An Exploration of the Roles of Ferric Iron Chelation-Strategy Components in the Leaves and Roots of Maize Plants

 ,
,  and
and 
Abstract
1. Introduction
2. Results
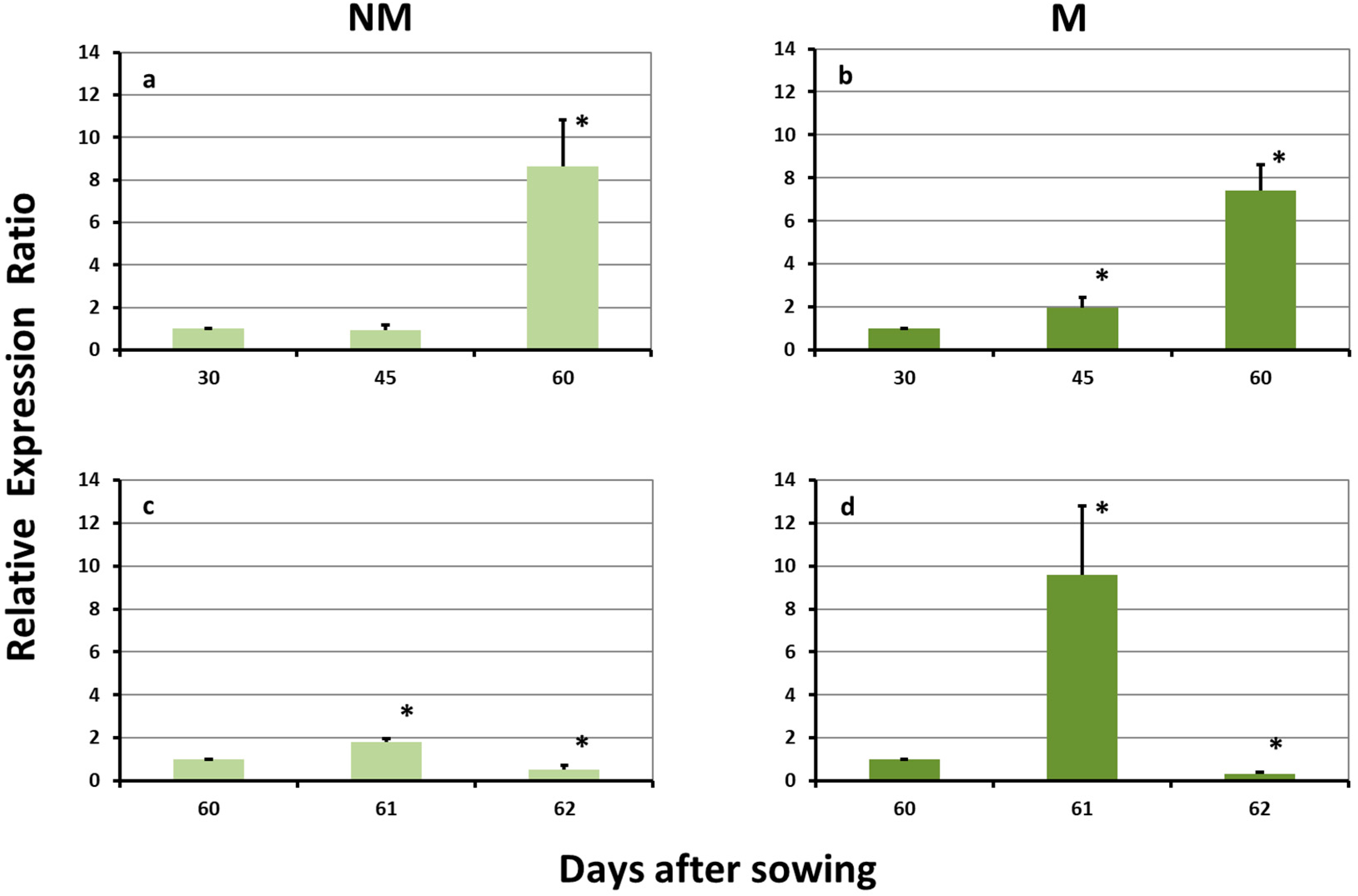
2.1. Expression Levels of ZmDMAS1 in the Leaves
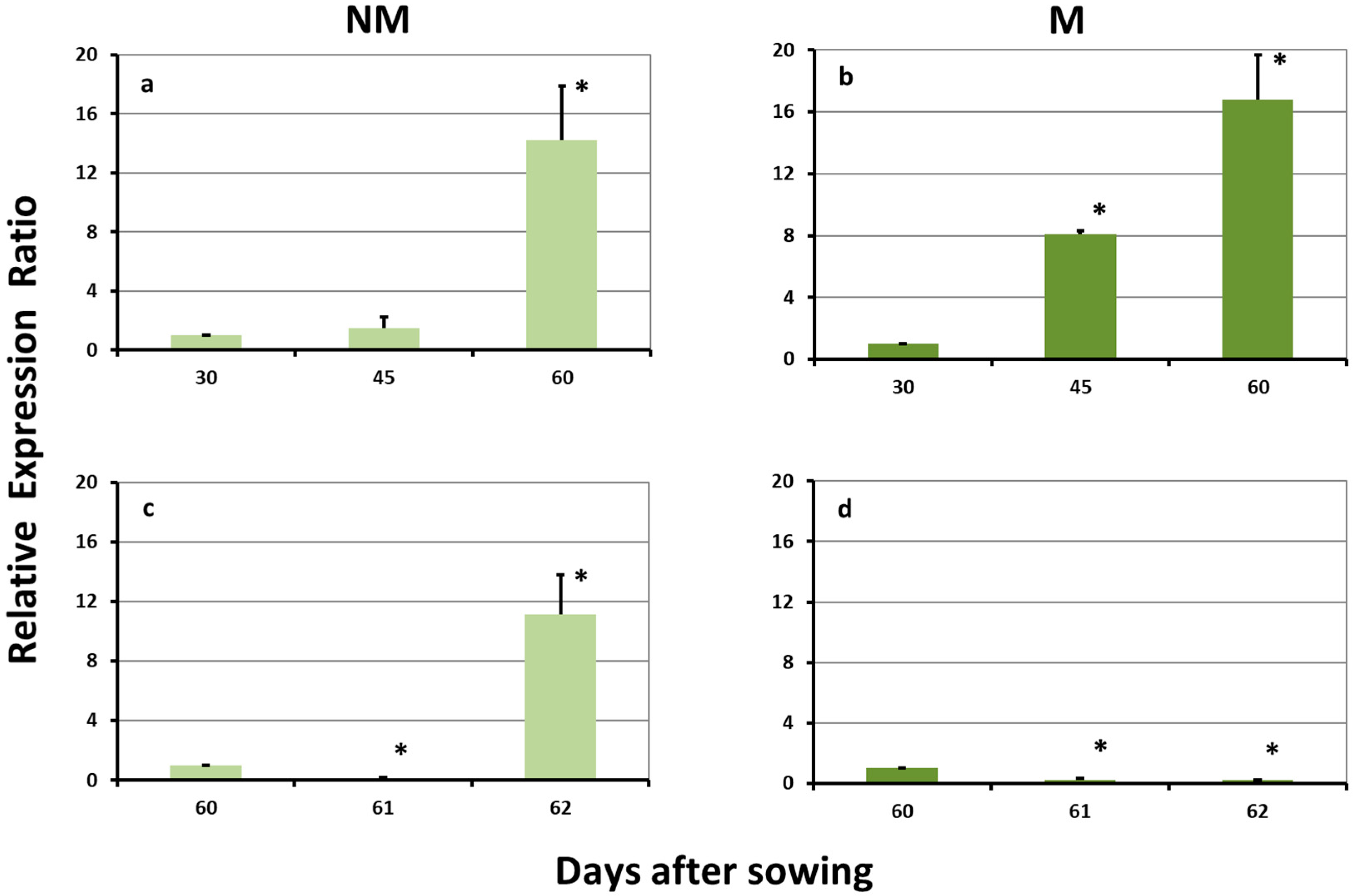
2.2. Expression Levels of ZmTOM1 in the Leaves
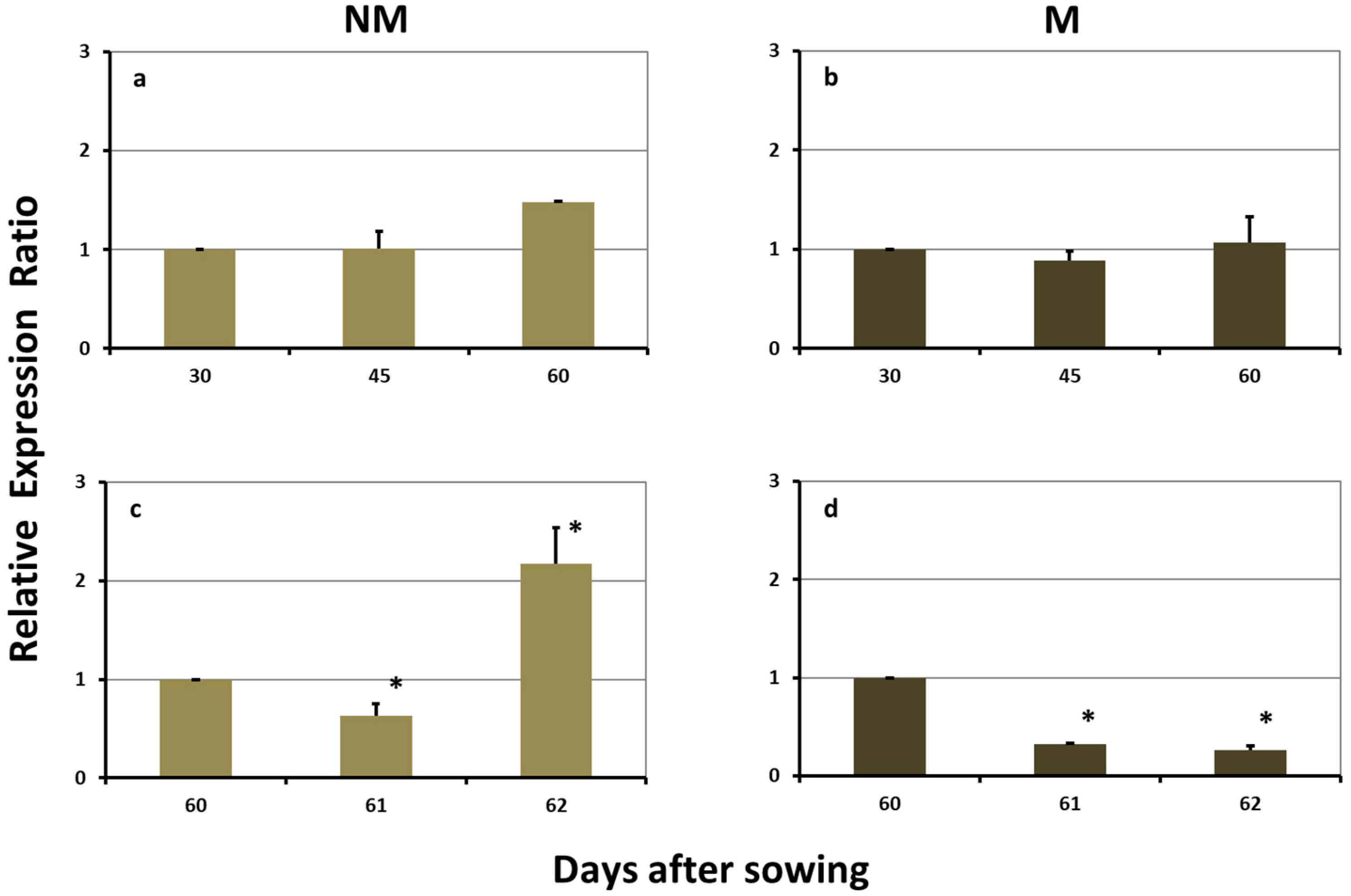
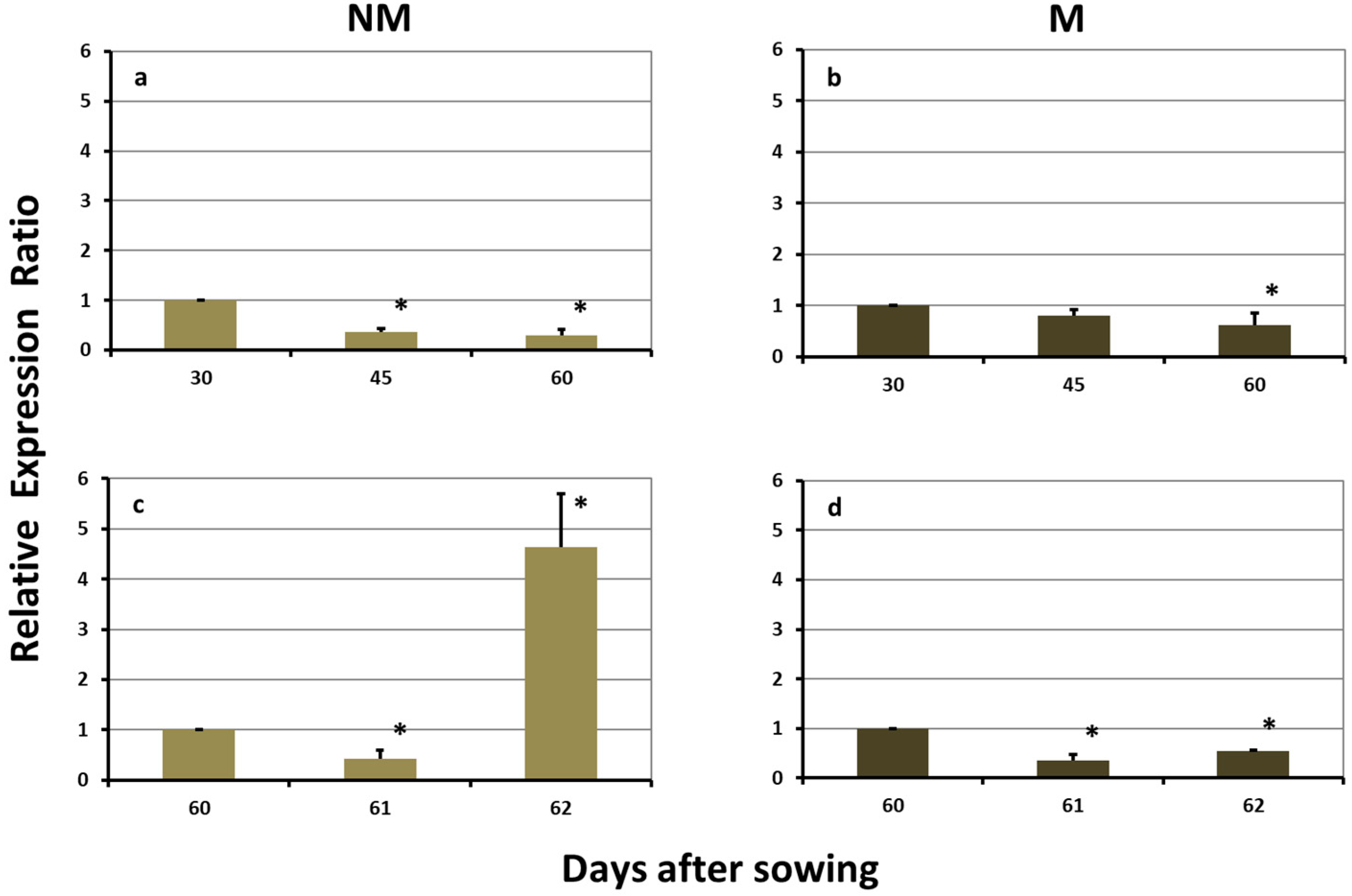
2.3. Expression Levels of ZmYS1 in the Leaves
2.4. Expression Levels of ZmDMAS1 in the Roots
2.5. Expression Levels of ZmTOM1 in the Roots
2.6. Data Meta-Analysis
3. Discussion
3.1. Roles of Ferric Iron Chelation Components in Maize Leaves
3.2. Arbuscular Mycorrhizal Fungus Provides Iron to M Plants
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Plant Samplings
4.3. Gene Expression Analysis
4.4. Data Meta-Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Connorton, J.M.; Balk, J.; Rodriguez-Celma, J. Iron homeostasis in plants—A brief overview. Metallomics 2017, 9, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yu, S.; Strong, J.; Wang, H. Are the biogeochemical cycles of carbon, nitrogen, sulfur, and phosphorus driven by the “FeIII-FeII redox wheel” in dynamic redox environments? J. Soils Sediments 2012, 12, 683–693. [Google Scholar] [CrossRef]
- Bashir, K.; Inoue, H.; Nagasaka, S.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Cloning and characterization of deoxymugineic acid synthase genes from graminaceous plants. J. Biol. Chem. 2006, 281, 32395–32402. [Google Scholar] [CrossRef] [PubMed]
- Ariga, Τ.; Hazama, Κ.; Yanagisawa, S.; Yoneyama, T. Chemical forms of iron in xylem sap from graminaceous and non-graminaceous plants. Soil Sci. Plant Nutr. 2014, 60, 460–469. [Google Scholar] [CrossRef]
- Chorianopoulou, S.N.; Saridis, Y.I.; Dimou, M.; Katinakis, P.; Bouranis, D.L. Arbuscular mycorrhizal symbiosis alters the expression patterns of three key iron homeostasis genes, ZmNAS1, ZmNAS3, and ZmYS1, in S deprived maize plants. Front. Plant Sci. 2015, 6, 257. [Google Scholar] [CrossRef] [PubMed]
- Saridis, G.I.; Chorianopoulou, S.N.; Katinakis, P.; Bouranis, D.L. Evidence for regulation of the iron uptake pathway by sulfate supply in S-deprived maize plants. In Sulfur Metabolism in Higher Plants—Fundamental, Environmental and Agricultural Aspects, Proceedings of the International Plant Sulfur Workshop, Goslar, Germany, 1–4 September 2015; De Kok, L.J., Hawkesford, M.J., Haneklaus, S.H., Schnug, E., Eds.; Springer International Publishing AG.: Berlin/Heidelberg, Germany, 2017; pp. 175–180. [Google Scholar]
- Nozoye, T.; Nagasaka, S.; Kobayashi, T.; Takahashi, M.; Sato, Y.; Sato, Y.; Uozumi, N.; Nakanishi, H.; Nishizawa, N.K. Phytosiderophore efflux transporters are crucial for iron acquisition in graminaceous plants. J. Biol. Chem. 2011, 86, 5446–5454. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.A.; Pierson, A.J.; Panaviene, Z.; Walker, E.L. Yellow Stripe1. Expanded roles for the maize iron-phytosiderophore transporter. Plant Physiol. 2004, 135, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Ueno, D.; Yamaji, N.; Ma, J.F. Further characterization of ferric—Phytosiderophore transporters ZmYS1 and HvYS1 in maize and barley. J. Exp. Bot. 2009, 60, 3513–3520. [Google Scholar] [CrossRef] [PubMed]
- Fodor, F. Heavy metals competing with iron under conditions involving phytoremediation. In Iron Nutrition in Plants and Rhizospheric Microorganisms; Barton, L.L., Abadía, J., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 129–151. [Google Scholar]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.; Karlen, Y.; Bakker, O.; van den Hoff, M.J.; Moorman, A.F. Amplification efficiency: Linking base line and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
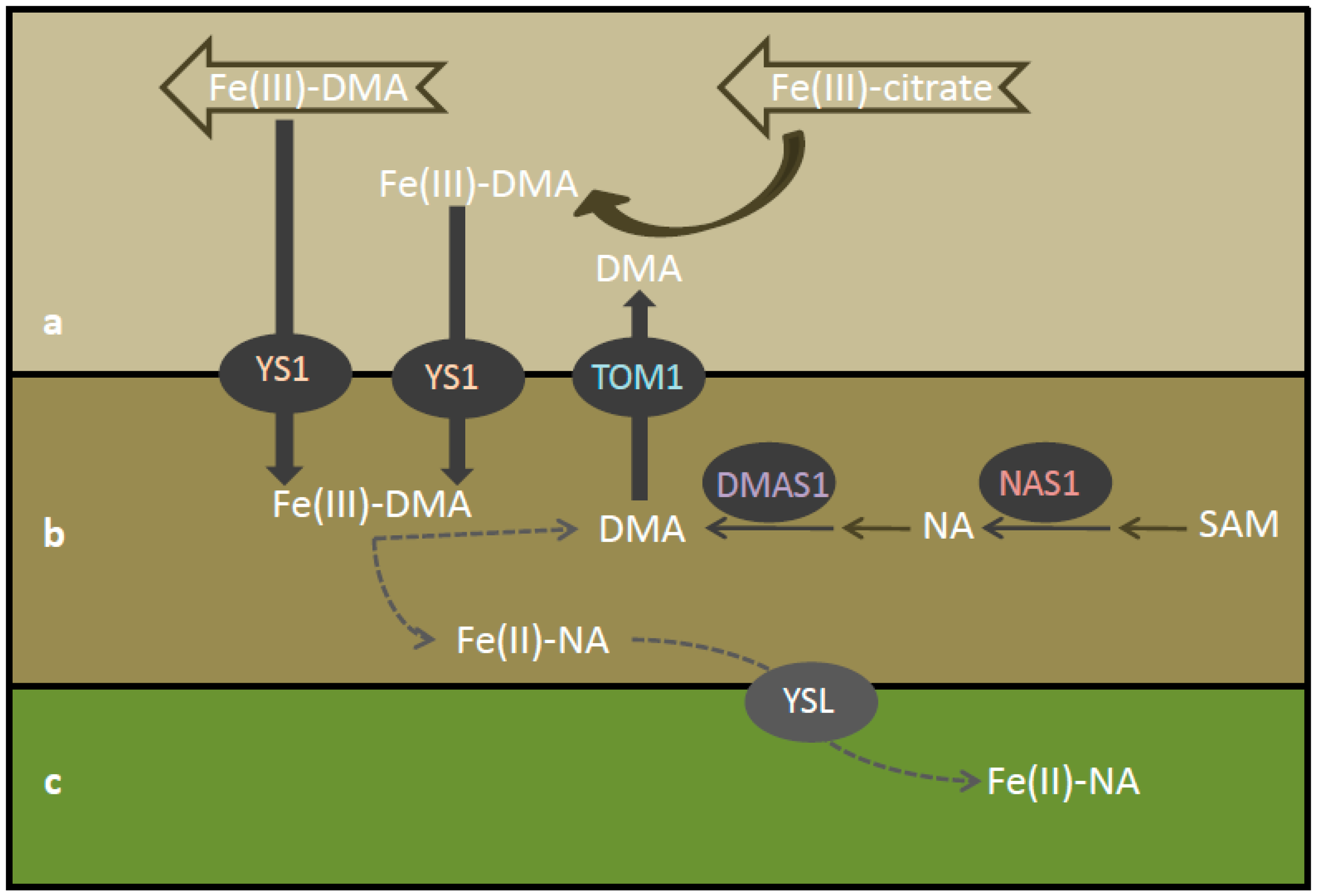{kind=link}
| Days after Sowing | ||||||
|---|---|---|---|---|---|---|
| Gene | 30 | 45 | 60 | 61 | 62 | |
| Leaves | ZmNAS3 | 0.91 ± 0.34 | 1.28 ± 0.12 | 5.32 ± 1.25 | 2.85 ± 0.16 | 0.45 ± 0.11 |
| ZmNAS1 | 2.01 ± 0.27 | 2.69 ± 0.43 | 1.13 ± 0.26 | 1.19 ± 0.20 | 0.55 ± 0.03 | |
| ZmDMAS1 | 0.61 ± 0.16 | 1.37 ± 0.21 | 1.80 ± 0.09 | 0.47 ± 0.03 | 1.27 ± 0.15 | |
| ZmTOM1 | 1.99 ± 0.12 | 9.04 ± 1.68 | 4.12 ± 1.09 | 13.76 ± 3.54 | 3.31 ± 0.36 | |
| ZmYS1 | 0.11 ± 0.06 | 1.76 ± 0.21 | 0.35 ± 0.14 | 0.28 ± 0.13 | 0.03 ± 0.01 | |
| Roots | ZmNAS3 | 0.83 ± 0.13 | 1.94 ± 0.11 | 0.99 ± 0.07 | 1.33 ± 0.31 | 0.67 ± 0.12 |
| ZmNAS1 | 0.96 ± 0.22 | 0.73 ± 0.08 | 0.63 ± 0.05 | 1.76 ± 0.12 | 0.41 ± 0.07 | |
| ZmDMAS1 | 1.12 ± 0.18 | 1.10 ± 0.25 | 0.85 ± 0.20 | 0.48 ± 0.09 | 0.41 ± 0.15 | |
| ZmTOM1 | 0.18 ± 0.10 | 0.38 ± 0.06 | 0.48 ± 0.14 | 0.45 ± 0.16 | 0.12 ± 0.02 | |
| ZmYS1 | 1.56 ± 0.31 | 0.78 ± 0.04 | 1.36 ± 0.19 | 1.05 ± 0.08 | 0.12 ± 0.08 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saridis, G.; Chorianopoulou, S.N.; Ventouris, Y.E.; Sigalas, P.P.; Bouranis, D.L. An Exploration of the Roles of Ferric Iron Chelation-Strategy Components in the Leaves and Roots of Maize Plants. Plants 2019, 8, 133. https://doi.org/10.3390/plants8050133
Saridis G, Chorianopoulou SN, Ventouris YE, Sigalas PP, Bouranis DL. An Exploration of the Roles of Ferric Iron Chelation-Strategy Components in the Leaves and Roots of Maize Plants. Plants. 2019; 8(5):133. https://doi.org/10.3390/plants8050133
Chicago/Turabian StyleSaridis, Georgios, Styliani N. Chorianopoulou, Yannis E. Ventouris, Petros P. Sigalas, and Dimitris L. Bouranis. 2019. "An Exploration of the Roles of Ferric Iron Chelation-Strategy Components in the Leaves and Roots of Maize Plants" Plants 8, no. 5: 133. https://doi.org/10.3390/plants8050133
APA StyleSaridis, G., Chorianopoulou, S. N., Ventouris, Y. E., Sigalas, P. P., & Bouranis, D. L. (2019). An Exploration of the Roles of Ferric Iron Chelation-Strategy Components in the Leaves and Roots of Maize Plants. Plants, 8(5), 133. https://doi.org/10.3390/plants8050133