Lipophilic Metabolites and Anatomical Acclimatization of Cleome amblyocarpa in the Drought and Extra-Water Areas of the Arid Desert of UAE

 ,
, 
Abstract
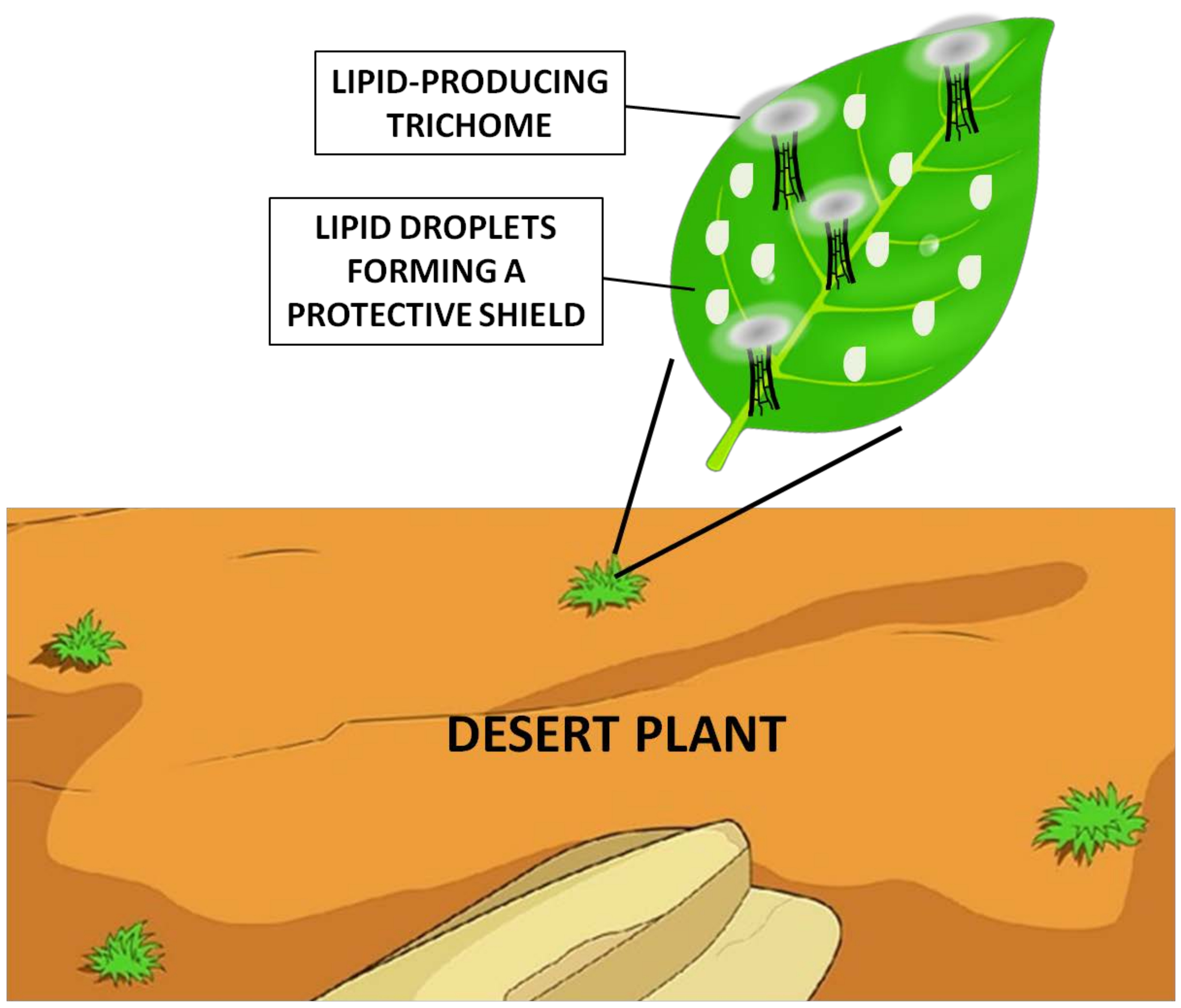{kind=link}
{kind=link}
{kind=link}
{kind=link}
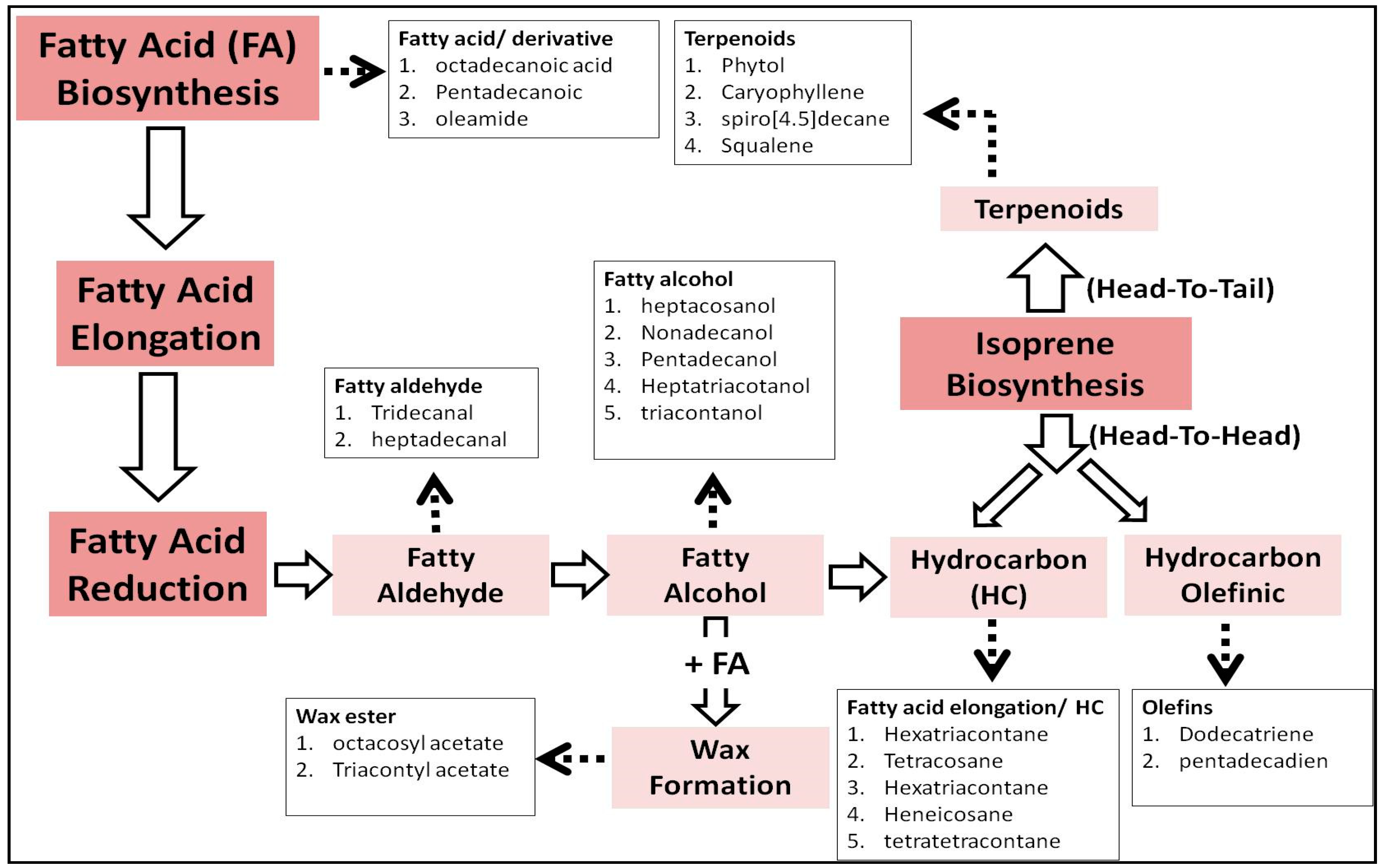{kind=link}
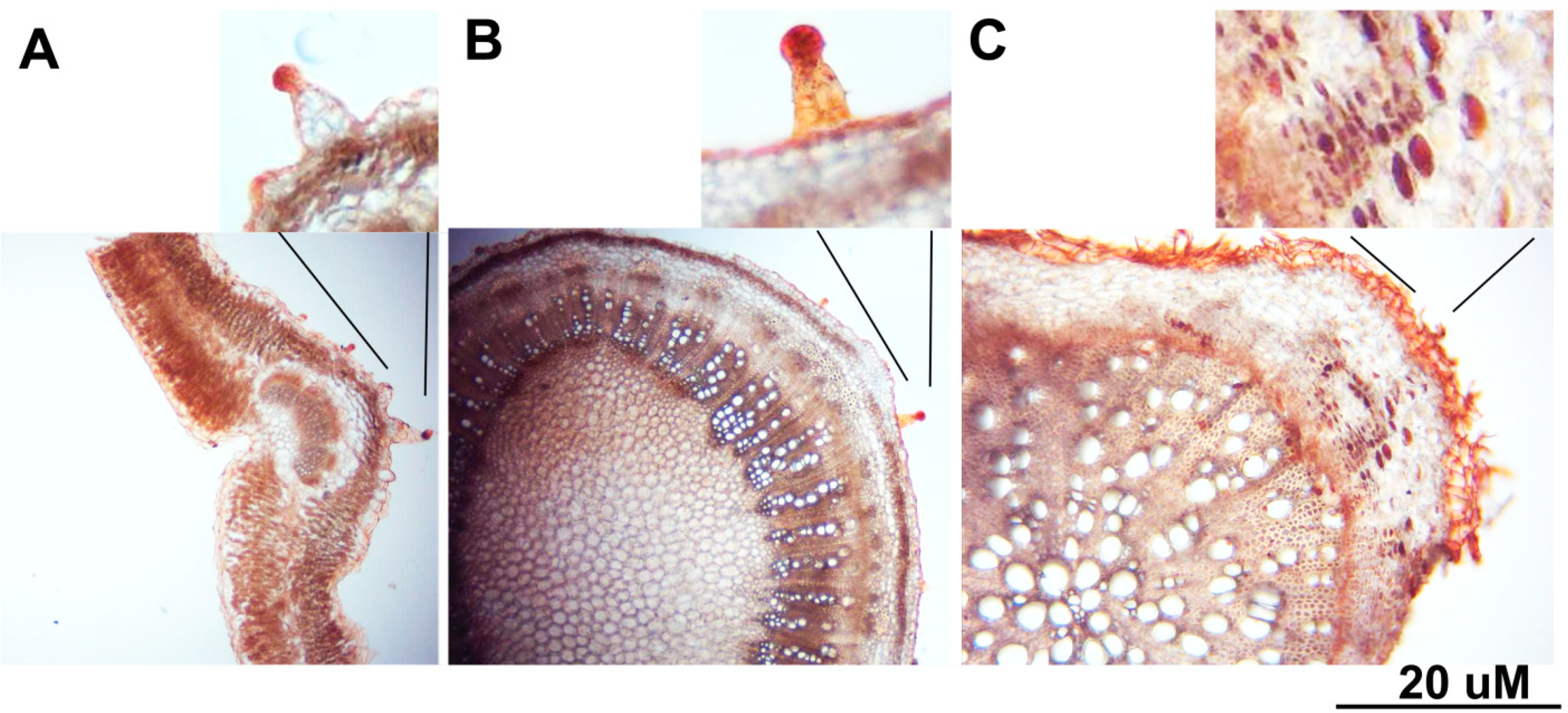{kind=link}
1. Introduction
2. Material and Methods
2.1. Plant Material
2.2. Lipophilic Metabolites Extraction
2.3. GC-MS of C. amblyocarpa Plants Collected from Both Sites
2.4. Light Microscopy
3. Results and Discussion
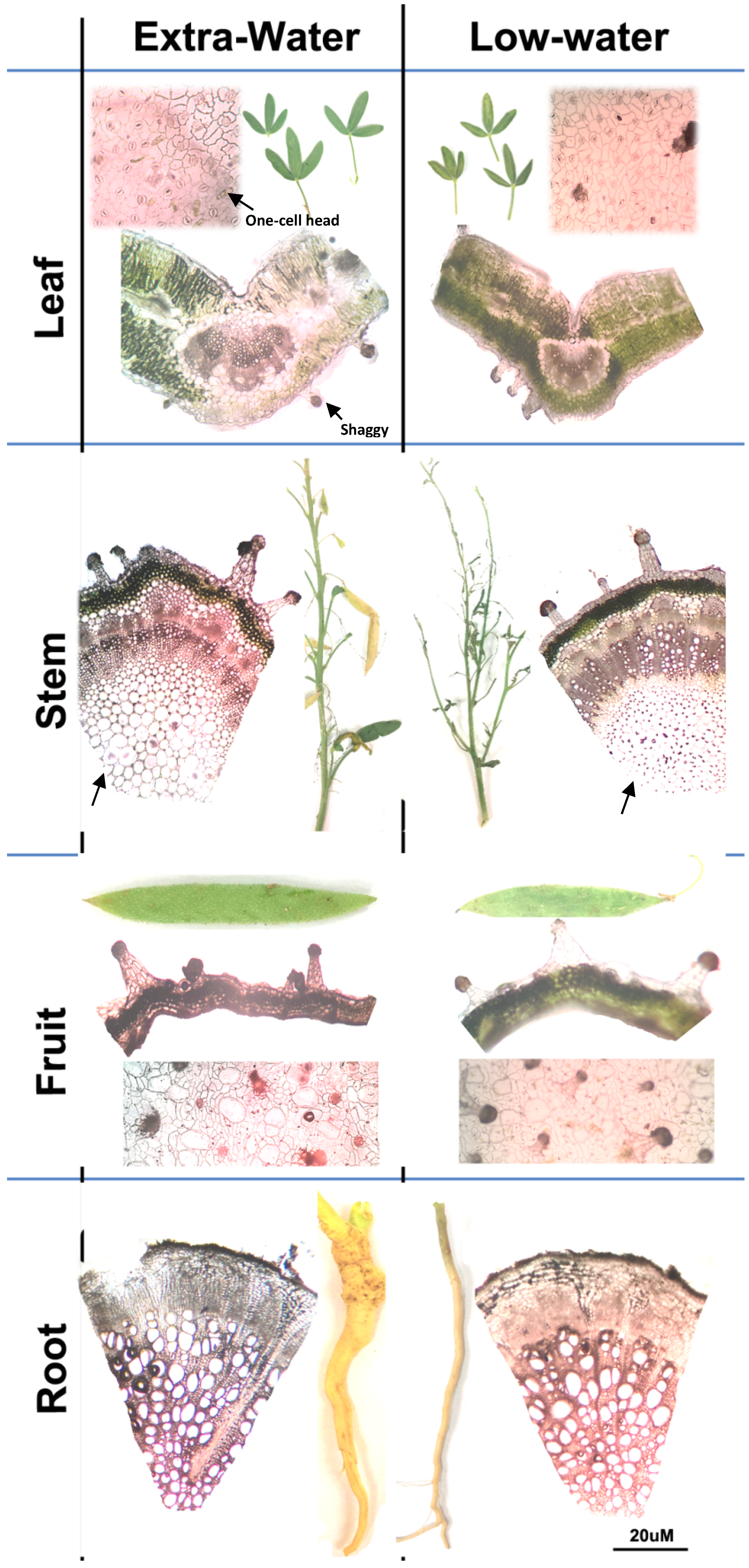
3.1. Anatomical Characteristics of C. amblyocarpa in the Arid Dessert of UAE
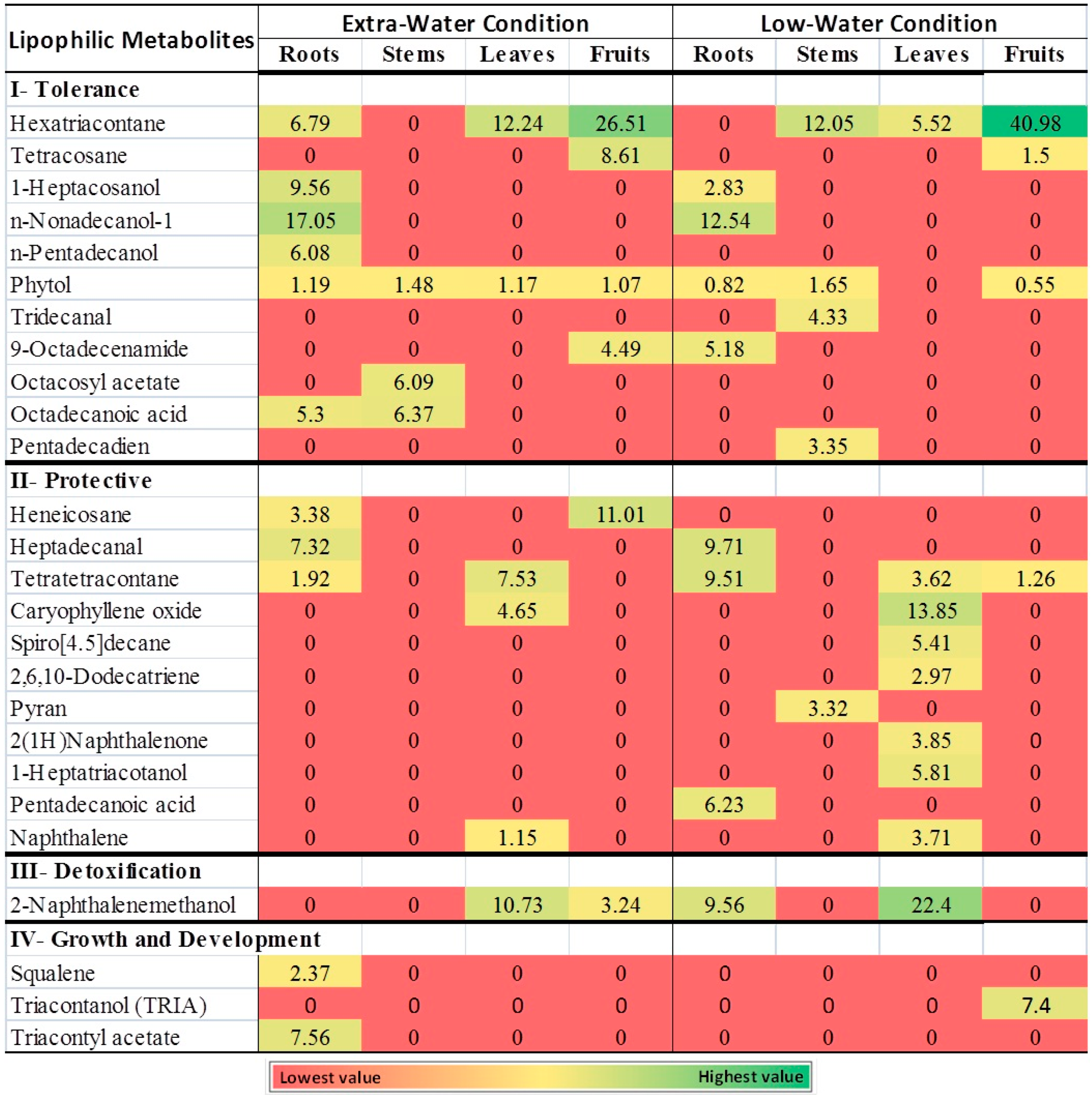
3.2. C. amblyocarpa Growing at the Arid Dessert of UAE Produced Lipophilic Metabolites
3.2.1. Lipophilic Metabolites and Drought Tolerance
3.2.2. Lipophilic Metabolites and Protection from Pathogens Invasion
3.2.3. Lipophilic Metabolites and Detoxification
3.2.4. Lipophilic Metabolites and Growth And Development
3.3. Lipophilic Metabolites Localization: Roots versus Aerial Parts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
References
- Urano, K.; Kurihara, Y.; Seki, M.; Shinozaki, K. ‘Omics’ analyses of regulatory networks in plant abiotic stress responses. Curr. Opin. Plant Biol. 2010, 13, 132–138. [Google Scholar] [CrossRef]
- Zhang, Q.; Cai, M.; Yu, X.; Wang, L.; Guo, C.; Ming, R.; Zhang, J. Transcriptome dynamics of Camellia sinensis in response to continuous salinity and drought stress. Tree Genet. Genomes 2017, 13, 78. [Google Scholar] [CrossRef]
- Bouaziz, M.; Dhouib, A.; Loukil, S.; Boukhris, M.; Sayadi, S. Polyphenols content, antioxidant and antimicrobial activities of extracts of some wild plants collected from the south of Tunisia. Afr. J. Biotechnol. 2009, 8, 7017–7027. [Google Scholar]
- Arbona, V.; Manzi, M.; de Ollas, C.; Gómez-Cadenas, A. Metabolomics as a tool to investigate abiotic stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 4885–4911. [Google Scholar] [CrossRef]
- Wotton, H.R.; Strange, R.N. Increased susceptibility and reduced phytoalexin accumulation in drought-stressed peanut kernels challenged with Aspergillus flavus. Appl. Environ. Microbiol. 1987, 53, 270–273. [Google Scholar] [PubMed]
- Cotty, P.J.; Bayman, P.; Egel, D.S.; Elias, K.S. Agriculture, aflatoxins and Aspergillus. In The Genus Aspergillus: From Taxonomy and Genetics to Industrial Application; Powell, K.A., Renwick, A., Peberdy, J.F., Eds.; Springer: Boston, MA, USA, 1994; pp. 1–27. [Google Scholar] [CrossRef]
- Wiermer, M.; Feys, B.J.; Parker, J.E. Plant immunity: The EDS1 regulatory node. Curr. Opin. Plant Biol. 2005, 8, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Polle, A. Dissecting the superoxide dismutase-ascorbate-glutathione-pathway in chloroplasts by metabolic modeling. Computer simulations as a step towards flux analysis. Plant Physiol. 2001, 126, 445–462. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Shah, J. Lipids, lipases, and lipid-modifying enzymes in plant disease resistance. Annu. Rev. Phytopathol. 2005, 43, 229–260. [Google Scholar] [CrossRef] [PubMed]
- Mueller, M.J. Archetype signals in plants: The phytoprostanes. Curr. Opin. Plant Biol. 2004, 7, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Delude, C.; Moussu, S.; Joubès, J.; Ingram, G.; Domergue, F. Plant surface lipids and epidermis development. In Lipids in Plant and Algae Development. Subcellular Biochemistry; Nakamura, Y., Li-Beisson, Y., Eds.; Springer: Cham, Switzerland, 2016; Volume 86. [Google Scholar]
- Karim, F.; Fawzy, N. Flora of the United Arab Emirates; UAE University Publication: Al-Ain, UAE, 2007. [Google Scholar]
- Jongbloed, M. The Comprehensive Guide to the Wild Flowers of the United Arab Emirates; Environmental Research and Wildlife Development Agency: Abu-Dhabi, UAE, 2003. [Google Scholar]
- Marshall, D.M.; Muhaidat, R.; Brown, N.J.; Liu, Z.; Stanley, S.; Griffiths, H.; Sage, R.F.; Hibberd, J.M. Cleome, a genus closely related to Arabidopsis, contains species spanning a developmental progression from C3 to C4 photosynthesis. Plant J. 2007, 51, 886–896. [Google Scholar] [CrossRef] [PubMed]
- Al-Shanwani, M. Plants Used in Saudi Folk Medicine; King Abdul Aziz City for Scienceand Technology: Riyadh, Saudi Arabia, 1996. [Google Scholar]
- Mossa, J.; Al-Yahya, M.; Al-Meshal, I. Medicinal Plants of Saudi Arabia; King Saud University Press: Riyadh, Saudi Arabia, 2000. [Google Scholar]
- Baba, M.; Shiraiwa, Y. Biosynthesis of Lipids and Hydrocarbons in Algae. In Photosynthesis; Dubinsky, Z., Ed.; IntechOpen: London, UK, 2013. [Google Scholar]
- Boucher, Y.; Kamekura, M.; Doolittle, W.F. Origins and evolution of isoprenoid lipid biosynthesis in archaea. Mol. Microbiol. 2004, 52, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Feulner, G.R. Rainfall and climate records from Sharjah Airport: Historical data for the study of recent climatic periodicity in the UAE. Tribulus 2006, 16, 3–9. [Google Scholar]
- Soliman, S.; Alsaadi, A.; Youssef, E.; Khitrov, G.; Noreddin, A.; Husseiny, M.; Ibrahim, A. Calli essential oils synergize with lawsone against multidrug resistant pathogens. Molecules 2017, 22, 2223. [Google Scholar] [CrossRef] [PubMed]
- Soliman, S.; Mohammad, M.G.; El-Keblawy, A.A.; Omar, H.; Abouleish, M.; Madkour, M.; Elnaggar, A.; Hosni, R.M. Mechanical and phytochemical protection mechanisms of Calligonum comosum in arid deserts. PLoS ONE 2018, 13, e0192576. [Google Scholar] [CrossRef] [PubMed]
- Werker, E. Trichome diversity and development. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2000; Volume 31, pp. 1–35. [Google Scholar]
- Kamel, W.M.; El-Ghani, M.M.A.; El-Bous, M.M. Cleomaceae as a distinct family in the flora of Egypt. Afr. J. Plant Sci. Biotechnol. 2010, 4, 11–16. [Google Scholar]
- Nobel, P. Physicochemical and Environmental Plant Physiology; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Lusa, M.G.; Cardoso, E.C.; Machado, S.R.; Appezzato-da-Glória, B. Trichomes related to an unusual method of water retention and protection of the stem apex in an arid zone perennial species. AoB Plants 2015, 7, plu088. [Google Scholar] [CrossRef]
- Eui, C.Y.; Soon, L.; Hyun-Jung, K.; Yeon, H.J.; Mi-Hyun, L.; Yanyan, Y.; Ji-Ah, K.; Yun-Soo, K. Tobacco NtLTP1, a glandular-specific lipid transfer protein, is required for lipid secretion from glandular trichomes. Plant J. 2012, 70, 480–491. [Google Scholar] [CrossRef]
- David, O.A.; Osonubi, O.; Olaiya, C.O.; Agbolade, J.O.; Ajiboye, A.A.; Komolafe, R.J.; Chukwuma, D.M.; Akomolafe, G.F. Anatomical response of wheat cultivars to drought stress. IFE J. Sci. 2017, 19, 323–331. [Google Scholar] [CrossRef]
- Matthew, H.; Mauro, C.; Alessio, G.; Giovanni, M.; Noemi, P.; Donatella, C.; Anna, D.C.; Francesco, L. Xylem morphology determines the drought response of two Arundo donax ecotypes from contrasting habitats. Gcb Bioenergy 2017, 9, 119–131. [Google Scholar] [CrossRef]
- Malik, W.; Ahmed, D.; Izhar, S. Tyrosinase inhibitory activities of Carissa opaca Stapf ex haines roots extracts and their phytochemical analysis. Pharmacogn. Mag. 2017, 13, 544–548. [Google Scholar] [CrossRef]
- Abdullah, W.; Elsayed, W.M.; Abdelshafeek, K.A.; Nazif, N.M.; Singab, A.N.B. Chemical constituents and biological activities of Cleome genus: A brief review. Int. J. Pharmacogn. Phytochem. Res. 2016, 8, 777–787. [Google Scholar]
- Van Miltenburg, J.C.; Oonk, H.A.J.; Ventola, L. Heat capacities and derived thermodynamic functions of 1-octadecanol, 1-nonadecanol, 1-eicosanol, and 1-docosanol between 10 k and 370 k. J. Chem. Eng. Data 2001, 46, 90–97. [Google Scholar] [CrossRef]
- Buschhaus, C.; Jetter, R. Composition and physiological function of the wax layers coating Arabidopsis leaves: β-amyrin negatively affects the intracuticular Water barrier. Plant Physiol. 2012, 160, 1120–1129. [Google Scholar] [CrossRef]
- Spicher, L.; Almeida, J.; Gutbrod, K.; Pipitone, R.; Dörmann, P.; Glauser, G.; Rossi, M.; Kessler, F. Essential role for phytol kinase and tocopherol in tolerance to combined light and temperature stress in tomato. J. Exp. Bot. 2017, 68, 5845–5856. [Google Scholar] [CrossRef]
- Chimona, C.; Karioti, A.; Skaltsa, H.; Rhizopoulou, S. Occurrence of secondary metabolites in tepals of Asphodelus ramosus L. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2014, 148, 31–34. [Google Scholar] [CrossRef]
- Zhang, D.; Terschak, J.A.; Harley, M.A.; Lin, J.; Hardege, J.D. Simultaneously hermaphroditic shrimp use lipophilic cuticular hydrocarbons as contact sex pheromones. PLoS ONE 2011, 6, e17720. [Google Scholar] [CrossRef]
- Getachew, P.; Getachew, M.; Joo, J.; Choi, Y.S.; Hwang, D.S.; Hong, Y.-K. The slip agents oleamide and erucamide reduce biofouling by marine benthic organisms (diatoms, biofilms and abalones). Toxicol. Environ. Health Sci. 2016, 8, 341–348. [Google Scholar] [CrossRef]
- Seenivasagan, T.; Sharma, K.R.; Sekhar, K.; Ganesan, K.; Prakash, S.; Vijayaraghavan, R. Electroantennogram, flight orientation, and oviposition responses of Aedes aegypti to the oviposition pheromone n-heneicosane. Parasitol. Res. 2009, 104, 827–833. [Google Scholar] [CrossRef]
- Sheoran, N.; Valiya Nadakkakath, A.; Munjal, V.; Kundu, A.; Subaharan, K.; Venugopal, V.; Rajamma, S.; Eapen, S.J.; Kumar, A. Genetic analysis of plant endophytic Pseudomonas putida BP25 and chemo-profiling of its antimicrobial volatile organic compounds. Microbiol. Res. 2015, 173, 66–78. [Google Scholar] [CrossRef]
- Fidyt, K.; Fiedorowicz, A.; Strządała, L.; Szumny, A. β-caryophyllene and β-caryophyllene oxide—Natural compounds of anticancer and analgesic properties. Cancer Med. 2016, 5, 3007–3017. [Google Scholar] [CrossRef] [PubMed]
- Essien, E.E.; Ogunwande, I.A.; Setzer, W.N.; Ekundayo, O. Chemical composition, antimicrobial, and cytotoxicity studies on S. erianthum and S. macranthum essential oils. Pharm. Biol. 2012, 50, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, S.A.; Islam, R.; Islam, R.; Jamal, A.H.M.; Parvin, T.; Rahman, A. Chemical composition and antifungal properties of the essential oil and various extracts of Mikania scandens (L.) Willd. Arab. J. Chem. 2017, 10, S2170–S2174. [Google Scholar] [CrossRef]
- Helms, A.M.; De Moraes, C.M.; Tröger, A.; Alborn, H.T.; Francke, W.; Tooker, J.F.; Mescher, M.C. Identification of an insect-produced olfactory cue that primes plant defenses. Nat. Commun. 2017, 8, 337. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, M.M.; Wang, Q.; Webster, F.X.; Kiemle, D.; Hong, Y.J.; Tantillo, D.J.; Coates, R.M.; Wray, A.T.; Askew, W.; O’Donnell, C.; et al. Formation of the unusual semivolatile diterpene rhizathalene by the Arabidopsis class i terpene synthase tps08 in the root stele is involved in defense against belowground herbivory. Plant Cell 2013, 25, 1108–1125. [Google Scholar] [CrossRef] [PubMed]
- Geen, G.R.; Evans, J.M.; Vong, A.K. Pyrans and Their Benzo Derivatives: Applications; SmithKline Beecham Pharmaceuticals: Harlow, UK, 2008; pp. 469–500. [Google Scholar]
- Amand, S.; Vallet, M.; Guedon, L.; Genta-Jouve, G.; Wien, F.; Mann, S.; Dupont, J.; Prado, S.; Nay, B. A reactive eremophilane and its antibacterial 2(1H)-naphthalenone rearrangement product, witnesses of a microbial chemical warfare. Org. Lett. 2017, 19, 4038–4041. [Google Scholar] [CrossRef] [PubMed]
- Vaithiyanathan, V.; Mirunalini, S. Quantitative variation of bioactive phyto compounds in ethyl acetate and methanol extracts of Pergularia daemia (Forsk.) Chiov. J. Biomed. Res. 2015, 29, 169–172. [Google Scholar] [CrossRef]
- Madhavan, M. Phytochemical constituents of leaves of Spatholobus parviflorus a rare threatened climber of South India. Int. J. Pharmacogn. Phytochem. Res. 2015, 7, 991–994. [Google Scholar]
- Hussein, H.J.; Hadi, M.Y.; Hameed, I.H. Study of chemical composition of Foeniculum vulgare using Fourier transform infrared spectrophotometer and gas chromatography—Mass spectrometry. J. Pharmacogn. Phytother. 2016, 8, 60–89. [Google Scholar] [CrossRef]
- Mansuy, D. Metabolism of xenobiotics: Beneficial and adverse effects. Biol. Aujourd’hui 2013, 207, 33–37. [Google Scholar] [CrossRef]
- Bloch, K.E. Sterol, structure and membrane function. Crit. Rev. Biochem. 1983, 14, 47–92. [Google Scholar] [CrossRef]
- Naeem, M.; Khan, M.M.A.; Moinuddin. Triacontanol: A potent plant growth regulator in agriculture. J. Plant Interact. 2012, 7, 129–142. [Google Scholar] [CrossRef]
- Post-Beittenmiller, D. Biochemistry and molecular biology of wax production in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 405–430. [Google Scholar] [CrossRef]
- Barthlott, W.; Mail, M.; Bhushan, B.; Koch, K. Plant surfaces: Structures and functions for biomimetic innovations. Nano-Micro Lett. 2017, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Singh, A.K.; Khade, P.K. 3-Nitro-2-naphthalenemethanol: A photocleavable protecting group for carboxylic acids. Tetrahedron 2005, 61, 10007–10012. [Google Scholar] [CrossRef]
- Hao, S.; Ma, Y.; Zhao, S.; Ji, Q.; Zhang, K.; Yang, M.; Yao, Y. McWRI1, a transcription factor of the AP2/SHEN family, regulates the biosynthesis of the cuticular waxes on the apple fruit surface under low temperature. PLoS ONE 2017, 12, e0186996. [Google Scholar] [CrossRef]
- Singer, S.D.; Zou, J.; Weselake, R.J. Abiotic factors influence plant storage lipid accumulation and composition. Plant Sci. 2016, 243, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Feakins, S.J.; Kirby, M.E.; Cheetham, M.I.; Ibarra, Y.; Zimmerman, S.R.H. Fluctuation in leaf wax D/H ratio from a southern California lake records significant variability in isotopes in precipitation during the late Holocene. Org. Geochem. 2014, 66, 48–59. [Google Scholar] [CrossRef]
- Feakins, S.J.; Peters, T.; Wu, M.S.; Shenkin, A.; Salinas, N.; Girardin, C.A.J.; Bentley, L.P.; Blonder, B.; Enquist, B.J.; Martin, R.E.; et al. Production of leaf wax n-alkanes across a tropical forest elevation transect. Org. Geochem. 2016, 100, 89–100. [Google Scholar] [CrossRef]
- Mota, A.H.; Silva, C.O.; Nicolai, M.; Baby, A.; Palma, L.; Rijo, P.; Ascensão, L.; Reis, C.P. Design and evaluation of novel topical formulation with olive oil as natural functional active. Pharm. Dev. Technol. 2017, 1–12. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soliman, S.S.M.; Abouleish, M.; Abou-Hashem, M.M.M.; Hamoda, A.M.; El-Keblawy, A.A. Lipophilic Metabolites and Anatomical Acclimatization of Cleome amblyocarpa in the Drought and Extra-Water Areas of the Arid Desert of UAE. Plants 2019, 8, 132. https://doi.org/10.3390/plants8050132
Soliman SSM, Abouleish M, Abou-Hashem MMM, Hamoda AM, El-Keblawy AA. Lipophilic Metabolites and Anatomical Acclimatization of Cleome amblyocarpa in the Drought and Extra-Water Areas of the Arid Desert of UAE. Plants. 2019; 8(5):132. https://doi.org/10.3390/plants8050132
Chicago/Turabian StyleSoliman, Sameh S.M., Mohamed Abouleish, Maged M.M. Abou-Hashem, Alshaimaa M. Hamoda, and Ali A. El-Keblawy. 2019. "Lipophilic Metabolites and Anatomical Acclimatization of Cleome amblyocarpa in the Drought and Extra-Water Areas of the Arid Desert of UAE" Plants 8, no. 5: 132. https://doi.org/10.3390/plants8050132
APA StyleSoliman, S. S. M., Abouleish, M., Abou-Hashem, M. M. M., Hamoda, A. M., & El-Keblawy, A. A. (2019). Lipophilic Metabolites and Anatomical Acclimatization of Cleome amblyocarpa in the Drought and Extra-Water Areas of the Arid Desert of UAE. Plants, 8(5), 132. https://doi.org/10.3390/plants8050132