer, T. The Biology of Subcellular Nitric Oxide; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2012; ISBN 9789400728189. [Google Scholar]
- Thomas, T.E.; Harrison, P.J. A Comparison of Invitro and Invivo Nitrate Reductase Assays in 3 Intertidal Seaweeds. Bot. Mar. 1988, 31, 101–107. [Google Scholar] [CrossRef]
- Granbom, M.; Chow, F.; Lopes, P.F.; De Oliveira, M.C.; Colepicolo, P.; De Paula, E.J.; Pedersén, M. Characterisation of nitrate reductase in the marine macroalga Kappaphycus alvarezii (Rhodophyta). Aquat. Bot. 2004, 78, 295–305. [Google Scholar] [CrossRef]
- Lopes, P.F.; Oliveira, M.C.; Colepicolo, P. Diurnal Fluctuation of Nitrate Reductase Activity in the Marine Red Alga Gracilaria Tenuistipitata (Rhodophyta). J. Phycol. 1997, 33, 225–231. [Google Scholar] [CrossRef]
- Chow, F.; De Oliveira, M.C.; Pedersén, M. In vitro assay and light regulation of nitrate reductase in red alga Gracilaria chilensis. J. Plant Physiol. 2004, 161, 769–776. [Google Scholar] [CrossRef]
- Zhao, M.-G.; Chen, L.; Zhang, L.-L.; Zhang, W.-H. Nitric Reductase-Dependent Nitric Oxide Production Is Involved in Cold Acclimation and Freezing Tolerance in Arabidopsis. Plant Physiol. 2009, 151, 755–767. [Google Scholar] [CrossRef]
- Hwang, S.; Williams, S.; Brinhuis, B. Changes in internal dissolved nitrogen pools as related to nitrate uptake and assimilaitonin Gracilaria tikvahiae McLachlan (Rhodophyta). Bot. Mar. 1987, 30, 11–19. [Google Scholar] [CrossRef]
- Berges, J.A.; Harrison, P.J. Nitrate reductase activity quantitatively predicts the rate of nitrate incorporation under steady state light limitation. Limnol. Oceanogr. 1995, 40, 82–93. [Google Scholar] [CrossRef]
- Botsoglou, N.A.; Fletouris, D.J.; Papageorgiou, G.E.; Vassilopoulos, V.N.; Mantis, A.J.; Trakatellis, A.G. Rapid, Sensitive, and Specific Thiobarbituric Acid Method for Measuring Lipid-Peroxidation in Animal Tissue, Food, and Feedstuff Samples. J. Agric. Food Chem. 1994, 42, 1931–1937. [Google Scholar] [CrossRef]
- Du, Z.; Bramlage, W.J. Modified thiobarbituric acid assay for measuring lipid oxidation in sugar-rich plant tissue extracts. J. Agric. Food Chem. 1992, 40, 1566–1570. [Google Scholar] [CrossRef]
- Reilly, A.; Aust, S.D. Measurement of lipid peroxidation. Free Radic. Res. 1999, 28, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Fukuto, J.M.; Cho, J.Y.; Switzer, C.H. The Chemical Properties of Nitric Oxide and Related Nitrogen Oxides. In Nitric Oxide. Biology and Pathology; Ignarro, L., Ed.; Academic Press: Los Ángeles, CA, USA, 2000; pp. 23–40. ISBN 978-0-12-370420-7. [Google Scholar]
- Nagano, T. Practical methods for detection of nitric oxide, Luminescence. J. Biol. Chem. Lumin. 1999, 14, 283–290. [Google Scholar]
- Nussler, A.K.; Glanemann, M.; Schirmeier, A.; Liu, L.; Nussler, N.C. Fluorometric measurement of nitrite/nitrate by 2,3-diaminonaphthalene. Nat. Protoc. 2006, 1, 2223–2226. [Google Scholar] [CrossRef] [PubMed]
- Hope, B.T.; Vincent, S.R. Histochemical characterization of neuronal NADPH-diaphorase. J. Histochem. Cytochem. 1989, 37, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Hope, B.T.; Michael, G.J.; Knigge, K.M.; Vincent, S.R. Neuronal NADPH diaphorase is a nitric oxide synthase. Proc. Natl. Acad. Sci. USA 1991, 88, 2811–2814. [Google Scholar] [PubMed]
- Griess, P. Bemerkungen zu der Abhandlung der HH. Weselsky und Benedikt “Ueber einige Azoverbindungen.”. Berichte Dtsch. Chem. Ges. 1879, 12, 426–428. [Google Scholar] [CrossRef]
- Chaki, M.; Valderrama, R.; Fernández-Ocaña, A.M.; Carreras, A.; Gómez-Rodríguez, M.V.; Pedrajas, J.R.; Begara-Morales, J.C.; Sánchez-Calvo, B.; Luque, F.; Leterrier, M.; et al. Mechanical wounding induces a nitrosative stress by down-regulation of GSNO reductase and an increase in S-nitrosothiols in sunflower (Helianthus annuus) seedlings. J. Exp. Bot. 2011, 62, 1803–1813. [Google Scholar] [CrossRef] [PubMed]
- Noble, J.E.; Bailey, M.J.A. Quantitation of Protein. In Methods in Enzymology; Elsevier Inc.: Amsterdam, The Netherlands, 2009; Volume 463, pp. 73–95. ISBN 9780123745361. [Google Scholar]



{kind=link}
{kind=link}
| Time of Rehydration (min) | Lipid Peroxidation Level (nEq MDA/g Dry Weight) | p Value (Student’s t-Test) | |
|---|---|---|---|
| Control | 100 µM Tungstate | ||
| 5 | 81.47 ± 8.14 | 83.98 ± 6.28 | 0.809 |
| 30 | 102.21 ± 12.43 | 115.16 ± 7.42 | 0.381 |
| 60 | 113.70 ± 13.73 | 144.82 ± 18.42 | 0.189 |
| 120 | 131.41 ± 18.80 | 191.88 ± 11.06 | 0.011 * |
| 240 | 87.69 ± 7.61 | 108.60 ± 7.36 | 0.061 |
| Time of Rehydration (min) | Lipid Peroxidation Level (nEq MDA/g Dry Weight) | p Value (Student’s t-Test) | |
|---|---|---|---|
| Control | 300 µM L-NAME | ||
| 5 | 81.47 ± 8.14 | 110.86 ± 14.90 | 0.09741 |
| 30 | 110.51 ± 12.17 | 137.51 ± 11.77 | 0.12500 |
| 60 | 104.32 ± 11.76 | 121.60 ± 10.29 | 0.28040 |
| 120 | 72.77 ± 5.46 | 89.45 ± 4.69 | 0.03021 * |
| 240 | 44.74 ± 4.66 | 74.56 ± 6.29 | 0.00096 * |
| Time of Rehydration (min) | NO Endproducts Levels (% Relative to Controls) | |||
|---|---|---|---|---|
| 100 µM Tungstate | p Value | 300 µM L-NAME | p Value | |
| 5 | 72.70 % ± 19.23 % | 0.3496 | 88.77 % ± 16.45 % | 0.4554 |
| 30 | 28.11 % ± 4.20 % | 0.0018 * | 188.56 % ± 24.96 % | 0.0038 * |
| 60 | 67.22 % ± 14.34 % | 0.2946 | 128.40 % ± 18.73 % | 0.2369 |
| 120 | 55.65 % ± 11.11 % | 0.0077 * | 235.78 % ± 41.25 % | 0.0079 * |
| 240 | 73.62 % ± 22.93 % | 0.4206 | 143.39 % ± 11.59 % | 0.0439 * |
| Species | Taxonomic Group | NR Specific Activity (U = µmol Nitrite/min) |
|---|---|---|
| Ramalina farinacea | Lichen (Chlorophyta − Trebouxiophyceae + Ascomycota) | (0.91 ± 0.13) × 10−4 U/mg present work |
| Ulva intestinalis | Chlorophyta − Chlorophyceae | 0.27 × 10−2 U/mg [53] |
| Physcomitrella patens | Bryophyta | 0.40 × 10−4 U/mg [45] |
| Porphyra perforata | Rhodophyta, Bangiophyceae | 8.33 × 10−4 U/mg [53] |
| Kappaphycus alvarezii | Rhodophyta, Gigartinales | 0.16 U/mg [54] |
| Gracilaria tenuistipitata | Rhodophyta, Gracilariales | 3.00 ± 0.20 (apical) U/mg [55] 1.60 ± 0.10 (basal) U/mg [55] |
| Gracilaria tikvahiae | Rhodophyta, Gracilariales | 0.43 × 10−4 U/mg [58] |
| Gracilaria chilensis | Rhodophyta, Gracilariales | (2.53 ± 0.03) × 10−4 U/mg [56] |
| Gracilaria caudata | Rhodophyta, Gracilariales | 0.93 × 10−4 U/mg [39] |
| Arabidopsis thaliana | Magnoliophyta | 2.50 × 10−4 U /mg [57] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Expósito, J.R.; Martín San Román, S.; Barreno, E.; Reig-Armiñana, J.; García-Breijo, F.J.; Catalá, M. Inhibition of NO Biosynthetic Activities during Rehydration of Ramalina farinacea Lichen Thalli Provokes Increases in Lipid Peroxidation. Plants 2019, 8, 189. https://doi.org/10.3390/plants8070189
Expósito JR, Martín San Román S, Barreno E, Reig-Armiñana J, García-Breijo FJ, Catalá M. Inhibition of NO Biosynthetic Activities during Rehydration of Ramalina farinacea Lichen Thalli Provokes Increases in Lipid Peroxidation. Plants. 2019; 8(7):189. https://doi.org/10.3390/plants8070189
Chicago/Turabian StyleExpósito, Joana R., Sara Martín San Román, Eva Barreno, José Reig-Armiñana, Francisco José García-Breijo, and Myriam Catalá. 2019. "Inhibition of NO Biosynthetic Activities during Rehydration of Ramalina farinacea Lichen Thalli Provokes Increases in Lipid Peroxidation" Plants 8, no. 7: 189. https://doi.org/10.3390/plants8070189
APA StyleExpósito, J. R., Martín San Román, S., Barreno, E., Reig-Armiñana, J., García-Breijo, F. J., & Catalá, M. (2019). Inhibition of NO Biosynthetic Activities during Rehydration of Ramalina farinacea Lichen Thalli Provokes Increases in Lipid Peroxidation. Plants, 8(7), 189. https://doi.org/10.3390/plants8070189