Responses of Aspen Leaves to Heatflecks: Both Damaging and Non-Damaging Rapid Temperature Excursions Reduce Photosynthesis

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
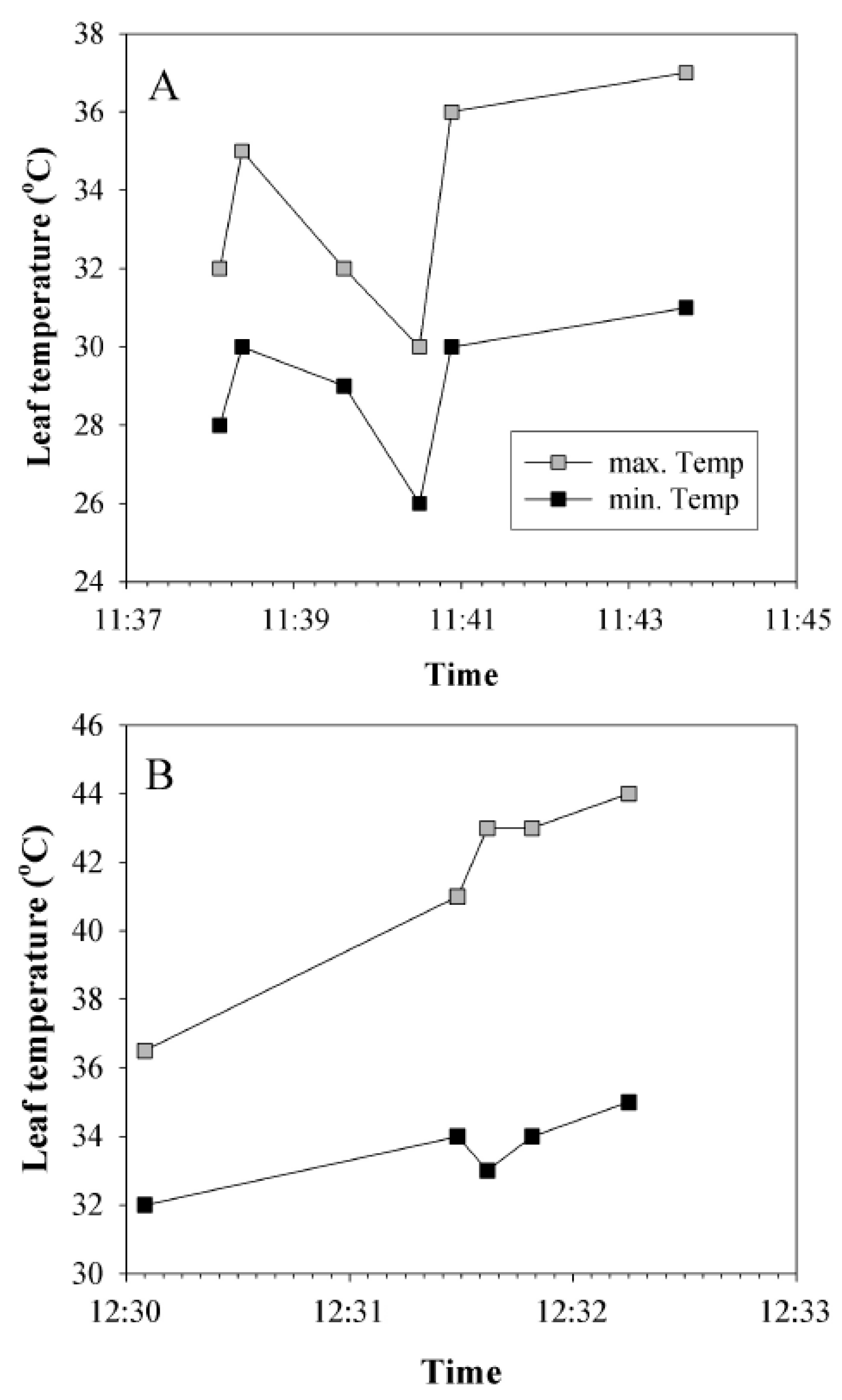
2.1. Assessment of Leaf Temperatures in the Field
2.2. Plant Material and Growth Conditions for Laboratory Experiments
2.3. Experimental Set-Up
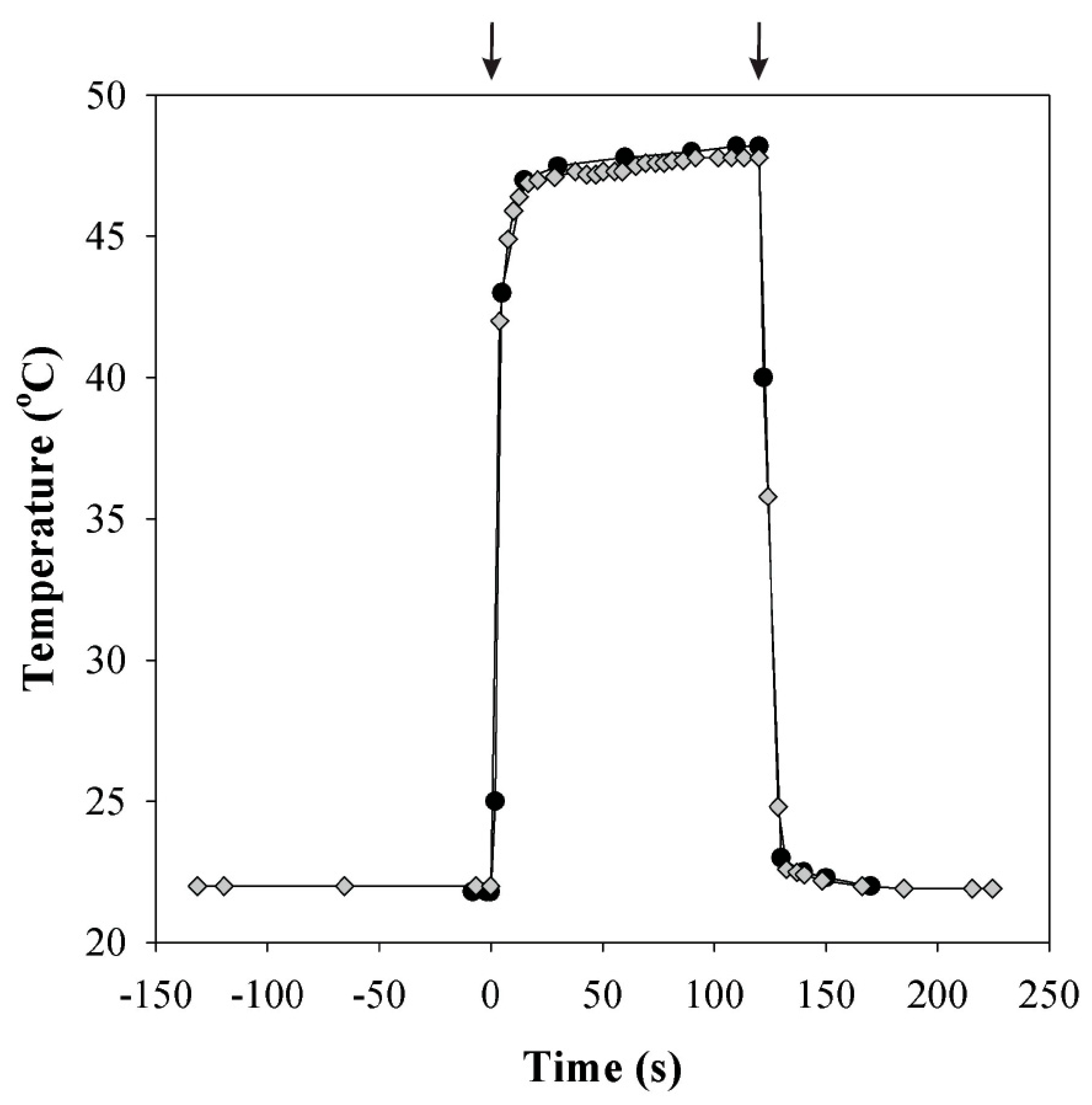
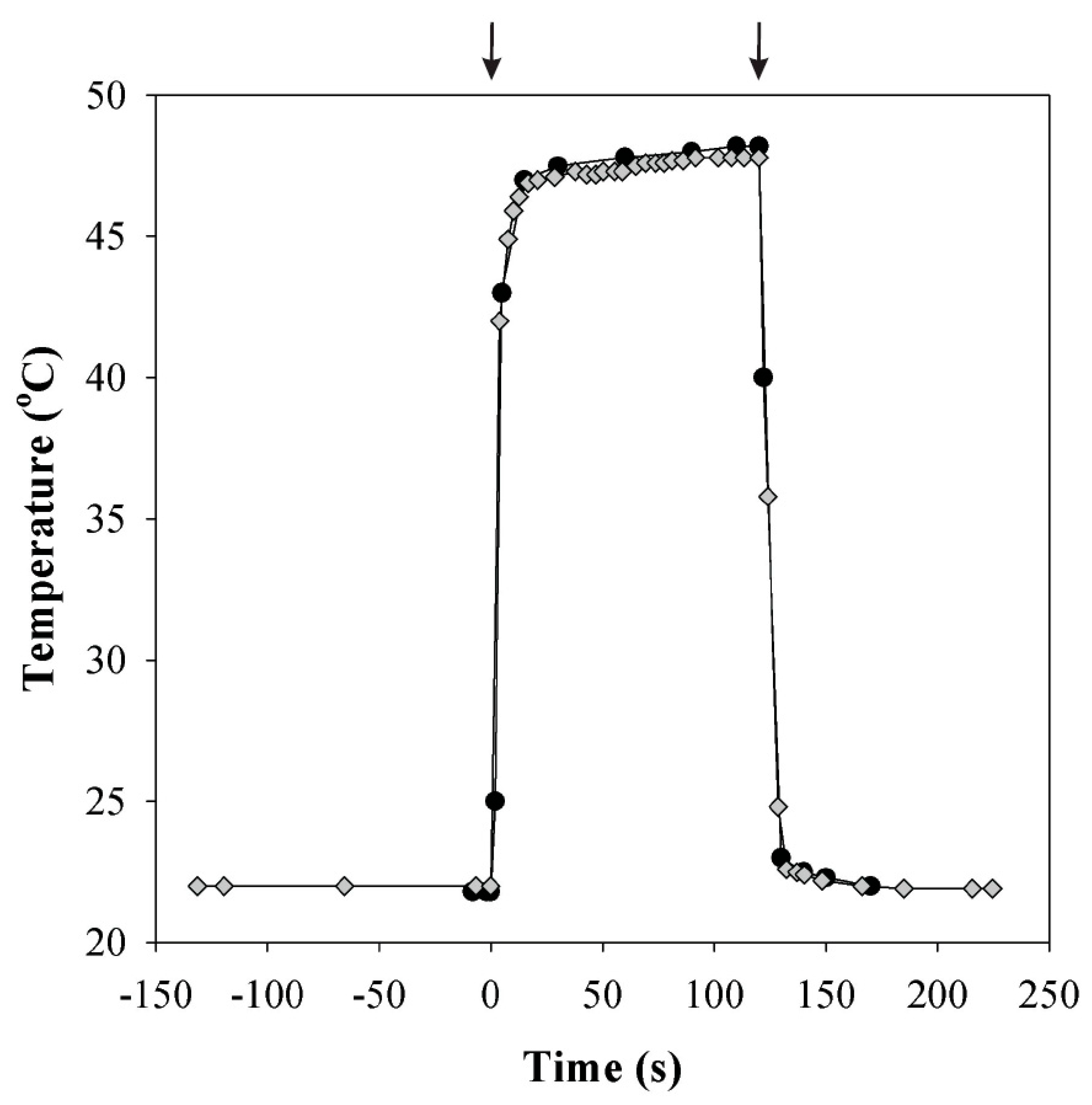
2.4. Temperature and Light Increase Experiments
2.5. Membrane Permeability Assays
2.6. Data Analyses
3. Results
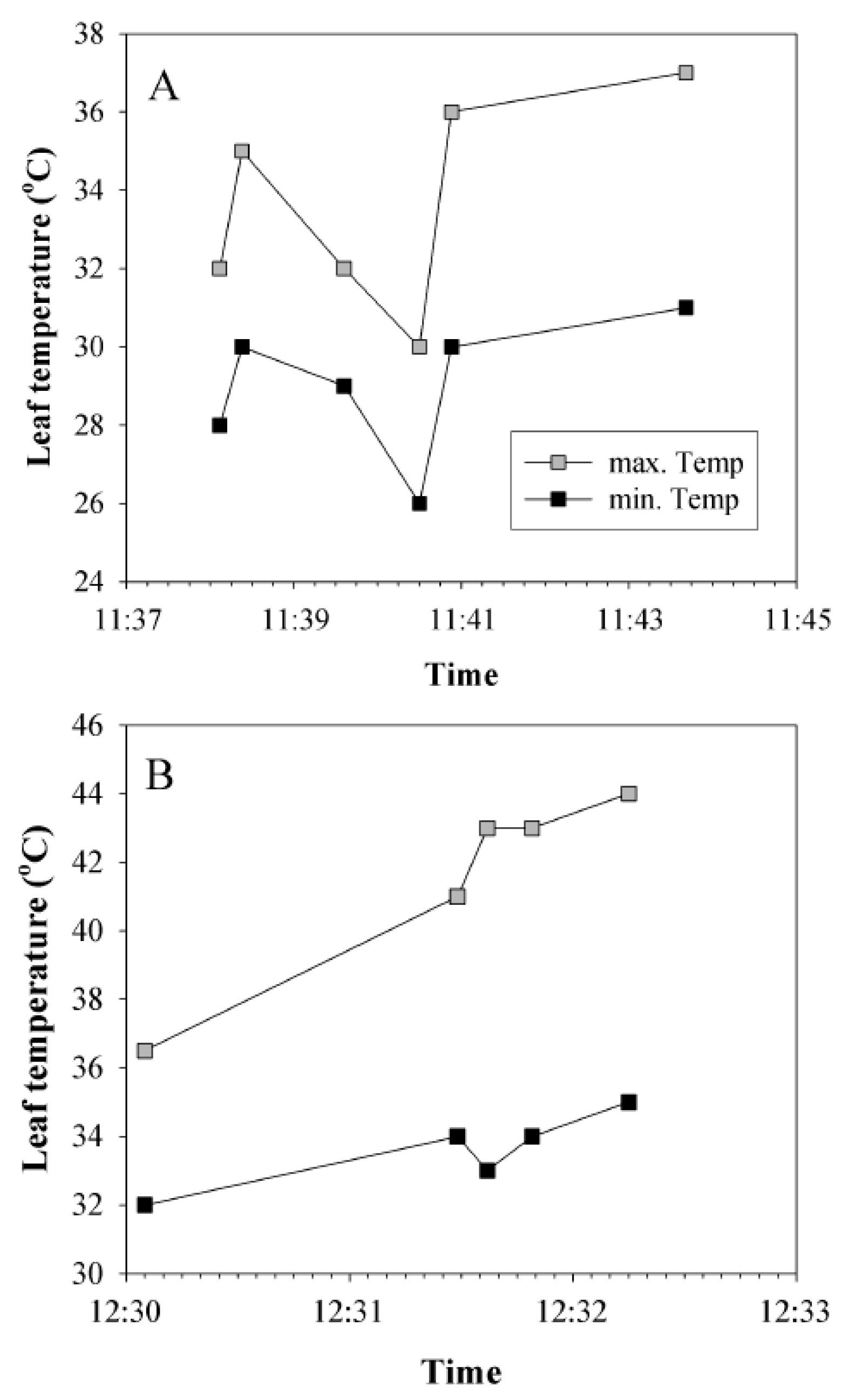
3.1. Leaf Temperatures in the Field
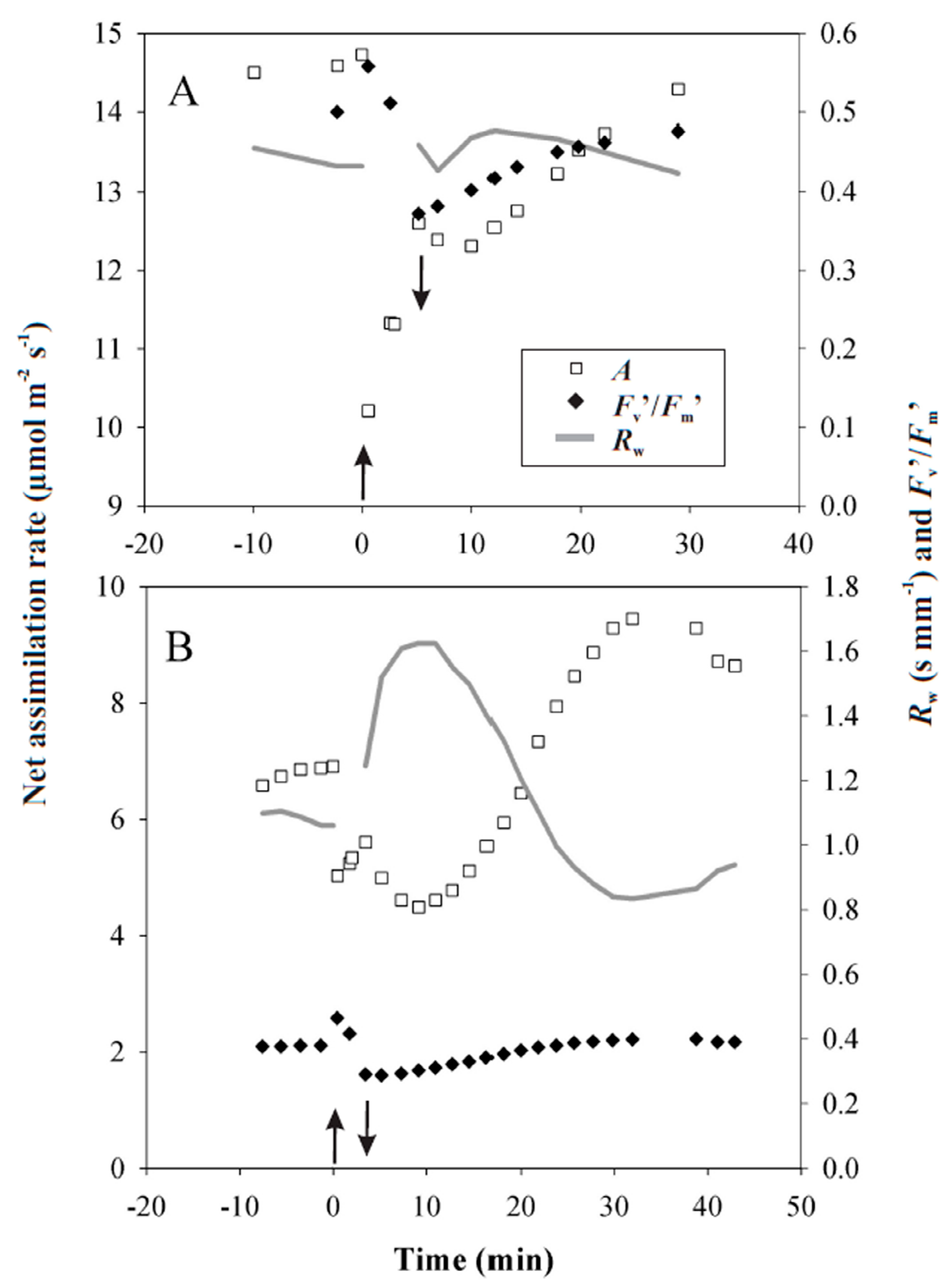
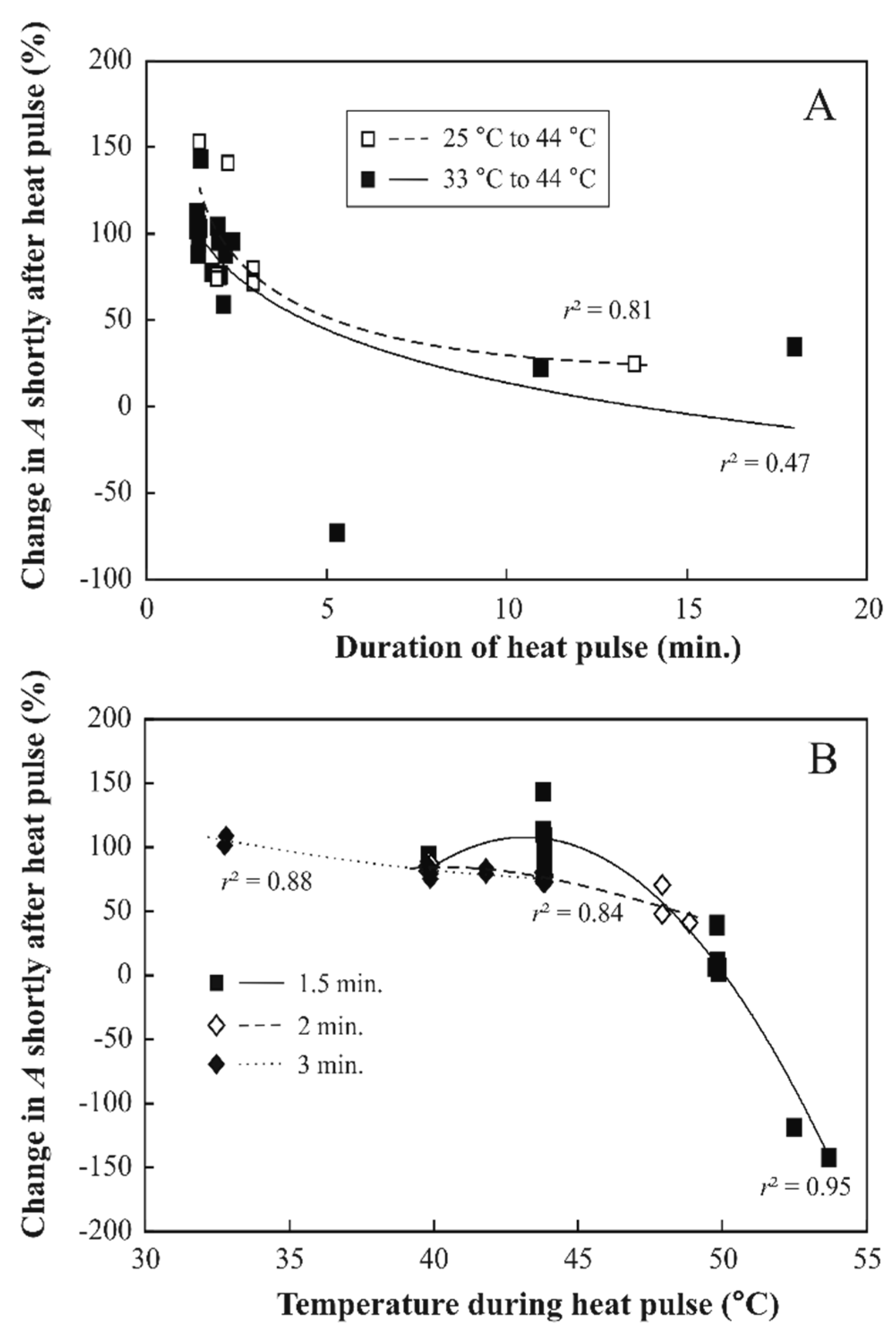
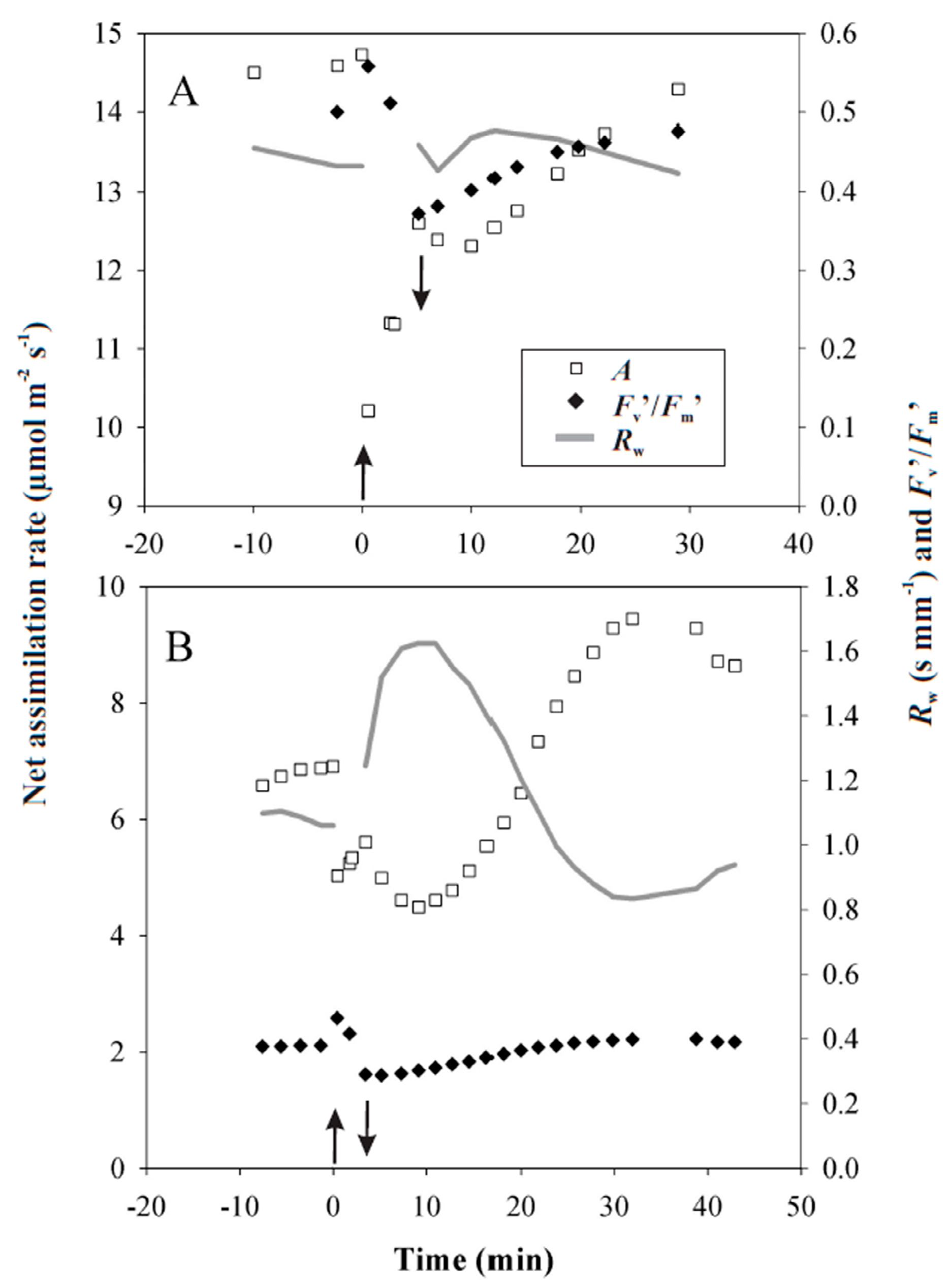
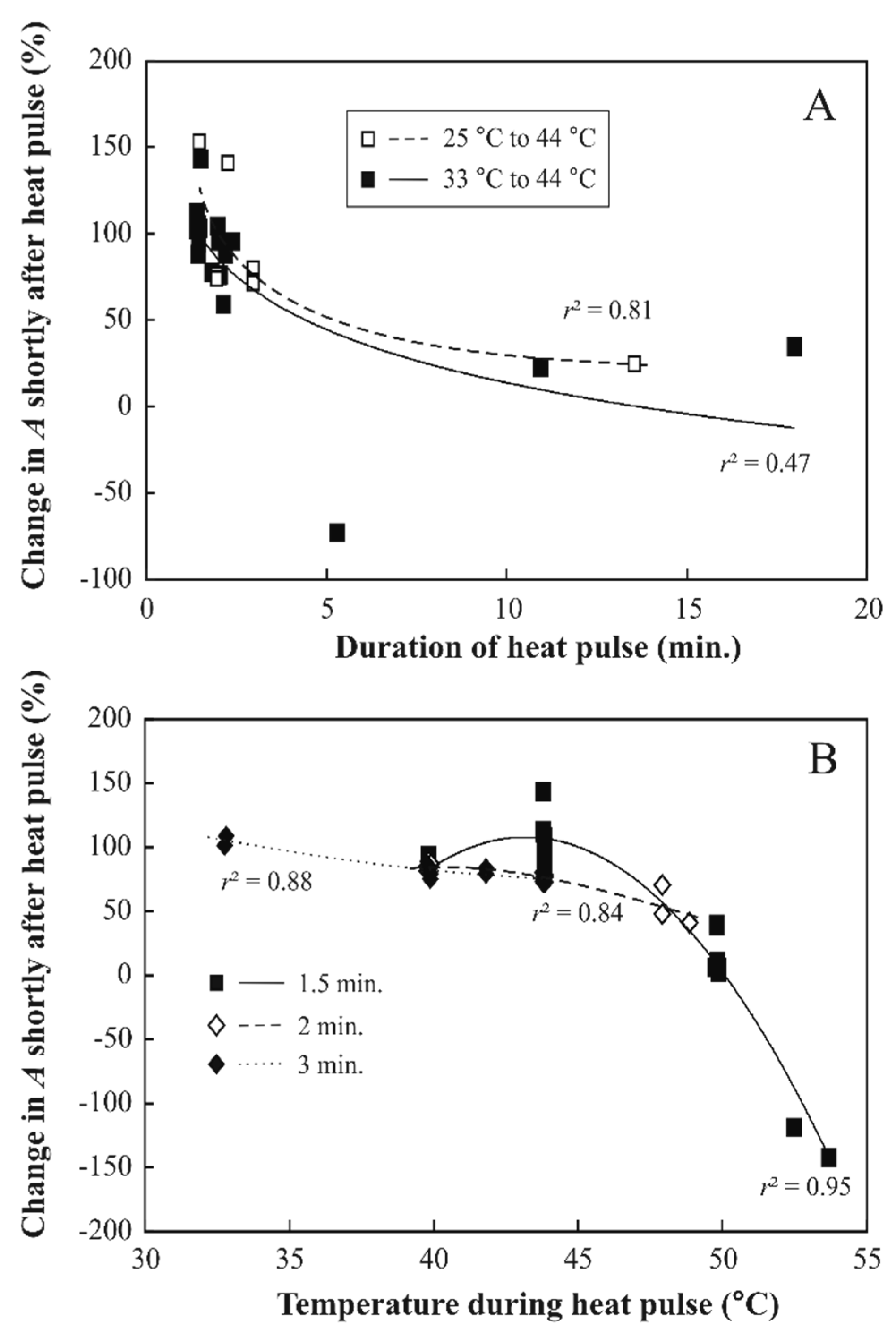
3.2. Responses of Photosynthesis to Heat Pulses Applied under Laboratory Conditions
3.3. Heat Pulses Combined with Higher Light Intensity
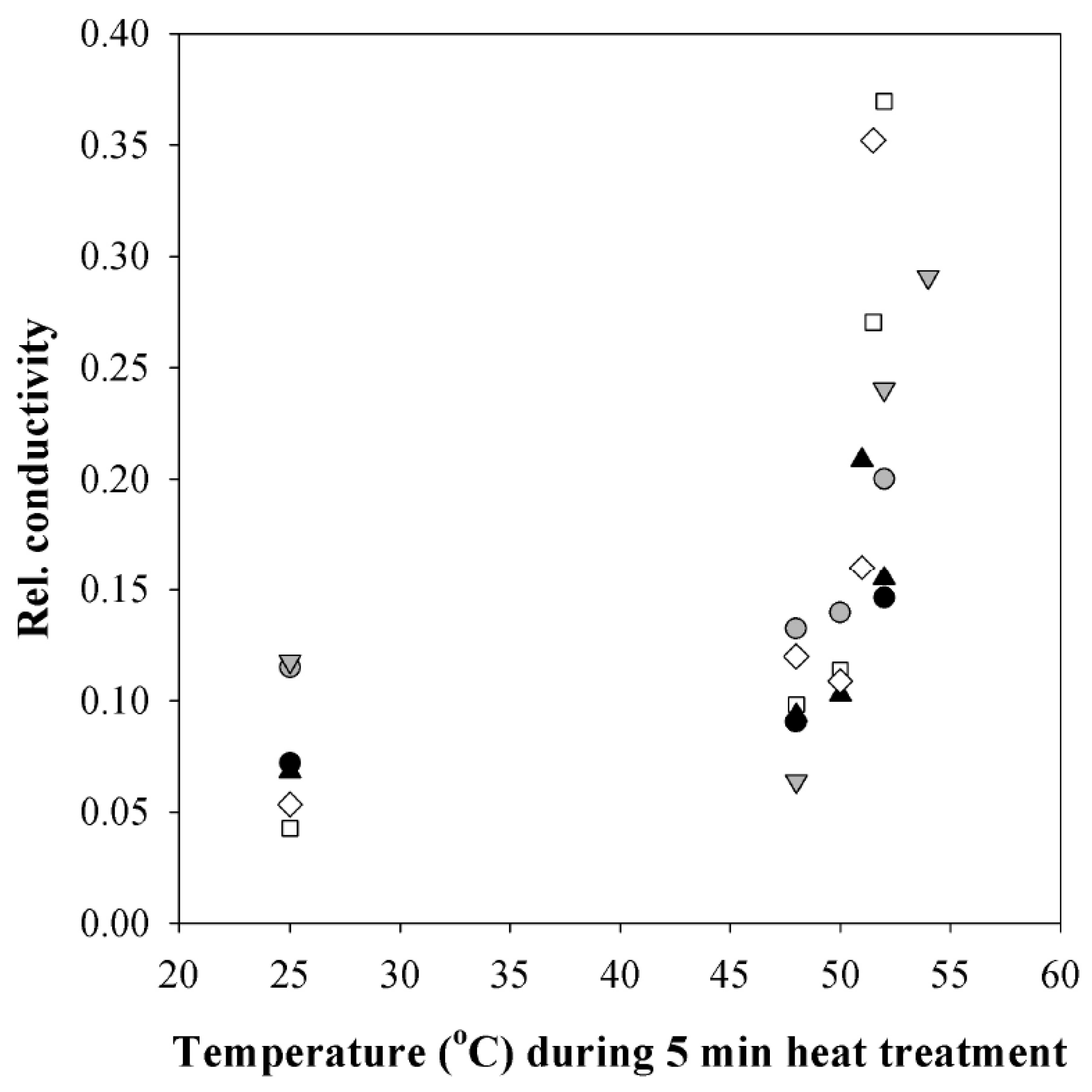
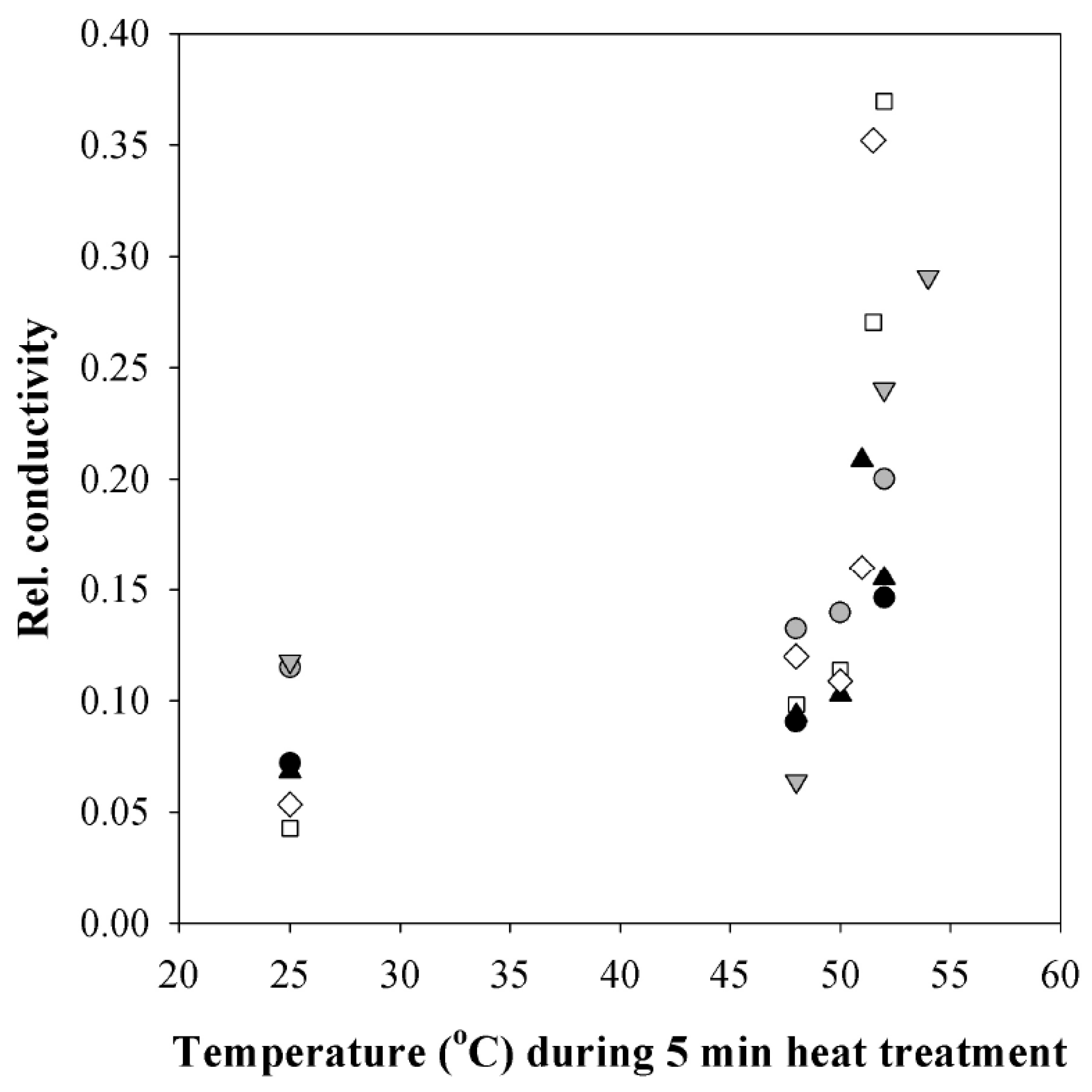
3.4. Changes in Plasmalemma Conductivity
4. Discussion
4.1. Leaf Temperatures in the Field
4.2. Reversible Changes in Photosynthetic Characteristics to Heat Pulses
4.3. Irreversible Photosynthetic Changes to Longer and Hotter Heat Pulses
4.4. Changes in Light Intensity during Heat Pulses Had Only A Minor Effect on Heat Resistance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| A: | net CO2 assimilation rate |
| F0: | dark-adapted minimum fluorescence yield |
| Fm: | dark-adapted maximum fluorescence yield |
| Fm´: | light-adapted maximum fluorescence yield |
| Fs´: | steady-state fluorescence yield |
| PSII: | photosystem II |
| Rw: | stomatal resistance to water vapor; |
| ΦPSII: | effective PSII quantum yield |
References
- Holbo, H.R.; Childs, S.W.; McNabb, D.H. Solar radiation at seedling sites below partial canopies. For. Ecol. Manag. 1985, 10, 115–124. [Google Scholar] [CrossRef]
- Pugnaire, F.I.; Valladares, F. The architecture of plant crowns: From design rules to light capture and performance. In Handbook of functional plant ecology, 2nd ed.; Pugnaire, F.I.; Valladares, F. CRC Press: Boca Raton, FL, USA, 2007; pp. 101–149. [Google Scholar]
- Porcar-Castell, A.; Palmroth, S. Modelling photosynthesis in highly dynamic environments: The case of sunflecks. Tree Physiol. 2012, 32, 1062–1065. [Google Scholar] [CrossRef] [PubMed]
- Way, D.A.; Pearcy, R.W. Sunflecks in trees and forests: From photosynthetic physiology to global change biology. Tree Physiol. 2012, 32, 1066–1081. [Google Scholar] [CrossRef] [PubMed]
- Pearcy, R.W. Sunflecks and photosynthesis in plant canopies. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1990, 41, 421–453. [Google Scholar] [CrossRef]
- Young, D.R.; Smith, W.K. Influence of sunflecks on the temperature and water relations of two subalpine understory congeners. Oecologia 1979, 43, 195–205. [Google Scholar] [CrossRef]
- Leakey, A.D.B.; Press, M.C.; Scholes, J.D. High-temperature inhibition of photosynthesis is greater under sunflecks than uniform irradiance in a tropical rain forest tree seedling. Plant Cell Environ. 2003, 26, 1681–1690. [Google Scholar] [CrossRef]
- Watling, J.R.; Robinson, S.A.; Woodrow, I.E.; Osmond, C.B. Responses of rainforest understorey plants to excess light during sunflecks. Aust. J. Plant Physiol. 1997, 24, 17–25. [Google Scholar] [CrossRef]
- Grace, J. Plant response to wind. Agric. Ecosyst. Environ. 1988, 22, 71–88. [Google Scholar] [CrossRef]
- Roden, J.S.; Pearcy, R.W. The effect of flutter on the temperature of poplar leaves and its implications for carbon gain. Plant Cell Environ. 1993, 16, 571–577. [Google Scholar] [CrossRef]
- Osmond, C.B.; Anderson, J.M.; Ball, M.C.; Egerton, J.G. Compromising efficiency: the molecular ecology of light-resource utilization in plants. In Physiological plant ecology. The 39th Symposium of the British Ecological Society held at the University of York, 7–9 September 1998; Press, M.C., Scholes, J.D., Barker, M.G., Eds.; Blackwell Science: Oxford, UK, 1999; pp. 1–24. [Google Scholar]
- Takahashi, S.; Murata, N. How do environmental stresses accelerate photoinhibition? Trends Plant Sci. 2008, 13, 178–182. [Google Scholar] [CrossRef]
- Gururani, M.A.; Venkatesh, J.; Phan Tran, L.-S. Regulation of photosynthesis during abiotic stress-induced photoinhibition. Mol. Plant 2015, 8, 1304–1320. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü.; Keenan, T.F. Photosynthetic responses to stress in Mediterranean evergreens: Mechanisms and models. Environ. Exp. Bot. 2014, 103, 24–41. [Google Scholar] [CrossRef]
- Singsaas, E.L.; Laporte, M.M.; Shi, J.-Z.; Monson, R.K.; Bowling, D.R.; Johnson, K.; Lerdau, M.; Jasentuliytana, A.; Sharkey, T.D. Kinetics of leaf temperature fluctuation affect isoprene emission from red oak (Quercus rubra) leaves. Tree Physiol. 1999, 19, 917–924. [Google Scholar] [CrossRef]
- Wise, R.R.; Olson, A.J.; Schrader, S.M.; Sharkey, T.D. Electron transport is the functional limitation of photosynthesis in field-grown Pima cotton plants at high temperature. Plant Cell Environ. 2004, 27, 717–724. [Google Scholar] [CrossRef]
- Michaletz, S.T.; Weiser, M.D.; Zhou, J.; Kaspari, M.; Helliker, B.R.; Enquist, B.J. Plant thermoregulation: Energetics, trait–environment interactions, and carbon economics. Trends Ecol. Evol. 2015, 30, 714–724. [Google Scholar] [CrossRef]
- Hamerlynck, E.P.; Knapp, A.K. Leaf-level responses to light and temperature in two co-occurring Quercus (Fagaceae) species: Implications for tree distribution patterns. Forest Ecol. Manag. 1994, 68, 149–159. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.T.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Haldimann, P.; Feller, U. Inhibition of photosynthesis by high temperature in oak (Quercus pubescens L.) leaves grown under natural conditions closely correlates with a reversible heat-dependent reduction of the activation state of ribulose-1,5-bisphosphate carboxylase/oxygenase. Plant Cell Environ. 2004, 27, 1169–1183. [Google Scholar] [CrossRef]
- Salvucci, M.E.; Crafts-Brandner, S.J. Inhibition of photosynthesis by heat stress: The activation state of Rubisco as a limiting factor in photosynthesis. Physiol. Plant. 2004, 120, 179–186. [Google Scholar] [CrossRef]
- Kim, K.; Portis, A.R., Jr. Temperature dependence of photosynthesis in Arabidopsis plants with modifications in Rubisco activase and membrane fluidity. Plant Cell Physiol. 2005, 46, 522–530. [Google Scholar] [CrossRef]
- Sharkey, T.D. Effects of moderate heat stress on photosynthesis: Importance of thylakoid reactions, Rubisco deactivation, reactive oxygen species, and thermotolerance provided by isoprene. Plant Cell Environ. 2005, 28, 269–277. [Google Scholar] [CrossRef]
- Hüve, K.; Bichele, I.; Rasulov, B.; Niinemets, Ü. When it is too hot for photosynthesis: Heat-induced instability of photosynthesis in relation to respiratory burst, cell permeability changes and H2O2 formation. Plant Cell Environ. 2011, 34, 113–126. [Google Scholar] [CrossRef]
- Zhu, L.; Bloomfield, K.J.; Hocart, C.H.; Egerton, J.J.G.; O’Sullivan, O.S.; Penillard, A.; Weerasinghe, L.K.; Atkin, O.K. Plasticity of photosynthetic heat tolerance in plants adapted to thermally contrasting biomes. Plant Cell Environ. 2018, 41, 1251–1262. [Google Scholar] [CrossRef]
- O’Sullivan, O.S.; Heskel, M.A.; Reich, P.B.; Tjoelker, M.G.; Weerasinghe, K.W.L.K.; Penillard, A.; Zhu, L.; Egerton, J.J.G.; Bloomfield, K.J.; Creek, D.; et al. Thermal limits of leaf metabolism across biomes. Global Change Biol. 2017, 23, 209–223. [Google Scholar]
- Berry, J.; Björkman, O. Photosynthetic response and adaptation to temperature in higher plants. Annu. Rev. Plant Physiol. 1980, 31, 491–543. [Google Scholar] [CrossRef]
- Bilger, H.W.; Schreiber, U.; Lange, O.L. Determination of leaf heat resistance: Comparative investigation of chlorophyll fluorescence changes and tissue necrosis methods. Oecologia 1984, 63, 256–262. [Google Scholar] [CrossRef]
- Seemann, J.R.; Berry, J.A.; Downton, W.J.S. Photosynthetic response and adaptation to high temperature in desert plants. A comparison of gas exchange and fluorescence methods for studies of thermal tolerance. Plant Physiol. 1984, 75, 364–368. [Google Scholar] [CrossRef]
- Schrader, S.M.; Wise, R.R.; Wacholtz, W.F.; Ort, D.R.; Sharkey, T.D. Thylakoid membrane responses to moderately high leaf temperature in Pima cotton. Plant Cell Environ. 2004, 27, 725–735. [Google Scholar] [CrossRef]
- Schrader, S.M.; Kleinbeck, K.R.; Sharkey, T.D. Rapid heating of intact leaves reveals initial effects of stromal oxidation on photosynthesis. Plant Cell Environ. 2007, 30, 671–678. [Google Scholar] [CrossRef]
- Raison, J.K.; Berry, J.A. Viscotropic denaturation of chloroplast membranes and acclimation to temperature by adjustment of lipid viscosity. Carnegie Inst. Wash. Yearb. 1979, 78, 149–152. [Google Scholar]
- Smith, K.A.; Low, P.S. Identification and partial characterization of the denaturation transition of the photosystem II reaction center of spinach chloroplast membranes. Plant Physiol. 1989, 90, 575–581. [Google Scholar] [CrossRef]
- Yamane, Y.; Kashino, Y.; Koike, H.; Satoh, K. Effects of high temperatures on the photosynthetic systems in spinach: Oxygen-evolving activities, fluorescence characteristics and the denaturation process. Photosynth. Res. 1998, 57, 51–59. [Google Scholar] [CrossRef]
- Niinemets, Ü. When leaves go over the thermal edge. Plant Cell Environ. 2018, 41, 1247–1250. [Google Scholar] [CrossRef]
- Copolovici, L.; Kännaste, A.; Pazouki, L.; Niinemets, Ü. Emissions of green leaf volatiles and terpenoids from Solanum lycopersicum are quantitatively related to the severity of cold and heat shock treatments. J. Plant Physiol. 2012, 169, 664–672. [Google Scholar] [CrossRef]
- Tullus, A.; Tullus, H.; Vares, A.; Kanal, A. Early growth of hybrid aspen (Populus x wettseteinii Hämet-Ahti) plantations on former agricultural lands in Estonia. Forest Ecol. Manag. 2007, 245, 118–129. [Google Scholar] [CrossRef]
- Tullus, A.; Tullus, H.; Soo, T.; Pärn, L. Above-ground biomass characteristics of young hybrid aspen (Populus tremula L. × P. tremuloides Michx.) plantations on former agricultural land in Estonia. Biomass Bioenergy 2009, 33, 1617–1625. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Oja, V.; Kull, O. Shape of leaf photosynthetic electron transport versus temperature response curve is not constant along canopy light gradients in temperate deciduous trees. Plant Cell Environ. 1999, 22, 1497–1514. [Google Scholar] [CrossRef]
- Laisk, A.; Oja, V.; Rasulov, B.; Rämma, H.; Eichelmann, H.; Kasparova, I.; Pettai, H.; Padu, E.; Vapaavuori, E. A computer-operated routine of gas exchange and optical measurements to diagnose photosynthetic apparatus in leaves. Plant Cell Environ. 2002, 25, 923–943. [Google Scholar] [CrossRef]
- Bajji, M.; Kinet, J.-M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Whitlow, T.H.; Bassuk, N.L.; Ranney, T.G.; Reichert, D.L. An improved method for using electrolyte leakage to assess membrane competence in plant tissues. Plant Physiol. 1992, 98, 198–205. [Google Scholar] [CrossRef]
- Scotti Campos, P.; Quartin, V.; Cochicho Ramalho, J.; Nunes, M.A. Electrolyte leakage and lipid degradation account for cold sensitivity in leaves of Coffea sp. J. Plant Physiol. 2003, 160, 283–292. [Google Scholar] [CrossRef]
- R Development Core Team. R: A language and environment for statistical computing.; The R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: http://www.R-project.org/ (accessed on 1 May 2019)ISBN 3-900051-07-0.
- Mulkey, S.S.; Pearcy, R.W. Interactions between acclimation and photoinhibition of photosynthesis of a tropical forest understorey herb, Alocasia macrorrhiza, during simulated canopy gap formation. Funct. Ecol. 1992, 6, 719–729. [Google Scholar] [CrossRef]
- Skillman, J.B.; Winter, K. High photosynthetic capacity in a shade-tolerant Crassulacean acid metabolism plant. Implications for sunfleck use, nonphotochemical energy dissipation, and susceptibility to photoinhibition. Plant Physiol. 1997, 113, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü.; Kollist, H.; García-Plazaola, J.I.; Hernández, A.; Becerril, J.M. Do the capacity and kinetics for modification of xanthophyll cycle pool size depend on growth irradiance in temperate trees? Plant Cell Environ. 2003, 26, 1787–1801. [Google Scholar] [CrossRef]
- Singsaas, E.L.; Sharkey, T.D. The regulation of isoprene emission responses to rapid leaf temperature fluctuations. Plant Cell Environ. 1998, 21, 1181–1188. [Google Scholar] [CrossRef]
- June, T.; Evans, J.R.; Farquhar, G.D. A simple equation for the reversible temperature dependence of photosynthetic electron transport: A study on soybean leaf. Funct. Plant Biol. 2004, 31, 275–283. [Google Scholar] [CrossRef]
- Rekika, D.; Kara, Y.; Souyris, I.; Nachit, M.M.; Asbati, A.; Monneveux, P. The tolerance of PSII to high temperatures in durum wheat (T. turgidum conv. durum): Genetic variation and relationship with yield under heat stress. Cereal Res. Commun. 2000, 28, 395–402. [Google Scholar]
- Havaux, M. Rapid photosynthetic adaptation to heat stress triggered in potato leaves by moderately elevated temperatures. Plant Cell Environ. 1993, 16, 461–467. [Google Scholar] [CrossRef]
- Königer, M.; Harris, G.C.; Pearcy, R.W. Interaction between photon flux density and elevated temperatures on photoinhibition in Alocasia macrorrhiza. Planta 1998, 205, 214–222. [Google Scholar] [CrossRef]
- Bilger, W.; Schreiber, U.; Lange, O.L. Chlorophyll fluorescence as an indicator of heat induced limitation of photosynthesis in Arbutus unedo L. In Plant response to stress. Functional analysis in Mediterranean ecosystems; Tenhunen, J.D., Catarino, F.M., Lange, O.L., Oechel, W.C., Eds.; Springer: Berlin, Germany, 1987; pp. 391–399. [Google Scholar]
- Hüve, K.; Bichele, I.; Ivanova, H.; Keerberg, O.; Pärnik, T.; Rasulov, B.; Tobias, M.; Niinemets, Ü. Temperature responses of dark respiration in relation to leaf sugar concentration. Physiol. Plant. 2012, 144, 320–334. [Google Scholar]
- Zhang, R.; Sharkey, T.D. Photosynthetic electron transport and proton flux under moderate heat stress. Photosynth. Res. 2009, 100, 29–43. [Google Scholar] [CrossRef]
- Zhang, R.; Cruz, J.A.; Kramer, D.M.; Magallanes-Lundback, M.E.; DellaPenna, D.; Sharkey, T.D. Moderate heat stress reduces the pH component of the transthylakoid proton motive force in light-adapted, intact tobacco leaves. Plant Cell Environ. 2009, 32, 1538–1547. [Google Scholar] [CrossRef]
- Eliáš, P. Stomatal oscillations in adult forest trees in natural environment. Biol. Plant. 1979, 21, 71–74. [Google Scholar] [CrossRef]
- Laisk, A.; Oja, V.; Walker, D.; Heber, U. Oscillations in photosynthesis and reduction of photosystem I acceptor side in sunflower leaves. Functional cytochrome b6/f-photosystem I ferredoxin-NADP reductase supercomplexes. Photosynthetica 1992, 27, 465–479. [Google Scholar]
- Rasulov, B.; Talts, E.; Niinemets, Ü. Spectacular oscillations in plant isoprene emission under transient conditions explain the enigmatic CO2 response. Plant Physiol. 2016, 172, 2275–2285. [Google Scholar] [CrossRef]
- Steppe, K.; Dzikiti, S.; Lemeur, R.; Milford, J.R. Stomatal oscillations in orange trees under natural climatic conditions. Ann. Bot. 2006, 97, 831–835. [Google Scholar] [CrossRef]
- Kask, K.; Kännaste, A.; Talts, E.; Copolovici, L.; Niinemets, Ü. How specialized volatiles respond to chronic and short-term physiological and shock heat stress in Brassica nigra. Plant Cell Environ. 2016, 39, 2027–2042. [Google Scholar] [CrossRef]
- Pazouki, L.; Kanagendran, A.; Li, S.; Kännaste, A.; Rajabi Memari, H.; Bichele, R.; Niinemets, Ü. Mono- and sesquiterpene release from tomato (Solanum lycopersicum) leaves upon mild and severe heat stress and through recovery: From gene expression to emission responses. Environ. Exp. Bot. 2016, 132, 1–15. [Google Scholar] [CrossRef]
- Wright, I.J.; Ning, D.; Maire, V.; Prentice, I.C.; Westoby, M.; Díaz, S.; Gallagher, R.V.; Jacobs, B.F.; Kooyman, R.; Law, E.A.; et al. Global climatic drivers of leaf size. Science 2017, 357, 917–921. [Google Scholar] [CrossRef] [Green Version]
- Pshybytko, N.L.; Kruk, J.; Kabashnikova, L.F.; Strzalka, K. Function of plastoquinone in heat stress reactions of plants. Biochim. Biophys. Acta 2008, 1777, 1393–1399. [Google Scholar] [CrossRef] [Green Version]
- Laisk, A.; Rasulov, B.H.; Loreto, F. Thermoinhibition of photosynthesis as analyzed by gas exchange and chlorophyll fluorescence. Russ. J. Plant Physiol. 1998, 45, 412–421. [Google Scholar]
- Kubien, D.S.; Sage, R.F. The temperature response of photosynthesis in tobacco with reduced amounts of Rubisco. Plant Cell Environ. 2008, 31, 407–418. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Badger, M.R.; von Caemmerer, S.; Andrews, T.J. Increased heat sensitivity of photosynthesis in tobacco plants with reduced Rubisco activase. Photosynth. Res. 2001, 67, 147–156. [Google Scholar] [CrossRef]
- Tóth, S.Z.; Schansker, G.; Kissimon, J.; Kovács, L.; Garab, G.; Strasser, R.J. Biophysical studies of photosystem II-related recovery processes after a heat pulse in barley seedlings (Hordeum vulgare L.). J. Plant Physiol. 2005, 162, 181–194. [Google Scholar] [CrossRef]
- Havaux, M.; Greppin, H.; Strasser, R.J. Functioning of photosystems I and II in pea leaves exposed to heat stress in the presence or absence of light. Analysis using in-vivo fluorescence, absorbance, oxygen and photoacoustic measurements. Planta 1991, 186, 88–98. [Google Scholar] [CrossRef]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hüve, K.; Bichele, I.; Kaldmäe, H.; Rasulov, B.; Valladares, F.; Niinemets, Ü. Responses of Aspen Leaves to Heatflecks: Both Damaging and Non-Damaging Rapid Temperature Excursions Reduce Photosynthesis. Plants 2019, 8, 145. https://doi.org/10.3390/plants8060145
Hüve K, Bichele I, Kaldmäe H, Rasulov B, Valladares F, Niinemets Ü. Responses of Aspen Leaves to Heatflecks: Both Damaging and Non-Damaging Rapid Temperature Excursions Reduce Photosynthesis. Plants. 2019; 8(6):145. https://doi.org/10.3390/plants8060145
Chicago/Turabian StyleHüve, Katja, Irina Bichele, Hedi Kaldmäe, Bahtijor Rasulov, Fernando Valladares, and Ülo Niinemets. 2019. "Responses of Aspen Leaves to Heatflecks: Both Damaging and Non-Damaging Rapid Temperature Excursions Reduce Photosynthesis" Plants 8, no. 6: 145. https://doi.org/10.3390/plants8060145