Rice Novel Semidwarfing Gene d60 Can Be as Effective as Green Revolution Gene sd1



Abstract
1. Introduction
2. Results
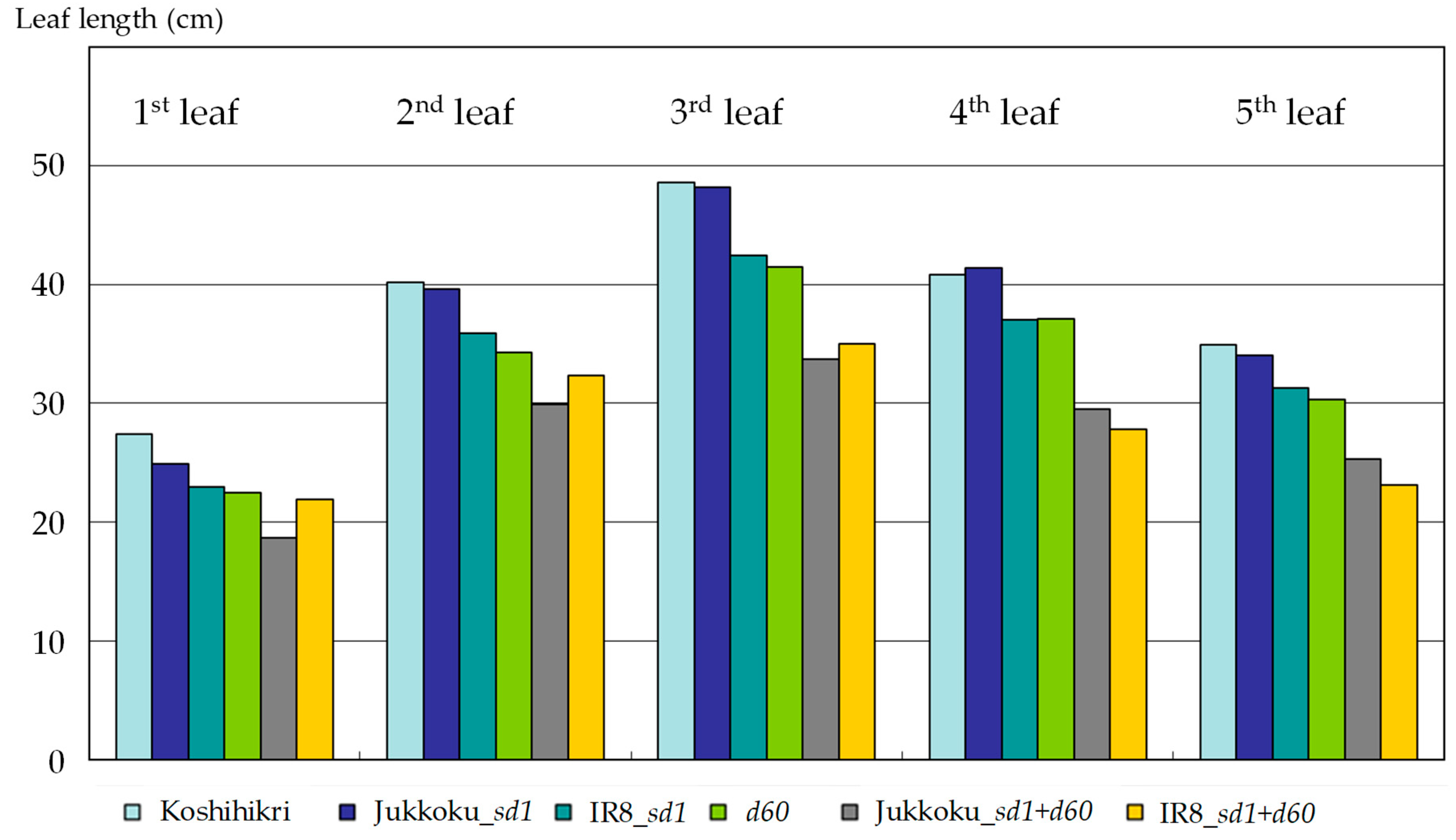
2.1. Effects of Semidwarf and Double Dwarf Genes on Growth
2.2. Effects of Semidwarf and Double Dwarf Genes on Yield
3. Discussion
4. Materials and Methods
4.1. Genetic Lines
4.2. Cultivation
4.3. Growth Analysis
4.4. Plant Phenotyping
4.5. Yield
- Yield of unpolished rice (g/m2) = (number of panicles/m2) × (floret number/panicle) × (proportion of fertile florets) × (weight of unpolished rice/grain)
- Yield = (yield index) × (weight of the plant parts above the ground)
Author Contributions
Funding
Conflicts of Interest
References
- Khush, G.S. Green revolution: Preparing for the 21st century. Genome 1999, 42, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Hergrove, T.; Coffman, W.R. Breeding history. In Rice that Changed the World: Cerebrating 50 Years of IR8; Special supplement focusing on IR8; International Rice Research Institute: Los Baños, Philippines, 2016; pp. 6–10. Available online: http://books.irri.org/RT_Supplement-IR8.pdf (accessed on 25 September 2019).
- Jennings, P. Rice revolutions in Latin America. In Rice that Changed the World: Cerebrating 50 Years of IR8; Special supplement focusing on IR8; International Rice Research Institute: Los Baños, Philippines, 2016; p. 19. Available online: http://books.irri.org/RT_Supplement-IR8.pdf (accessed on 25 September 2019).
- Okada, M.; Yamakawa, Y.; Fujii, K.; Nishiyama, H.; Motomura, H.; Kai, S.; Imai, T. On the new varieties of paddy rice, ‘Hoyoku’, ‘Kokumasari’ and ‘Shiranui’. J. Agric. Exp. Stn. Kyushu 1967, 2, 187–224. [Google Scholar]
- Futuhara, Y. Breeding of new rice variety Reimei by gamma ray irradiation. Gamma. Field Symp. 1968, 7, 87–109. [Google Scholar]
- Rutger, J.N.; Peterson, M.L.; Hu, C.H.; Lehman, W.F. Induction of usefil short stature and early maturing mutants in two japonica rice cultivars. Crop Sci. 1976, 16, 631–635. [Google Scholar] [CrossRef]
- Rutger, J.N.; Peterson, M.L.; Hu, C.H. Registration of Calrose76 rice. Crop Sci. 1977, 17, 978. [Google Scholar] [CrossRef]
- Aquino, R.C.; Jennings, P.R. Inheritance and significance of dwarfism in an indica rice variety. Crop Sci. 1966, 6, 551–554. [Google Scholar] [CrossRef]
- Heu, M.; Chang, T.T.; Cabanilla, V.L. The inheritance of culm length, panicle length, duration to heading and bacterial leaf blight reaction in rice cross: Sigadis×Taichung (Native)1. Jpn. J. Breed. 1968, 18, 7–11. [Google Scholar]
- Foster, K.W.; Rutger, J.N. Inheritance of semidwarfism in rice, Oryza sativa L. Genetics 1978, 88, 559–574. [Google Scholar]
- Mackill, D.J.; Rutger, J.N. The inheritance of induced-mutant semidwarfing genes bin rice. J. Hered. 1979, 70, 335–341. [Google Scholar] [CrossRef]
- Ikehashi, H.; Kikuchi, F. Genetic analysis of semidwarfness and their significance for breeding of high-yielding varieties in rice. JARQ 1982, 15, 231–235. [Google Scholar]
- Kikuchi, F.; Itakura, N.; Ikehashi, H.; Yokoo, M.; Nakane, A.; Maruyama, K. Genetic analysis of semidwarfism in high-yielding rice varieties in Japan. Bull. Natl. Inst. Agr. Sci. Ser. D 1985, 36, 125–145. [Google Scholar]
- Kikuchi, F.; Futsuhara, Y. Inheritance of morphological characters. 2. Inheritance of semidwarf. In Science of the Rice Plant, 3rd ed.; Matsuo, T., Shimizu, S., Tsunoda, S., Murata, Y., Kumazawa, K., Futsuhara, Y., Hoshikawa, K., Yamaguchi, H., Kikuchi, F., Eds.; Tokyo Food and Agricultural Policy Research Center: Tokyo, Japan, 1997; Volume 3, pp. 309–317. [Google Scholar]
- Tomita, M. Combining two semidwarfing genes d60 and sd1 for reduced height in ‘Minihikari’, a new rice germplasm in the ‘Koshihikari’ genetic background. Genet. Res. Camb. 2012, 94, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Suh, H.S.; Heu, M.H. The segregation mode of plant height in the cross of rice varieties. Ⅵ. Linkage analysis of the semi-dwarfness of the rice variety Tongil. Korean J. Breed. 1978, 10, 1–6. [Google Scholar]
- Maeda, H.; Ishii, T.; Mori, H.; Kuroda, J.; Horimoto, M.; Takamure, I.; Kinoshita, T.; Kamijima, O. High density molecular map of semidwarfing gene, sd-1, in rice (Oryza sativa L.). Breed. Sci. 1997, 47, 317–320. [Google Scholar] [CrossRef]
- Sasaki, A.; Ashikari, M.; Ueguchi-Tanaka, M.; Itoh, H.; Nishimura, A.; Swapan, D.; Ishiyama, K.; Saito, T.; Kobayashi, M.; Khush, G.S.; et al. Green revolution: A mutant gibberellin-synthesis gene in rice. Nature 2002, 416, 701–702. [Google Scholar] [CrossRef] [PubMed]
- Monna, L.; Kitazawa, N.; Yoshino, R.; Suzuki, J.; Masuda, H.; Maehara, Y.; Tanji, M.; Sato, M.; Nasu, S.; Minobe, Y. Positional cloning of rice semidwarfing gene, sd-1: Rice “Green revolution gene” encodes a mutant enzyme involved in gibberellin synthesis. DNA Res. 2002, 9, 11–17. [Google Scholar] [CrossRef]
- Spielmeyer, W.; Ellis, M.H.; Chandler, P.M. Semidwarf (sd-1), “green revolution” rice, contains a defective gibberellin 20-oxidase gene. Proc. Natl. Acad. Sci. USA 2002, 99, 9043–9048. [Google Scholar] [CrossRef]
- Ogi, Y.; Kato, H.; Maruyama, K.; Kikuchi, F. The effects of the culm length andother agronomic characters cause by semidwarfing genes at the sd-1 locus inrice. Jpn. J. Breed. 1993, 43, 267–275. [Google Scholar] [CrossRef]
- Murai, M.; Komazaki, T.; Sato, S. Effects of sd1 and Ur1 (Undulate rachis-1) on lodging resistance and related traits in rice. Breed. Sci. 2004, 54, 333–340. [Google Scholar] [CrossRef][Green Version]
- Tomita, M.; Matsumoto, S. Transcription of rice Green Revolution sd1 gene is clarified by comparative RNA diagnosis using the isogenic background. Genom. Appl. Biol. 2011, 2, 29–35. [Google Scholar]
- Tomita, M. Introgression of Green Revolution sd1 gene into isogenic genome of rice super cultivar Koshihikari to create novel semidwarf cultivar ‘Hikarishinseiki’ (Koshihikari-sd1). Field Crops Res. 2009, 114, 173–181. [Google Scholar] [CrossRef]
- Tabuchi, H.; Hashimoto, N.; Takeuchi, A.; Terao, T.; Fukuta, Y. Genetic analysis of semidwarfism of the japonica rice cultivar Kinuhikari. Breed. Sci. 2000, 50, 1–7. [Google Scholar] [CrossRef]
- Tomita, M.; Ishii, K. Genetic performance of the semidwarfing allele sd1 derived from a Japonica rice cultivar and minimum requirements to detect its single-nucleotide polymorphism by MiSeq whole-genome sequencing. BioMed Res. Int. 2018, 2018, 4241725. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Tomita, M. Identification of an isogenic semidwarf rice cultivar carrying the Green Revolution sd1 gene using multiplex codominant ASP-PCR and SSR markers. Biochem. Genet. 2013, 51, 530–542. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days after Sowing | 35 | 43 | 49 | 55 | 64 | 70 | 78 | 82 | 90 | 96 | 103 |
| Koshihikari | 212.6 | 345.0 | 415.4 | 452.4 | 529.5 | 657.0 | 805.6 | 852.6 | 913.1 | 1011.6 | 1020.3 |
| Jukkoku_sd1 | 166.8 * | 330.2 | 408.9 | 430.7 | 523.8 | 631.0 * | 743.0 * | 791.7 * | 828.7 * | 865.9 * | 880.8 * |
| IR8_sd1 | 170.3 * | 308.9 * | 393.2 | 414.9 * | 488.8 * | 578.2 * | 693.2 * | 734.4 * | 794.6 * | 837.9 * | 831.0 * |
| d60 | 189.1 * | 321.0 * | 406.5 | 415.9 * | 480.5 * | 550.4 * | 656.8 * | 689.3 * | 744.1 * | 810.6 * | 814.3 * |
| Jukkoku_sd1+d60 | 153.1 * | 275.4 * | 352.8 * | 357.8 * | 364.1 * | 409.2 * | 506.4 * | 543.4 * | 581.9 * | 616.6 * | 651.7 * |
| IR8_sd1+d60 | 153.7 * | 279.2 * | 364.8 * | 368.5 * | 369.1 * | 454.5 * | 558.4 * | 580.9 * | 630.9 * | 679.2 * | 684.2 * |
| Koshihikari | Jukkoku_sd1 | IR8_sd1 | d60 | Jukkoku _sd1+d60 | IR8_sd1+d60 | |
|---|---|---|---|---|---|---|
| Weight of unpolished rice/1000 grains (g) | 22.1 | 20.2 | 20.4 | 20.9 | 21.3 | 20.5 |
| Panicles /m2 | 397.4 | 461.8 * | 391.8 | 397.4 | 341.9 * | 399.6 |
| Floret number/panicle | 78.6 | 87.3 * | 82.8 | 79.3 | 72.1 * | 76.2 |
| Seed fertility (%) | 96.6 | 94.9 | 93.4 | 95.4 | 95.6 | 96.2 |
| Yield of unpolished rice (g/m2=kg/a) | 665.3 | 772.6 * | 617.9 * | 628.5 | 502.1 * | 600.5 * |
| Koshihikari | Jukkoku_sd1 | IR8_sd1 | d60 | Jukkoku _sd1+d60 | IR8_sd1+d60 | |
|---|---|---|---|---|---|---|
| No. of panicles/plant | 17.9 | 20.8 * | 17.7 | 17.9 | 15.4 * | 18.0 |
| Percent change (%) | - | +16.2 | −1.4 | ±0 | −14.0 | +0.6 |
| Koshihikari | Jukkoku_sd1 | IR8_sd1 | d60 | Jukkoku _sd1+d60 | IR8_sd1+d60 | |
|---|---|---|---|---|---|---|
| No. of Florets/panicle | 78.6 | 87.3 * | 82.8 | 79.3 | 72.1 * | 76.1 |
| Percent change (%) | - | +11.1 | +5.3 | +0.9 | −8.3 | −3.1 |
| Koshihikari | Jukkoku_sd1 | IR8_sd1 | d60 | Jukkoku_sd1+d60 | IR8_sd1+d60 | |
|---|---|---|---|---|---|---|
| Floret number/panicle | 78.6 | 87.3 * | 82.8 | 79.3 | 72.1 * | 76.1 |
| Panicle length (cm) | 16.2 | 16.6 | 15.8 | 15.7 | 15.8 | 15.7 |
| Floret density (/cm) | 4.85 | 5.26 * | 5.23 * | 5.05 | 4.55 * | 4.86 |
| Percent change of floret density (%) | - | +8.53 | +7.95 | +4.11 | −6.15 | +0.32 |
| Koshihikari | Jukkoku_sd1 | IR8_sd1 | d60 | Jukkoku _sd1+d60 | IR8_sd1+d60 | |
|---|---|---|---|---|---|---|
| Yield of unpolished rice (g/m2=kg/a) | 665.3 | 772.6 * | 617.9 * | 628.5 | 502.1 * | 600.5 * |
| Percent change (%) | - | +16.1 | −7.1 | −5.5 | −24.5 | −9.7 |
| Koshihikari | Jukkoku_sd1 | IR8_sd1 | d60 | Jukkoku _sd1+d60 | IR8_sd1+d60 | |
|---|---|---|---|---|---|---|
| Weight of winnowed paddy (g) | 26.9 | 27.3 | 26.4 | 22.7 * | 20.3 * | 23.9 * |
| Weight of the plant part above the ground (g) | 72.7 | 72.8 | 61.5 * | 58.9 * | 44.6 * | 51.4 * |
| Yield index (%) | 37.0 | 37.5 | 42.8 * | 38.6 | 45.6 * | 46.5 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomita, M.; Ishimoto, K. Rice Novel Semidwarfing Gene d60 Can Be as Effective as Green Revolution Gene sd1. Plants 2019, 8, 464. https://doi.org/10.3390/plants8110464
Tomita M, Ishimoto K. Rice Novel Semidwarfing Gene d60 Can Be as Effective as Green Revolution Gene sd1. Plants. 2019; 8(11):464. https://doi.org/10.3390/plants8110464
Chicago/Turabian StyleTomita, Motonori, and Keiichiro Ishimoto. 2019. "Rice Novel Semidwarfing Gene d60 Can Be as Effective as Green Revolution Gene sd1" Plants 8, no. 11: 464. https://doi.org/10.3390/plants8110464
APA StyleTomita, M., & Ishimoto, K. (2019). Rice Novel Semidwarfing Gene d60 Can Be as Effective as Green Revolution Gene sd1. Plants, 8(11), 464. https://doi.org/10.3390/plants8110464