Common Bean (Phaseolus vulgaris L.) Accumulates Most S-Methylcysteine as Its γ-Glutamyl Dipeptide



Abstract
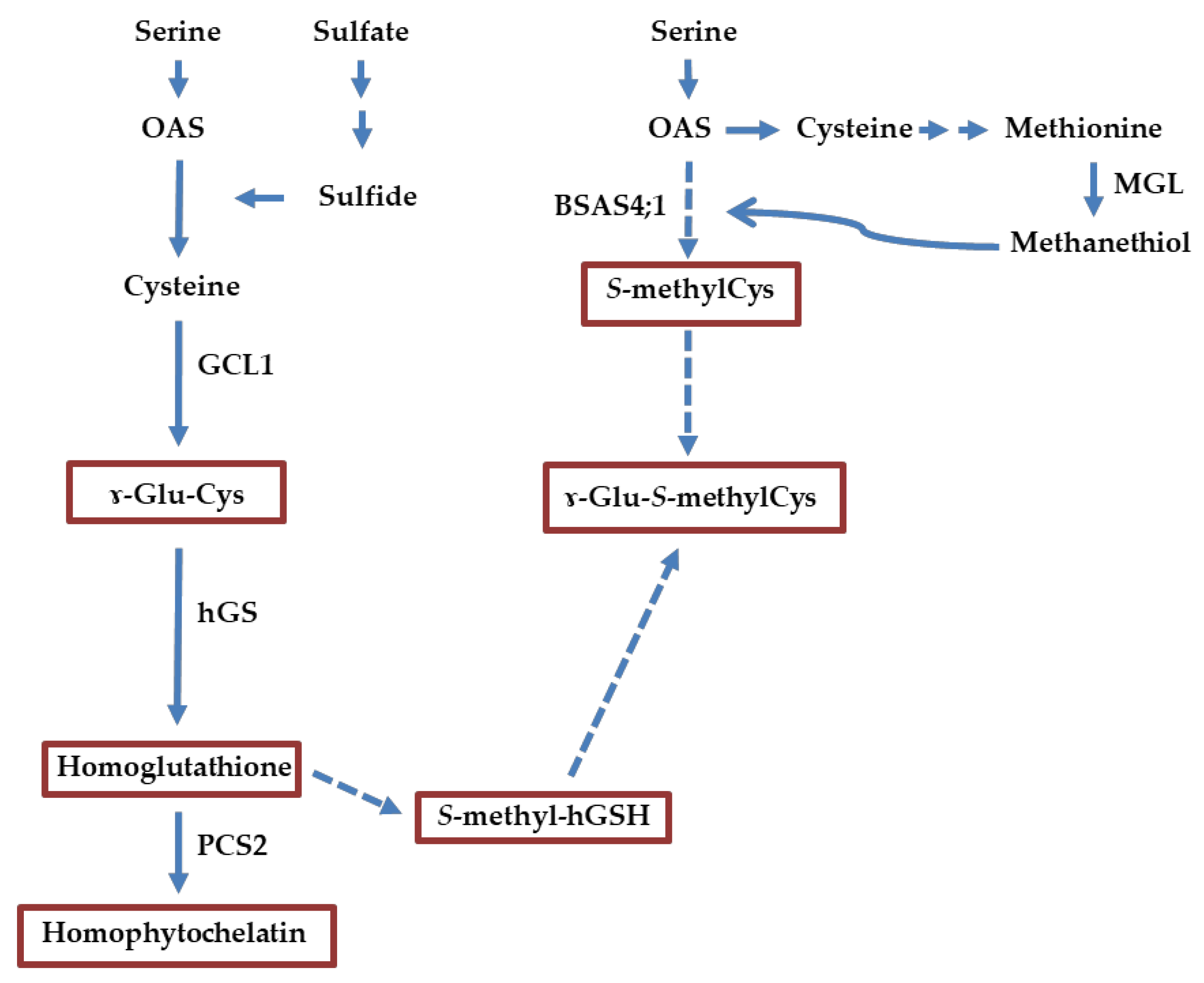
1. Introduction
2. Results
2.1. Analysis of Sulfur Metabolite Profiles During Seed Development
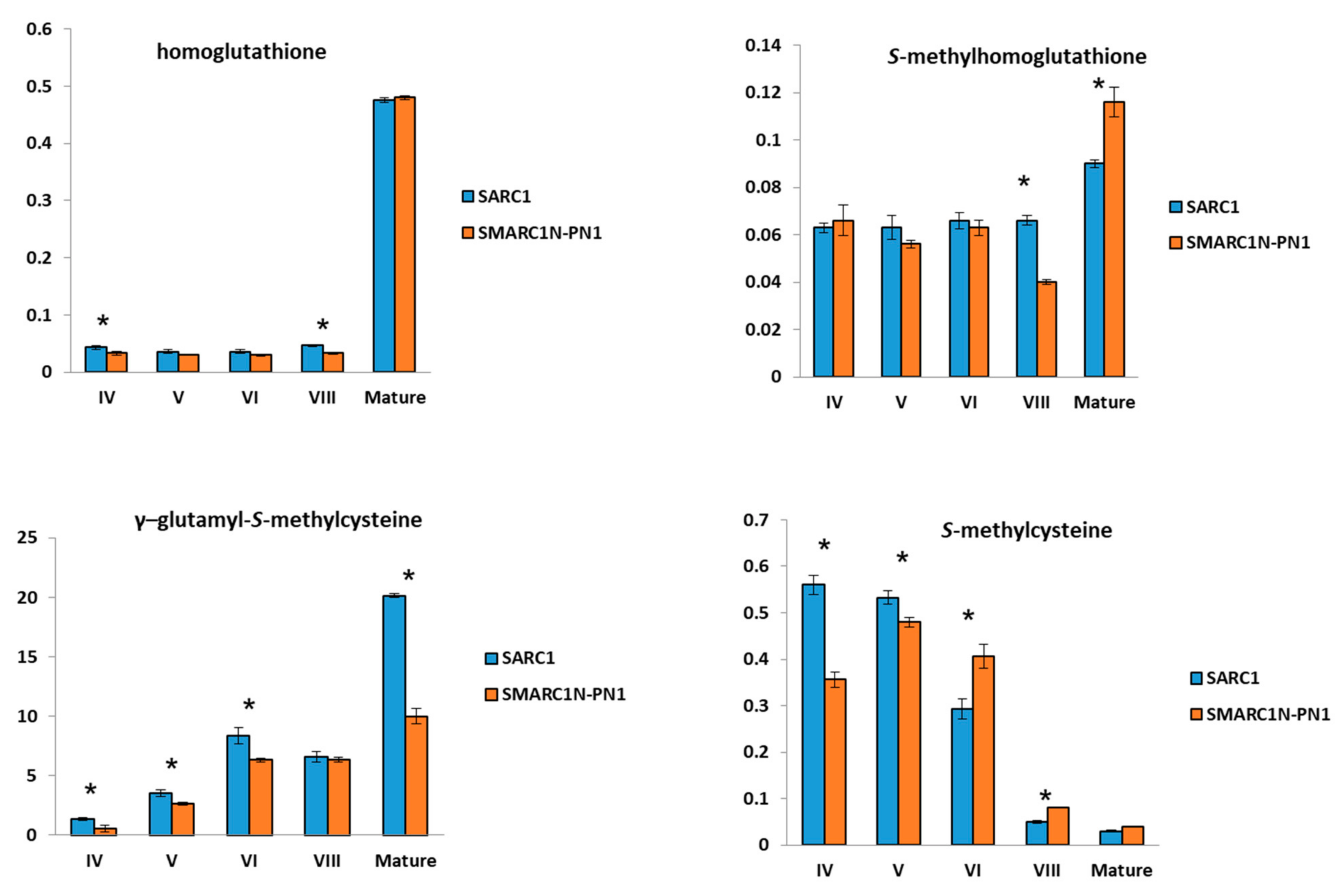
2.2. Differences in Sulfur Amino Acid Concentrations between SARC1 and SMARC1N-PN1 under Sulfate Sufficient Conditions
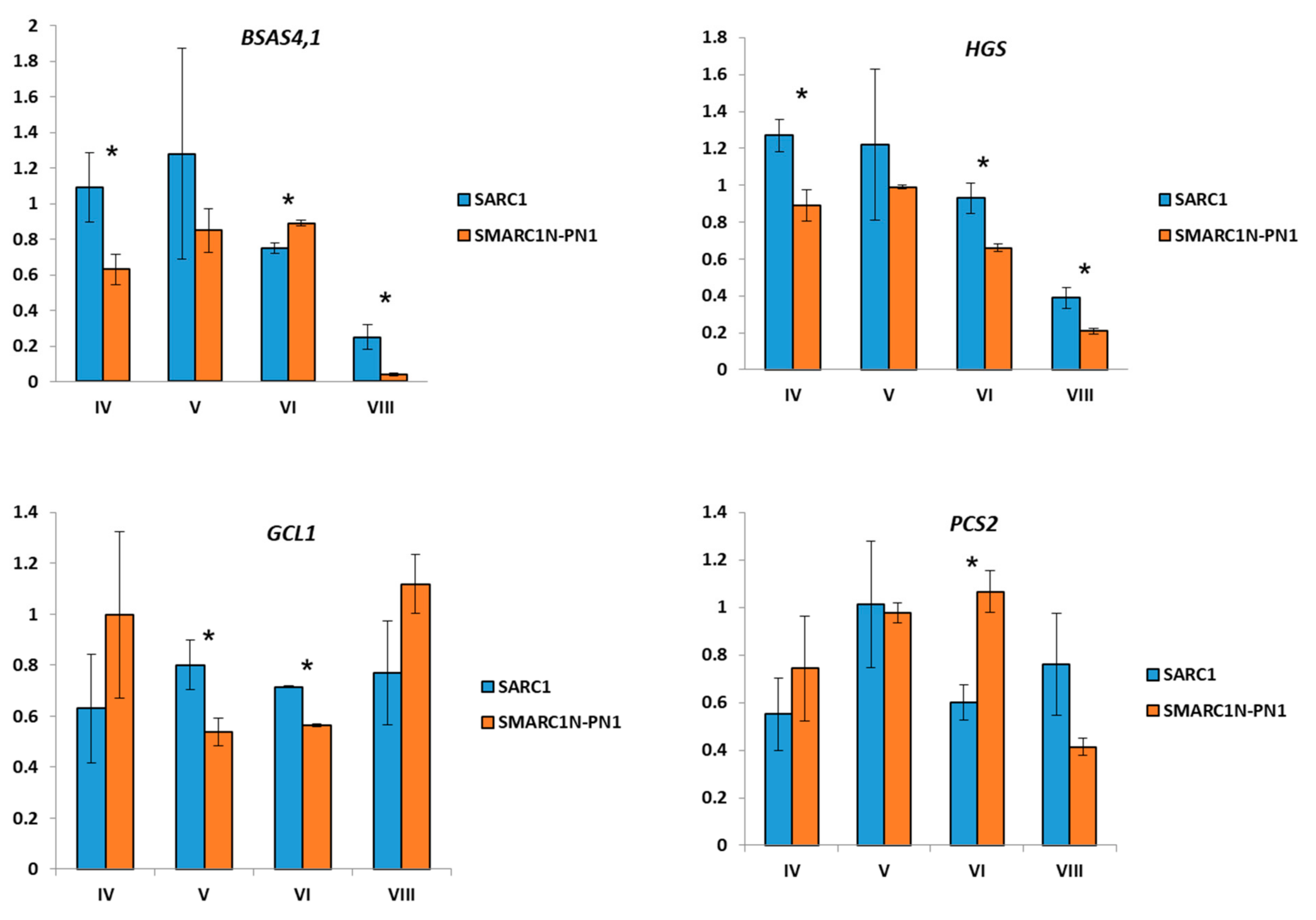
2.3. Expression Analysis of Genes Related to S-Methylcysteine and γ-Glutamyl-S-methylcysteine Biosynthesis
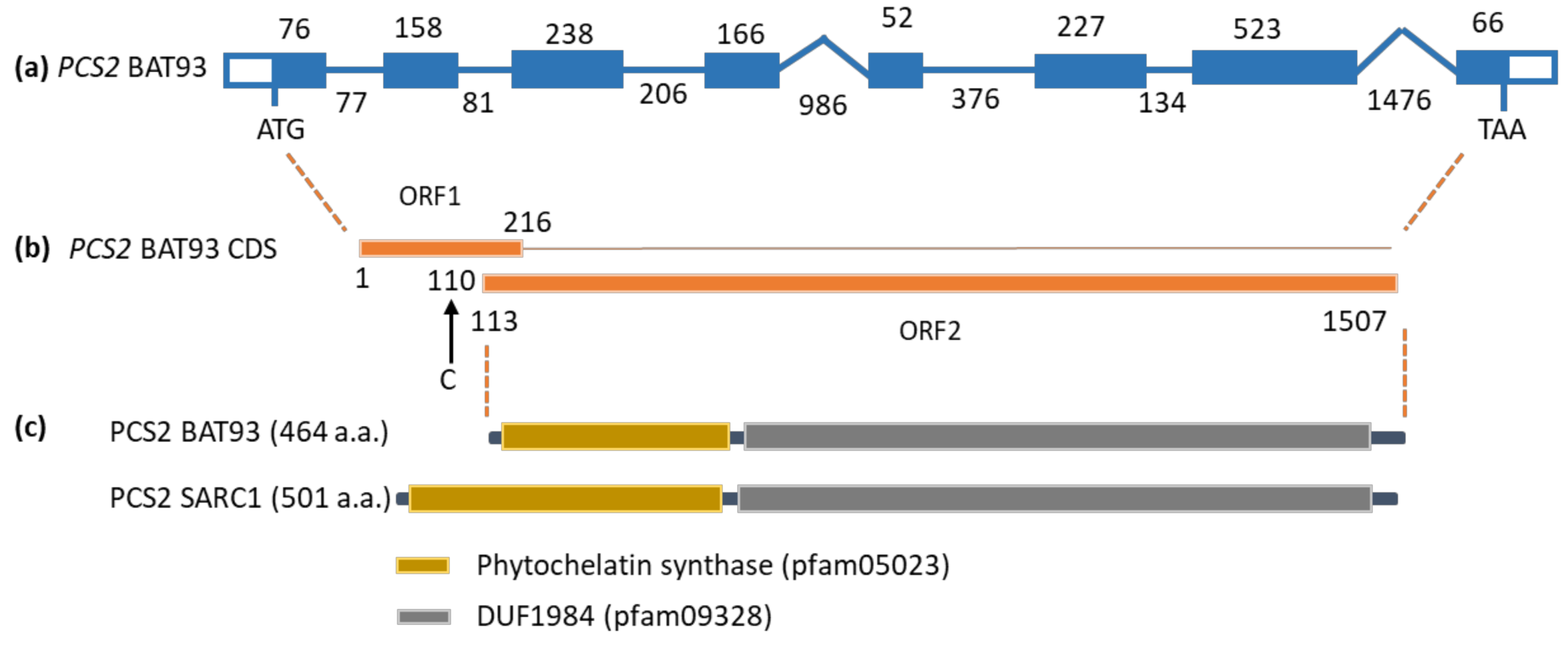
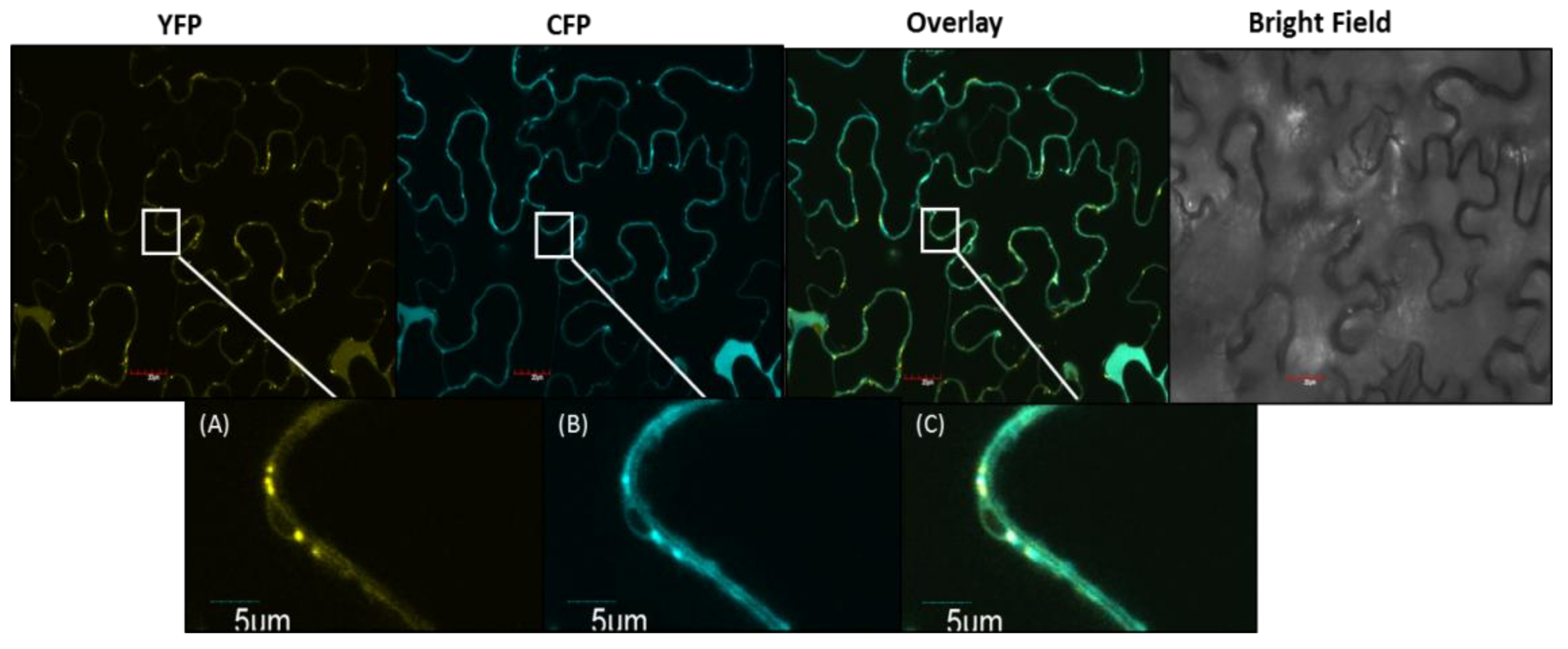
2.4. Subcellular Localization of PCS2
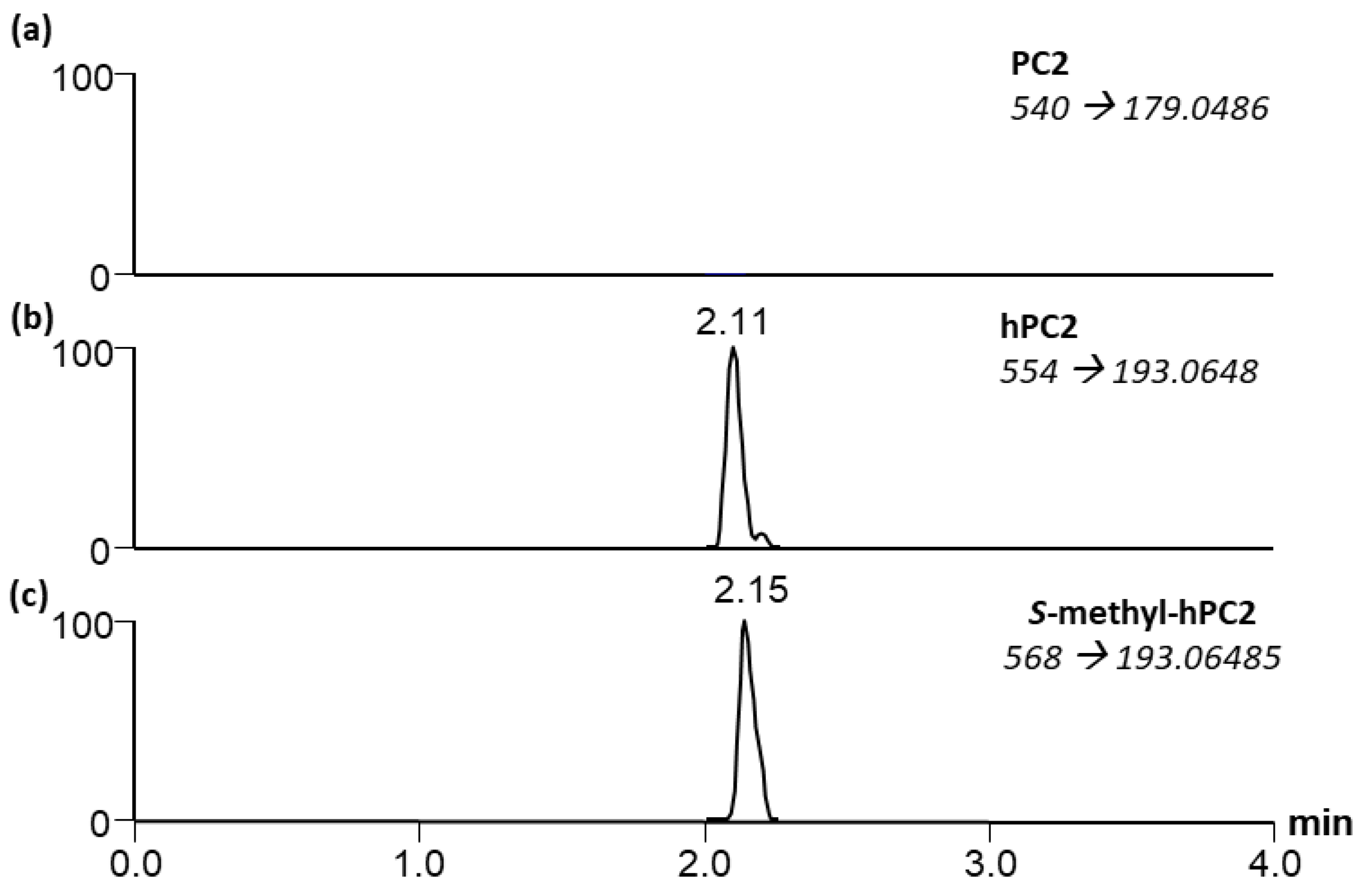
2.5. Analysis of S-Methylated Phytochelatins
3. Discussion
3.1. Most of the S-Methylcysteine Accumulates as γ-Glutamyl-S-methylcysteine in P. vulgaris Seed
3.2. The Concentration of Homoglutathione or S-Methylhomoglutathione does not Appear to Limit the Accumulation of γ-Glutamyl-S-Methylcysteine
3.3. Transcript Expression of BSAS4;1 and hGS is Correlated with the Accumulation of Free S-Methylcysteine and γ-Glutamyl-S-methylcysteine, Respectively
3.4. Mitochondrial Localization of PCS2 Prevents the Accumulation of S-methylated Phytochelatins
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Extraction and Quantification of Free Amino Acids
4.3. RNA Extraction and Quantitative RT-PCR
4.4. Cloning of PCS2 for Subcellular Localization
4.5. Plant Transformation and Confocal Observations
4.6. Analysis of S-methylated Phytochelatins
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Campos-Vega, R.; Reynoso-Camacho, R.; Pedraza-Aboytes, G.; Acosta-Gallegos, J.; Guzman-Maldonado, S.; Paredes-Lopez, O.; Oomah, B.; Loarca-Piña, G. Chemical composition and in vitro polysaccharide fermentation of different beans (Phaseolus vulgaris L.). J. Food Sci. 2009, 74, T59–T65. [Google Scholar] [CrossRef] [PubMed]
- Mojica, L.; de Mejía, E.G. Characterization and comparison of protein and peptide profiles and their biological activities of improved common bean cultivars (Phaseolus vulgaris L.) from Mexico and Brazil. Plant Foods Hum. Nutr. 2015, 70, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Xu, B. Polyphenol-rich dry common beans (Phaseolus vulgaris L.) and their health benefits. Int. J. Mol. Sci. 2017, 18, 2331. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, H.; Liu, R.; Mats, L.; Zhu, H.; Pauls, K.P.; Deng, Z.; Tsao, R. Antioxidant and anti-inflammatory polyphenols and peptides of common bean (Phaseolus vulgaris L.) milk and yogurt in Caco-2 and HT-29 cell models. J. Funct. Foods 2019, 53, 125–135. [Google Scholar] [CrossRef]
- Chen, P.X.; Zhang, H.; Marcone, M.F.; Pauls, K.P.; Liu, R.; Tang, Y.; Zhang, B.; Renaud, J.B.; Tsao, R. Anti-inflammatory effects of phenolic-rich cranberry bean (Phaseolus vulgaris L.) extracts and enhanced cellular antioxidant enzyme activities in Caco-2 cells. J. Funct. Foods 2017, 38, 675–685. [Google Scholar] [CrossRef]
- Nguyen, H.C.; Hoefgen, R.; Hesse, H. Improving the nutritive value of rice seeds: Elevation of cysteine and methionine contents in rice plants by ectopic expression of a bacterial serine acetyltransferase. J. Exp. Bot. 2012, 63, 5991–6001. [Google Scholar] [CrossRef]
- Xiang, X.; Wu, Y.; Planta, J.; Messing, J.; Leustek, T. Overexpression of serine acetyltransferase in maize leaves increases seed-specific methionine-rich zeins. Plant Biotechnol. J. 2017. [Google Scholar] [CrossRef]
- Kim, W.S.; Chronis, D.; Juergens, M.; Schroeder, A.C.; Hyun, S.W.; Jez, J.M.; Krishnan, H.B. Transgenic soybean plants overexpressing O-acetylserine sulfhydrylase accumulate enhanced levels of cysteine and Bowman-Birk protease inhibitor in seeds. Planta 2012, 235, 13–23. [Google Scholar] [CrossRef]
- Galili, G.; Amir, R. Fortifying plants with the essential amino acids lysine and methionine to improve nutritional quality. Plant Biotechnol. J. 2013, 11, 211–222. [Google Scholar] [CrossRef]
- Chiaiese, P.; Ohkama-Ohtsu, N.; Molvig, L.; Godfree, R.; Dove, H.; Hocart, C.; Fujiwara, T.; Higgins, T.J.V.; Tabe, L.M. Sulphur and nitrogen nutrition influence the response of chickpea seeds to an added, transgenic sink for organic sulphur. J. Exp. Bot. 2004, 55, 1889–1901. [Google Scholar] [CrossRef]
- Zhang, Y.; Schernthaner, J.; Labbé, N.; Hefford, M.A.; Zhao, J.; Simmonds, D.H. Improved protein quality in transgenic soybean expressing a de novo synthetic protein, MB-16. Transgenic Res. 2014, 23, 455–467. [Google Scholar] [CrossRef] [PubMed]
- Van, K.; McHale, L.K. Meta-analyses of QTLs associated with protein and oil contents and compositions in soybean [Glycine max (L.) Merr.] seed. Int. J. Mol. Sci. 2017, 18, 1180. [Google Scholar] [CrossRef]
- Kasai, T.; Shiroshita, Y.; Sakamura, S. γ-Glutamyl peptides of Vigna radiata seeds. Phytochemistry 1986, 25, 679–682. [Google Scholar] [CrossRef]
- Giada, M.D.L.R.; Miranda, M.T.M.; Marquez, U.M.L. Sulphur γ-glutamyl peptides in mature seeds of common beans (Phaseolus vulgaris L.). Food Chem. 1998, 61, 177–184. [Google Scholar] [CrossRef]
- Baldi, G.; Salamini, F. Variability of essential amino acid content in seeds of 22 Phaseolus species. Theor. Appl. Genet. 1973, 43, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Li, C.J.; Brownson, D.M.; Mabry, T.J.; Perera, C.; Bell, E.A. Nonprotein amino acids from seeds of Cycas circinalis and Phaseolus vulgaris. Phytochemistry 1996, 42, 443–445. [Google Scholar] [CrossRef]
- Evans, I.M.; Boulter, D. S-Methyl-L-cysteine content of various legume meals. Plant Foods Hum. Nutr. 1975, 24, 257–261. [Google Scholar] [CrossRef]
- Liao, D.; Cram, D.; Sharpe, A.G.; Marsolais, F. Transcriptome profiling identifies candidate genes associated with the accumulation of distinct sulfur γ-glutamyl dipeptides in Phaseolus vulgaris and Vigna mungo seeds. Front. Plant Sci. 2013, 4, 60. [Google Scholar] [CrossRef] [PubMed]
- Osborn, T.; Hartweck, L.; Harmsen, R.; Vogelzang, R.; Kmiecik, K.; Bliss, F. Registration of Phaseolus vulgaris genetic stocks with altered seed protein compositions. Crop Sci. 2003, 43, 1570–1572. [Google Scholar] [CrossRef]
- Taylor, M.; Chapman, R.; Beyaert, R.; Hernández-Sebastià, C.; Marsolais, F. Seed storage protein deficiency improves sulfur amino acid content in common bean (Phaseolus vulgaris L.): Redirection of sulfur from γ-glutamyl-S-methyl-cysteine. J. Agric. Food Chem. 2008, 56, 5647–5654. [Google Scholar] [CrossRef]
- Padovese, R.; Kina, S.M.; Barros, R.M.C.; Borelli, P.; Marquez, U.M.L. Biological importance of γ-glutamyl-S-methylcysteine of kidney bean (Phaseolus vulgaris L.). Food Chem. 2001, 73, 291–297. [Google Scholar] [CrossRef]
- Hawkesford, M.J.; De Kok, L.J. Managing sulphur metabolism in plants. Plant Cell Environ. 2006, 29, 382–395. [Google Scholar] [CrossRef]
- Takahashi, H.; Watanabe-Takahashi, A.; Smith, F.W.; Blake-Kalff, M.; Hawkesford, M.J.; Saito, K. The roles of three functional sulphate transporters involved in uptake and translocation of sulphate in Arabidopsis thaliana. Plant J. 2000, 23, 171–182. [Google Scholar] [CrossRef]
- Hell, R.; Wirtz, M. Molecular biology, biochemistry and cellular physiology of cysteine metabolism in Arabidopsis thaliana. Arabidopsis Book 2011, 9, e0154. [Google Scholar] [CrossRef]
- Smith, F.W.; Ealing, P.M.; Hawkesford, M.J.; Clarkson, D.T. Plant members of a family of sulfate transporters reveal functional subtypes. Proc. Natl. Acad. Sci. USA 1995, 92, 9373–9377. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Kopriva, S.; Giordano, M.; Saito, K.; Hell, R. Sulfur assimilation in photosynthetic organisms: Molecular functions and regulations of transporters and assimilatory enzymes. Annu. Rev. Plant Biol. 2011, 62, 157–184. [Google Scholar] [CrossRef]
- Liao, D.; Pajak, A.; Karcz, S.R.; Chapman, B.P.; Sharpe, A.G.; Austin, R.S.; Datla, R.; Dhaubhadel, S.; Marsolais, F. Transcripts of sulphur metabolic genes are co-ordinately regulated in developing seeds of common bean lacking phaseolin and major lectins. J. Exp. Bot. 2012, 63, 6283–6295. [Google Scholar] [CrossRef] [PubMed]
- Hell, R.; Jost, R.; Berkowitz, O.; Wirtz, M. Molecular and biochemical analysis of the enzymes of cysteine biosynthesis in the plant Arabidopsis thaliana. Amino Acids 2002, 22, 245–257. [Google Scholar] [CrossRef]
- Bonner, E.R.; Cahoon, R.E.; Knapke, S.M.; Jez, J.M. Molecular basis of cysteine biosynthesis in plants: Structural and functional analysis of O-acetylserine sulfhydrylase from Arabidopsis thaliana. J. Biol. Chem. 2005, 280, 38803–38813. [Google Scholar] [CrossRef]
- Hatzfeld, Y.; Maruyama, A.; Schmidt, A.; Noji, M.; Ishizawa, K.; Saito, K. β-Cyanoalanine synthase is a mitochondrial cysteine synthase-like protein in spinach and Arabidopsis. Plant Physiol. 2000, 123, 1163–1172. [Google Scholar] [CrossRef]
- Hell, R.; Bergmann, L. γ-Glutamylcysteine synthetase in higher plants: Catalytic properties and subcellular localization. Planta 1990, 180, 603. [Google Scholar] [CrossRef] [PubMed]
- Hicks, L.M.; Cahoon, R.E.; Bonner, E.R.; Rivard, R.S.; Sheffield, J.; Jez, J.M. Thiol-based regulation of redox-active glutamate-cysteine ligase from Arabidopsis thaliana. Plant Cell 2007, 19, 2653–2661. [Google Scholar] [CrossRef]
- Yadav, S. Heavy metals toxicity in plants: An overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. S. Afr. J. Bot. 2010, 76, 167–179. [Google Scholar] [CrossRef]
- Oven, M.; Raith, K.; Neubert, R.H.; Kutchan, T.M.; Zenk, M.H. Homo-phytochelatins are synthesized in response to cadmium in azuki beans. Plant Physiol. 2001, 126, 1275–1280. [Google Scholar] [CrossRef]
- Yin, F.; Pajak, A.; Chapman, R.; Sharpe, A.; Huang, S.; Marsolais, F. Analysis of common bean expressed sequence tags identifies sulfur metabolic pathways active in seed and sulfur-rich proteins highly expressed in the absence of phaseolin and major lectins. BMC Genomics 2011, 12, 268. [Google Scholar] [CrossRef]
- O’Rourke, J.A.; Iniguez, L.P.; Fu, F.; Bucciarelli, B.; Miller, S.S.; Jackson, S.A.; McClean, P.E.; Li, J.; Dai, X.; Zhao, P.X.; et al. An RNA-Seq based gene expression atlas of the common bean. BMC Genomics 2014, 15, 866. [Google Scholar] [CrossRef] [PubMed]
- Rébeillé, F.; Jabrin, S.; Bligny, R.; Loizeau, K.; Gambonnet, B.; Van Wilder, V.; Douce, R.; Ravanel, S. Methionine catabolism in Arabidopsis cells is initiated by a γ-cleavage process and leads to S-methylcysteine and isoleucine syntheses. Proc. Natl. Acad. Sci. USA 2006, 103, 15687–15692. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, N.; Saito, K. Biosynthesis of S-alk(en)yl-L-cysteine sulfoxides in Allium: Retro perspective. In Sulfur Metabolism in Higher Plants—Fundamental, Environmental and Agricultural Aspects; De Kok, L.J., Hawkesford, M.J., Haneklaus, S.H., Schnug, E., Eds.; Springer International Publishing: Cham, Switzerland, 2017; Volume 49–60. [Google Scholar]
- Watanabe, M.; Mochida, K.; Kato, T.; Tabata, S.; Yoshimoto, N.; Noji, M.; Saito, K. Comparative genomics and reverse genetics analysis reveal indispensable functions of the serine acetyltransferase gene family in Arabidopsis. Plant Cell 2008, 20, 2484–2496. [Google Scholar] [CrossRef]
- Haas, F.H.; Heeg, C.; Queiroz, R.; Bauer, A.; Wirtz, M.; Hell, R. Mitochondrial serine acetyltransferase functions as a pacemaker of cysteine synthesis in plant cells. Plant Physiol. 2008, 148, 1055–1067. [Google Scholar] [CrossRef]
- Walbot, V.; Clutter, M.; Sussex, I.M. Reproductive development and embryogeny in Phaseolus. Phytomorphology 1972, 22, 59–68. [Google Scholar]
- Bobb, A.J.; Eiben, H.G.; Bustos, M.M. PvAlf, an embryo-specific acidic transcriptional activator enhances gene expression from phaseolin and phytohemagglutinin promoters. Plant J. 1995, 8, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Pandurangan, S.; Sandercock, M.; Beyaert, R.; Conn, K.L.; Hou, A.; Marsolais, F. Differential response to sulfur nutrition of two common bean genotypes differing in storage protein composition. Front. Plant Sci. 2015, 6, 92. [Google Scholar] [CrossRef]
- Schmutz, J.; McClean, P.E.; Mamidi, S.; Wu, G.A.; Cannon, S.B.; Grimwood, J.; Jenkins, J.; Shu, S.; Song, Q.; Chavarro, C.; et al. A reference genome for common bean and genome-wide analysis of dual domestications. Nat. Genet. 2014, 46, 707–713. [Google Scholar] [CrossRef]
- Vatamaniuk, O.K.; Mari, S.; Lu, Y.-P.; Rea, P.A. Mechanism of heavy metal ion activation of phytochelatin (PC) synthase: Blocked thiols are sufficient for PC synthase-catalyzed transpeptidation of glutathione and related thiol peptides. J. Biol. Chem. 2000, 275, 31451–31459. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.; Capella-Gutierrez, S.; Rendon-Anaya, M.; Hernandez-Onate, M.; Minoche, A.E.; Erb, I.; Camara, F.; Prieto-Barja, P.; Corvelo, A.; Sanseverino, W.; et al. Genome and transcriptome analysis of the Mesoamerican common bean and the role of gene duplications in establishing tissue and temporal specialization of genes. Genome Biol. 2016, 17, 32. [Google Scholar] [CrossRef]
- Pandurangan, S.; Diapari, M.; Yin, F.; Munholland, S.; Perry, G.; Chapman, B.P.; Huang, S.; Sparvoli, F.; Bollini, R.; Crosby, W.; et al. Genomic analysis of storage protein deficiency in genetically related lines of common bean (Phaseolus vulgaris). Front. Plant Sci. 2016, 7, 389. [Google Scholar] [CrossRef]
- Blum, R.; Meyer, K.C.; Wünschmann, J.; Lendzian, K.J.; Grill, E. Cytosolic action of phytochelatin synthase. Plant Physiol. 2010, 153, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Kuramata, M.; Abe, T.; Takagi, H.; Ozawa, K.; Ishikawa, S. Phytochelatin synthase OsPCS1 plays a crucial role in reducing arsenic levels in rice grains. Plant J. 2017, 91, 840–848. [Google Scholar] [CrossRef]
- Shine, A.M.; Shakya, V.P.; Idnurm, A. Phytochelatin synthase is required for tolerating metal toxicity in a basidiomycete yeast and is a conserved factor involved in metal homeostasis in fungi. Fungal Biol. Biotechnol. 2015, 2, 3. [Google Scholar] [CrossRef]
- Mendoza-Cózatl, D.G.; Jobe, T.O.; Hauser, F.; Schroeder, J.I. Long-distance transport, vacuolar sequestration, tolerance, and transcriptional responses induced by cadmium and arsenic. Curr. Opin. Plant Biol. 2011, 14, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Ruiz, J.M.; Romero, L. Proline metabolism in response to nitrogen toxicity in fruit of French Bean plants (Phaseolus vulgaris L. cv Strike). Sci. Hortic. 2002, 93, 225–233. [Google Scholar] [CrossRef]
- Jafari, M.; Rajabzadeh, A.R.; Tabtabaei, S.; Marsolais, F.; Legge, R.L. Physicochemical characterization of a navy bean (Phaseolus vulgaris) protein fraction produced using a solvent-free method. Food Chem. 2016, 208, 35–41. [Google Scholar] [CrossRef]
- Bruneau, L.; Chapman, R.; Marsolais, F. Co-occurrence of both L-asparaginase subtypes in Arabidopsis: At3g16150 encodes a K+-dependent L-asparaginase. Planta 2006, 224, 668–679. [Google Scholar] [CrossRef]
- Earley, K.W.; Haag, J.R.; Pontes, O.; Opper, K.; Juehne, T.; Song, K.; Pikaard, C.S. Gateway-compatible vectors for plant functional genomics and proteomics. Plant J. 2006, 45, 616–629. [Google Scholar] [CrossRef]
- Nelson, B.K.; Ca, X.; Nebenführ, A. A multicolored set of in vivo organelle markers for co-localization studies in Arabidopsis and other plants. Plant J. 2007, 51, 1126–1136. [Google Scholar] [CrossRef]
- Yu, S.; Bian, Y.; Zhou, R.; Mou, R.; Chen, M.; Cao, Z. Robust method for the analysis of phytochelatins in rice by high-performance liquid chromatography coupled with electrospray tandem mass spectrometry based on polymeric column materials. J. Sep. Sci. 2015, 38, 4146–4152. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Average Seed Weight (mg)/Develop-Mental Stage | 25/IV- Cotyledon | 52/V- Cotyledon | 105/V-Cotyledon | 157/VI- Maturation | 200/VI- Maturation | 348/VIII- Maturation | 160/Mature |
|---|---|---|---|---|---|---|---|
| Asp | 1.6 ± 0.1 | 0.80 ± 0.10 | 0.79 ± 0.02 | 1.2 ± 0.1 | 1.3 ± 0.1 | 1.4 ± 0.2 | 3.4 ± 0.4 |
| Glu | 4.5 ± 0.2 | 4.1 ± 0.2 | 4.0 ± 0.1 | 3.3 ± 0.1 | 2.9 ± 0.1 | 2.8 ± 0.1 | 4.4 ± 0.3 |
| hGSH | 0.06 ± 0.01 | 0.06 ±0.01 | 0.06 ± 0.01 | 0.12 ± 0.01 | 0.12 ± 0.02 | 0.11 ± 0.01 | 0.35 ± 0.06 |
| Ser | 1.3 ± 0.1 | 0.77 ± 0.09 | 0.56 ± 0.01 | 0.50 ± 0.01 | 0.39 ± 0.01 | 0.52 ± 0.12 | 0.24 ± 0.02 |
| His | 1.8 ± 0.1 | 1.3 ± 0.2 | 0.84 ± 0.02 | 0.98 ± 0.07 | 0.11 ± 0.01 | 0. 45 ± 0.29 | 2.5 ± 0.2 |
| γ-Glu-S-methylCys | 2.4 ± 0.1 | 7.0 ± 0.1 | 9.3 ± 0.2 | 10.2 ± 0.2 | 8.6 ± 0.3 | 11.4 ± 2.7 | 38.0 ± 2.0 |
| Gly | 0.33 ± 0.01 | 0.30 ± 0.01 | 0.25 ± 0.01 | 0.25 ± 0.01 | 0.24 ± 0.01 | 0.27 ± 0.03 | 0.58 ± 0.36 |
| Thr | 3.2 ± 0.1 | 3.3 ± 0.2 | 1.8 ± 0.1 | 1.6 ± 0.1 | 0.66 ± 0.02 | 1.3 ± 0.6 | 0.66 ± 0.06 |
| S-methylhGSH | 0.35 ± 0.02 | 0.28 ± 0.03 | 0.18 ± 0.01 | 0.17 ± 0.01 | 0.12 ± 0.01 | 0.13 ± 0.01 | 0.44 ± 0.04 |
| Arg | 5.2 ± 0.4 | 5.6 ± 0.4 | 2.9 ± 0.1 | 4.2 ± 3.2 | 0.41 ± 0.02 | 1.7 ± 1. 1 | 2.4 ± 0.2 |
| Ala | 2.6 ± 0.3 | 1.4 ± 0.1 | 1.3 ± 0. 1 | 0.82 ± 0.04 | 0.50 ± 0.02 | 0.44 ± 0.04 | 1.7± 0.1 |
| γ-Glu-Leu | 0.26 ± 0.05 | 0.39 ± 0.01 | 2.0 ± 0.1 | 1.3 ± 0.1 | 1.9 ± 0.1 | 1.1 ± 0.6 | 1.9 ± 1.4 |
| Tyr | 0.18 ± 0.03 | 0.19 ± 0.02 | 0.17 ± 0.01 | 0.13 ± 0.01 | 0.10 ± 0.01 | 0.08 ± 0.02 | 0.22 ± 0.02 |
| S-methylCys | 1.3 ± 0.1 | 1.3 ± 0. 1 | 0.43 ± 0.01 | 0.28 ± 0.01 | 0.13 ± 0.01 | 0.18 ± 0.05 | 0.32 ± 0.02 |
| Val | 1.8 ± 0.1 | 3.0 ± 0.3 | 0.94 ± 0.02 | 0.90 ± 0.05 | 0.41 ± 0.01 | 0.64 ± 0.21 | 0.93 ± 0.08 |
| Met | 2.9 ± 0.5 | 2.4 ± 0.1 | 0.75 ± 0.02 | 0.55 ± 0.03 | 0.24 ± 0.01 | 0.31 ± 0.07 | 0.38 ± 0.01 |
| Trp | 0.70 ± 0.09 | 0.38 ± 0.11 | 0.95 ± 0.03 | 0.66 ± 0.01 | 0.66 ± 0.01 | 0.51 ± 0.12 | 0.49 ± 0.01 |
| Phe | 0.47 ± 0.01 | 0.39 ± 0.05 | 0.20 ± 0.03 | 0.14 ± 0.01 | 0.09 ± 0.01 | 0.11 ± 0.02 | 0.37 ± 0.03 |
| Ile | 1.9 ± 0.1 | 3.7 ± 0.3 | 1.6 ± 0.1 | 0.90 ± 0.02 | 0.90 ± 0.01 | 0.77 ± 0.11 | 0.28 ± 0.04 |
| Leu | 4.0 ± 2.3 | 1.4 ± 0.5 | 2.2 ± 0.1 | 0.39 ± 0.38 | 0.36 ± 0.01 | 0.42 ± 0.06 | 0.26 ± 0.02 |
| Lys | 0.30 ± 0.02 | 0.59 ± 0.07 | 0.19 ± 0.01 | 0.27 ± 0.02 | 0.14 ± 0.07 | 0.19 ± 0.02 | 0.35 ± 0.03 |
| Gene | Accession Number | Forward Primer Sequence (5́–3́) | Reverse Primer Sequence (5́–3́) |
|---|---|---|---|
| BSAS4;1 | Phvul.007G185200.2 | GCGGCTGATGGTGGTTATATTT | CACCAGTTCCTATCCCTGCA |
| GCL1 | Phvul.002G289200.1 | TGCATTACCAGCACTTTGGG | ACATCTGTCTTTCTTCTGGGGT |
| hGS | Phvul.006G094500.1 | CCGCAAAGAGAAGGAGGAGG | TGCTGGAAAAGTGGCTGGAA |
| PCS2 | Phvul.001G162700.1 | TTCTCTGGGAGGCAATGAGC | ACCTTCATCTCTACAACTCACAGT |
| Ubiquitin | Phvul.007G05260.1 | ACAGCTGGAGGATGAAAGGA | GTCCGAACTCTCCACCTCAA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saboori-Robat, E.; Joshi, J.; Pajak, A.; Solouki, M.; Mohsenpour, M.; Renaud, J.; Marsolais, F. Common Bean (Phaseolus vulgaris L.) Accumulates Most S-Methylcysteine as Its γ-Glutamyl Dipeptide. Plants 2019, 8, 126. https://doi.org/10.3390/plants8050126
Saboori-Robat E, Joshi J, Pajak A, Solouki M, Mohsenpour M, Renaud J, Marsolais F. Common Bean (Phaseolus vulgaris L.) Accumulates Most S-Methylcysteine as Its γ-Glutamyl Dipeptide. Plants. 2019; 8(5):126. https://doi.org/10.3390/plants8050126
Chicago/Turabian StyleSaboori-Robat, Elham, Jaya Joshi, Aga Pajak, Mahmood Solouki, Motahhareh Mohsenpour, Justin Renaud, and Frédéric Marsolais. 2019. "Common Bean (Phaseolus vulgaris L.) Accumulates Most S-Methylcysteine as Its γ-Glutamyl Dipeptide" Plants 8, no. 5: 126. https://doi.org/10.3390/plants8050126
APA StyleSaboori-Robat, E., Joshi, J., Pajak, A., Solouki, M., Mohsenpour, M., Renaud, J., & Marsolais, F. (2019). Common Bean (Phaseolus vulgaris L.) Accumulates Most S-Methylcysteine as Its γ-Glutamyl Dipeptide. Plants, 8(5), 126. https://doi.org/10.3390/plants8050126