HWA1- and HWA2-Mediated Hybrid Weakness in Rice Involves Cell Death, Reactive Oxygen Species Accumulation, and Disease Resistance-Related Gene Upregulation


 ,
,
Abstract
1. Introduction
2. Results
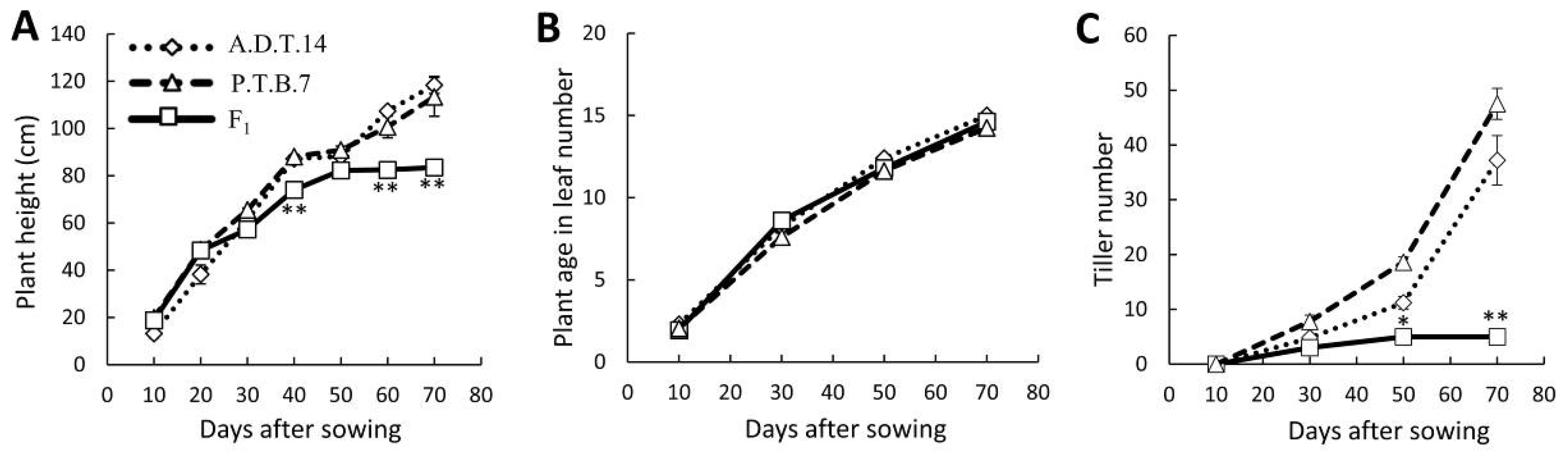
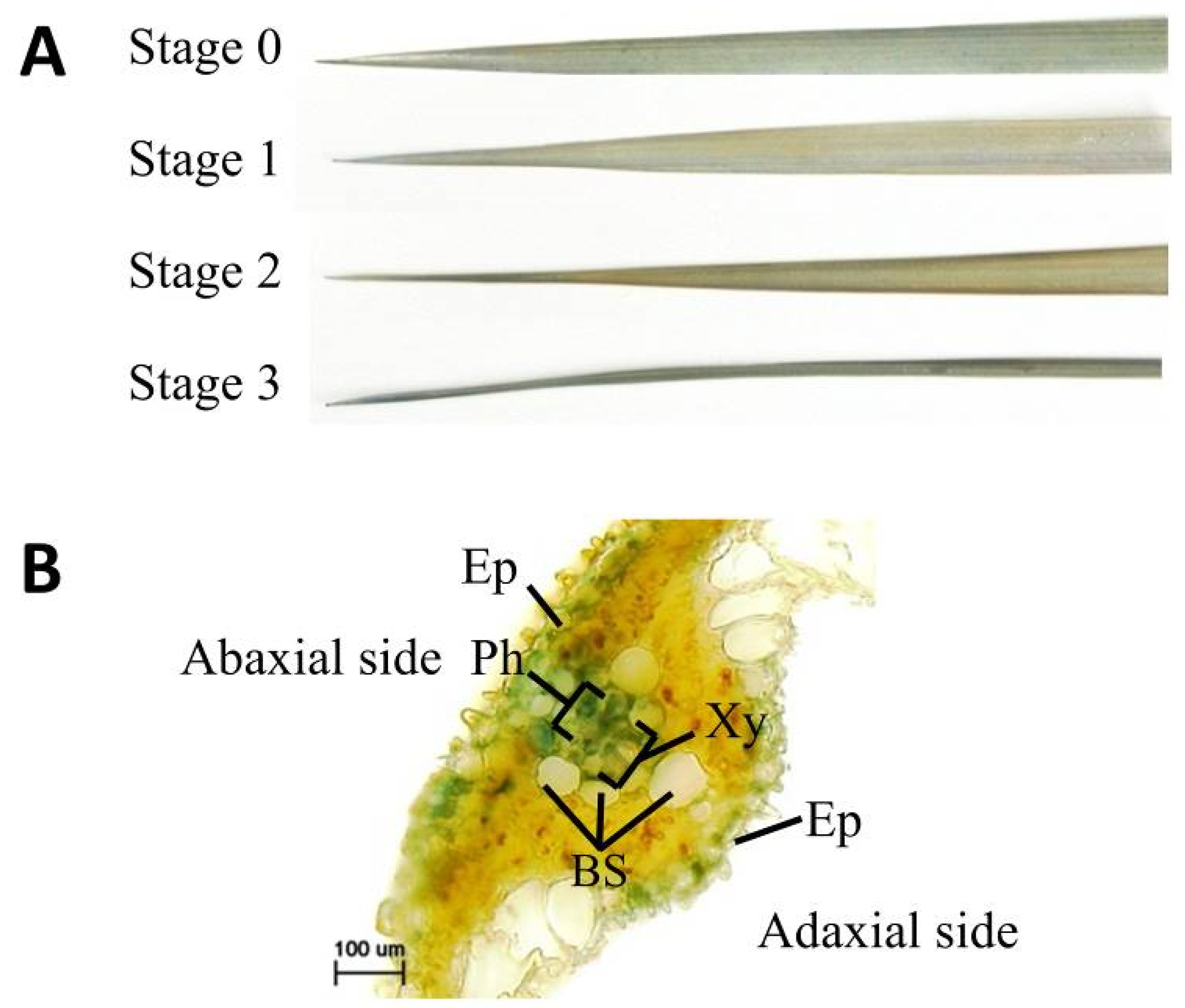
2.1. Hybrid Weakness Phenotypes
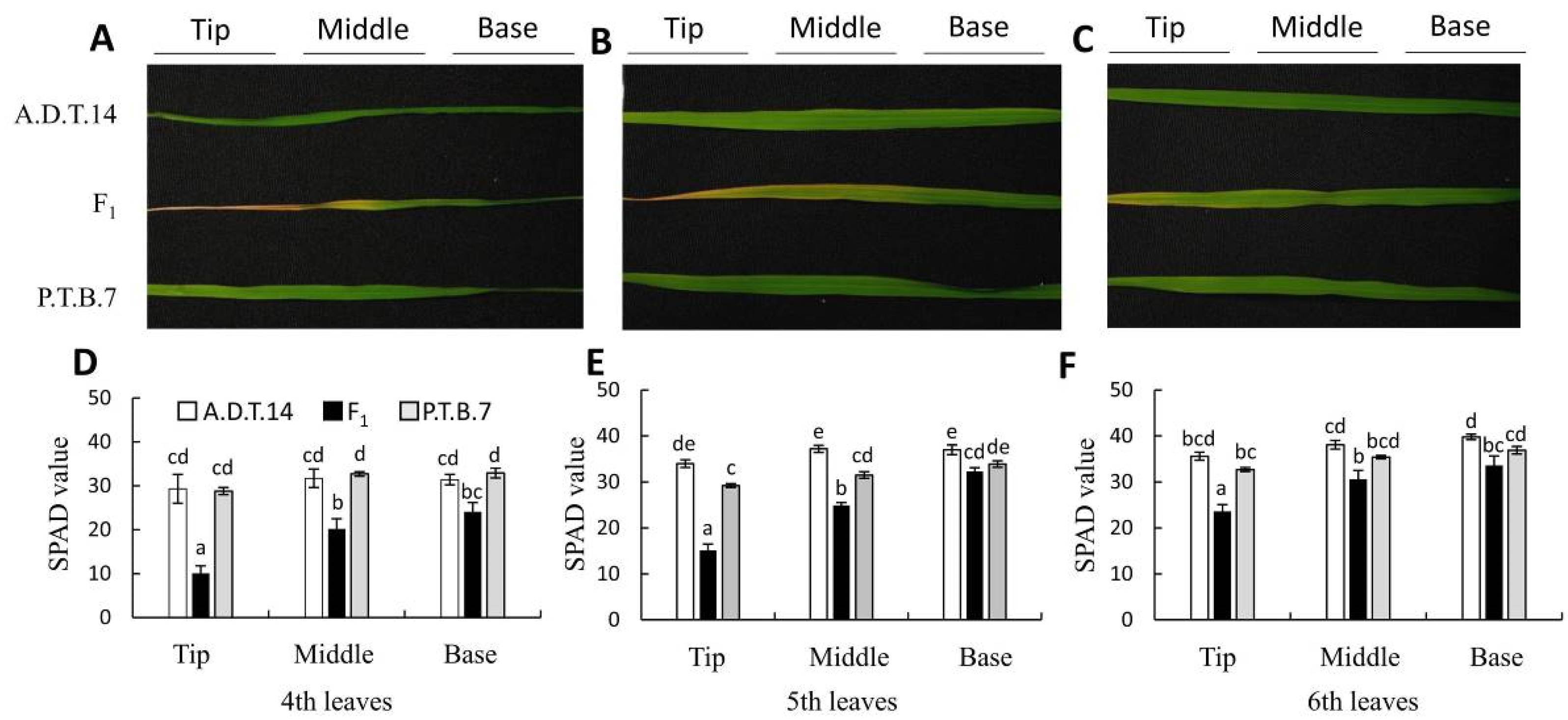
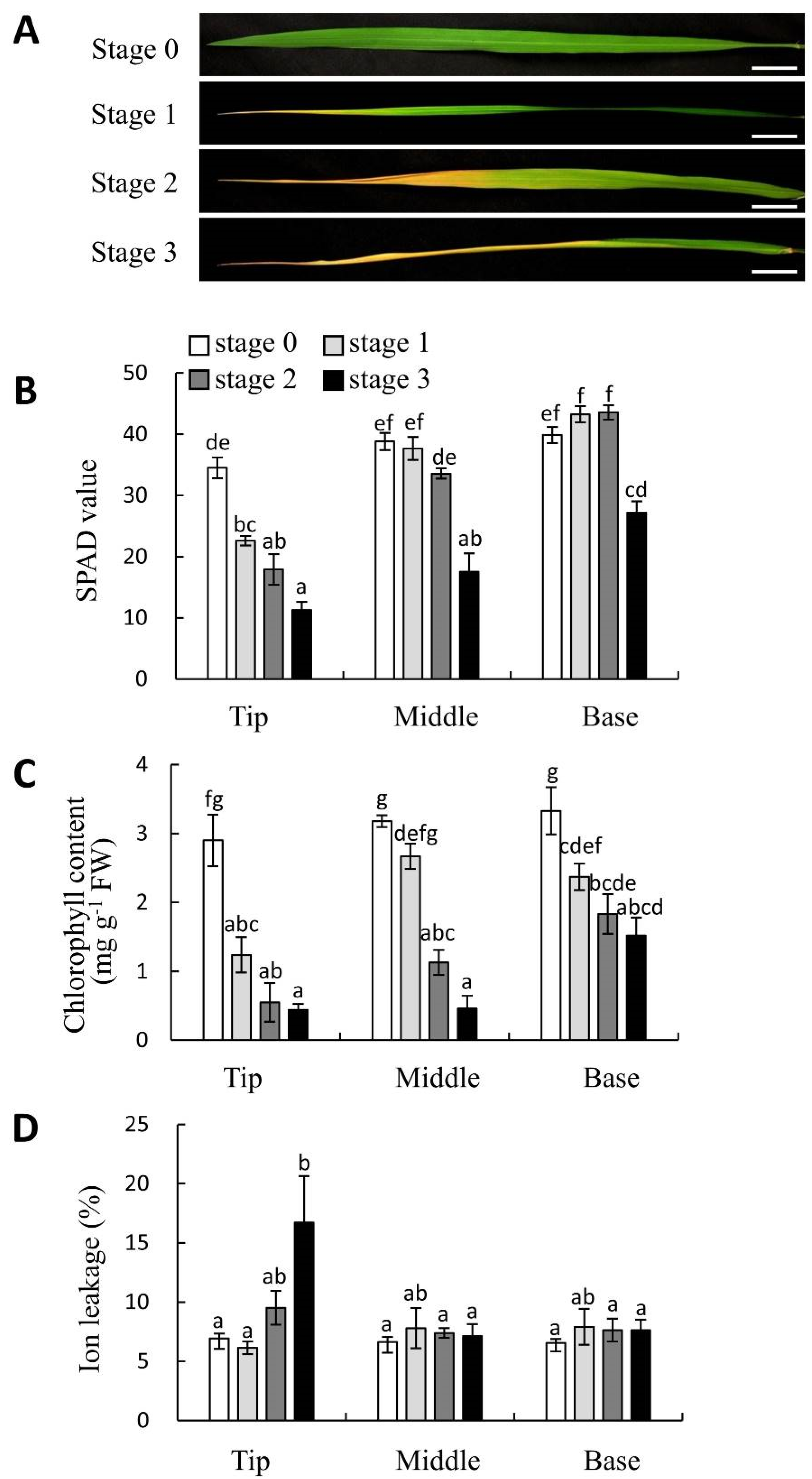
2.2. Cell Death and H2O2 Accumulation
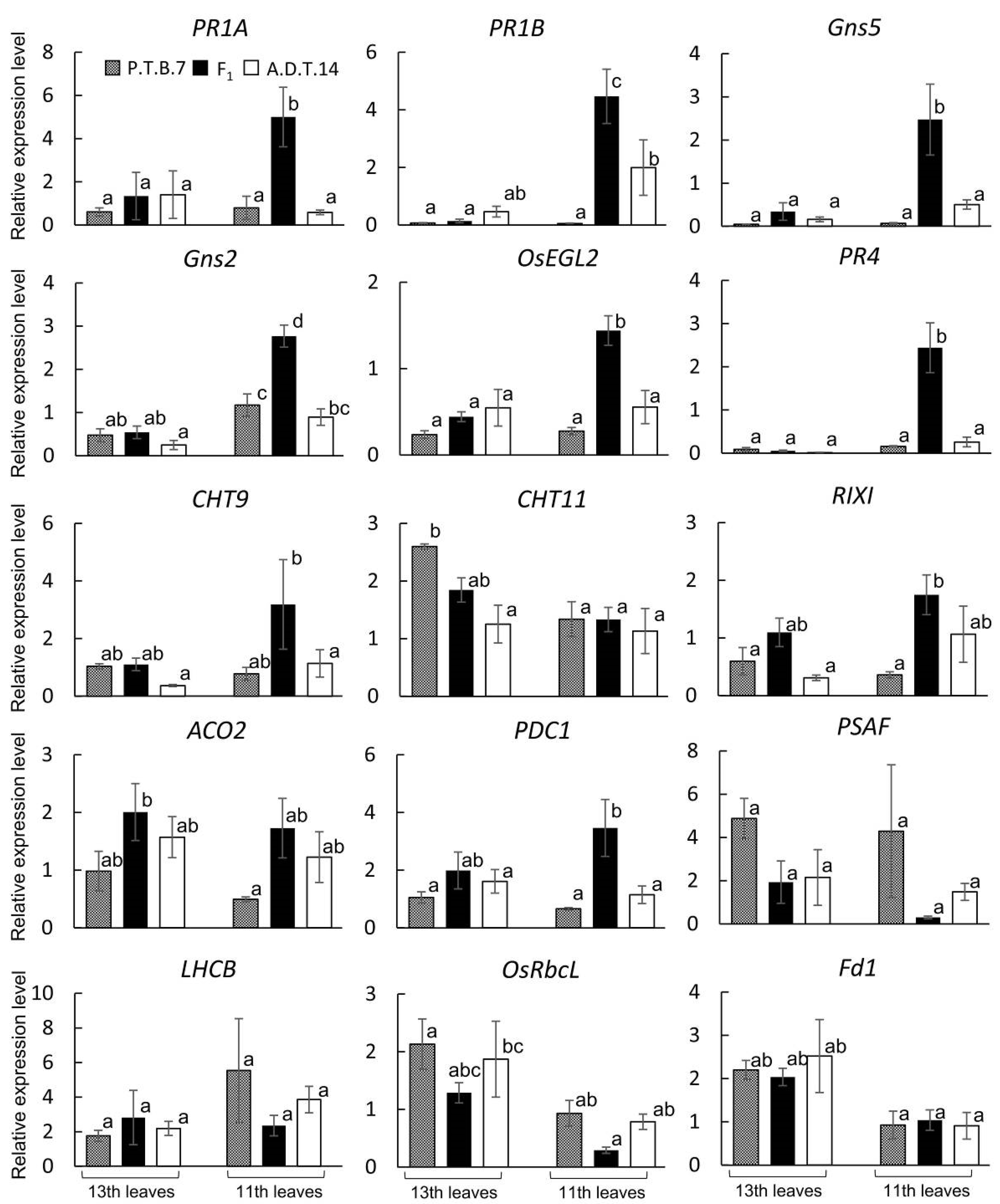
2.3. Hybrid Weakness-Related Gene Expression
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. SPAD and Chlorophyll Measurement
4.3. Ion leakage Measurement
4.4. Trypan Blue Staining
4.5. Detection of Hydrogen Peroxide Accumulation
4.6. Real-Time qRT-PCR
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Amemiya, A.; Akemine, H. Biochemical genetic studies on the root growth inhibiting complementary lethal in rice plant (Studies on the embryo culture in rice plant. 3). Bull. Natl. Inst. Agric. Sci. 1963, 10, 139–226. [Google Scholar]
- Chen, C.; Chen, H.; Lin, Y.S.; Shen, J.B.; Shan, J.X.; Qi, P.; Shi, M.; Zhu, M.Z.; Huang, X.H.; Feng, Q.; et al. A two-locus interaction causes interspecific hybrid weakness in rice. Nat. Commun. 2014, 5, 3357. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.Y.; Wang, F.; Sun, B.R.; Liu, W.G.; Li, J.H.; Deng, R.F.; Liu, D.L.; Liu, Z.R.; Zhu, M.S.; Liao, Y.L.; et al. Genetic and cytological analysis of a novel type of low temperature-dependent intrasubspecific hybrid weakness in rice. PLoS ONE 2013, 8, e73886. [Google Scholar] [CrossRef] [PubMed]
- Oka, H.I. Phylogenetic differentiation of cultivated rice. XV Complementary lethal genes in rice. Jpn. J. Genet. 1957, 32, 83–87. [Google Scholar] [CrossRef]
- Tezuka, T.; Marubashi, W. Genomic factors lead to programmed cell death during hybrid lethality in interspecific hybrids between Nicotiana tabacum and N. debneyi. SABRAO J. Breed Genet. 2006, 38, 69–81. [Google Scholar]
- Inai, S.; Ishikawa, K.; Nunomura, O.; Ikehashi, H. Genetic analysis of stunted growth by nuclear-cytoplasmic interaction in interspecific hybrids of Capsicum by using RAPD markers. Theor. Appl. Genet. 1993, 87, 416–422. [Google Scholar] [CrossRef]
- Bomblies, K.; Lempe, J.; Epple, P.; Warthmann, N.; Lanz, C.; Dangl, J.L.; Weigel, D. Autoimmune response as a mechanism for a Dobzhansky-Muller-type incompatibility syndrome in plants. PLoS Biol. 2007, 5, e236. [Google Scholar] [CrossRef]
- Tsunewaki, K. Monosomic and conventional analyses in common wheat. III Lethality. Jpn. J. Genet. 1960, 35, 71–75. [Google Scholar] [CrossRef]
- Tsunewaki, K. Aneuploid analysis of hybrid necrosis and hybrid chlorosis in tetraploid wheats using the D genome chromosome substitution lines of durum wheat. Genome 1992, 35, 594–601. [Google Scholar] [CrossRef]
- Silow, R.A. The comparative genetics of Gossypium anomalum and the cultivated Asiatic cottons. J. Genet. 1941, 259–358. [Google Scholar] [CrossRef]
- Reiber, J.M.; Neuman, D.S. Hybrid weakness in Phaseolus vulgaris L. II. Disruption of root-shoot integration. J. Plant Growth Regul. 1999, 18, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Bateson, W. Heredity and variation in modern science. In Darwin and Modern Science; Seward, A.C., Ed.; Cambridge University Press: Cambridge, UK, 1909; pp. 85–101. [Google Scholar]
- Dobzhansky, T. Genetics and the Origin of Species; Columbia University Press: New York, NY, USA, 1937. [Google Scholar]
- Muller, H.J. Isolating mechanisms, evolution, and temperature. Biol. Symp. 1942, 6, 71–124. [Google Scholar]
- Mizuno, N.; Hosogi, N.; Park, P.; Takumi, S. Hypersensitive response-like reaction is associated with hybrid necrosis in interspecific crosses between tetraploid wheat and Aegilops tauschii Coss. PLoS ONE 2010, 5, e11326. [Google Scholar] [CrossRef] [PubMed]
- Okada, M.; Yoshida, K.; Takumi, S. Hybrid incompatibilities in interspecific crosses between tetraploid wheat and its wild diploid relative Aegilops umbellulata. Plant Mol. Biol. 2017, 95, 625–645. [Google Scholar] [CrossRef]
- Ichitani, K.; Taura, S.; Tezuka, T.; Okiyama, Y.; Kuboyama, T. Chromosomal location of HWA1 and HWA2, complementary hybrid weakness genes in rice. Rice 2011, 4, 29–38. [Google Scholar] [CrossRef]
- Markwell, J.; Osterman, J.C.; Mitchell, J.L. Calibration of the Minolta SPAD-502 leaf chlorophyll meter. Photosyn. Res. 1995, 46, 467–472. [Google Scholar] [CrossRef]
- Agrawal, G.K.; Rakwal, R.; Jwa, N.S.; Agrawal, V.P. Signalling molecules and blast pathogen attack activates rice OsPR1a and OsPR1b genes: A model illustrating components participating during defence/stress response. Plant Physiol. Biochem. 2001, 39, 1095–1103. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef]
- Romero, G.O.; Simmons, C.; Yaneshita, M.; Doan, M.; Thomas, B.R.; Rodriguez, R.L. Characterization of rice endo-β-glucanase genes (Gns2–Gns14) defines a new subgroup within the gene family. Gene 1998, 223, 311–320. [Google Scholar] [CrossRef]
- Klarzynski, O.; Plesse, B.; Joubert, J.-M.; Yvin, J.-C.; Kopp, M.; Kloareg, B.; Fritig, B. Linear β-1,3 glucans are elicitors of defense responses in tobacco. Plant Physiol. 2000, 124, 1027–1038. [Google Scholar] [CrossRef]
- Kende, H. Ethylene biosynthesis. Annu. Rev. Plant. Biol. 1993, 44, 283–307. [Google Scholar] [CrossRef]
- Sugimori, M.; Kiribuchi, K.; Akimoto, C.; Yamaguchi, T.; Minami, E.; Shibuya, N.; Sobajima, H.; Cho, E.M.; Kobashi, N.; Nojiri, H.; et al. Cloning and characterization of cDNAs for the jasmonic acid-responsive genes RRJ1 and RRJ2 in suspension-cultured rice cells. Biosci. Biotechnol. Biochem. 2002, 66, 1140–1142. [Google Scholar] [CrossRef][Green Version]
- McCouch, S.R. Gene nomenclature system for rice. Rice 2008, 1, 72–84. [Google Scholar] [CrossRef]
- Saito, T.; Ichitani, K.; Suzuki, T.; Marubashi, W.; Kuboyama, T. Developmental observation and high temperature rescue from hybrid weakness in a cross between Japanese rice cultivars and Peruvian rice cultivar ‘Jamaica’. Breed. Sci. 2007, 57, 281–288. [Google Scholar] [CrossRef]
- Alcázar, R.; García, A.V.; Kronholm, I.; de Meaux, J.; Koornneef, M.; Parker, J.E.; Reymond, M. Natural variation at Strubbelig Receptor Kinase 3 drives immune-triggered incompatibilities between Arabidopsis thaliana accessions. Nat. Genet. 2010, 42, 1135–1139. [Google Scholar] [CrossRef]
- Tezuka, T.; Marubashi, W. Apoptotic cell death observed during the expression of hybrid lethality in interspecific hybrids between Nicotiana tabacum and N. suaveolens. Breed. Sci. 2004, 54, 59–66. [Google Scholar] [CrossRef]
- Yamada, T.; Marubashi, W.; Niwa, M. Apoptotic cell death induces temperature-sensitive lethality in hybrid seedlings and calli derived from the cross of Nicotiana suaveolens × N. tabacum. Planta 2000, 211, 614–622. [Google Scholar] [CrossRef]
- Huysmans, M.; Lema, A.S.; Coll, N.S.; Nowack, M.K. Dying two deaths—Programmed cell death regulation in development and disease. Curr. Opin. Plant Biol. 2017, 35, 37–44. [Google Scholar] [CrossRef]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef]
- Woo, H.R.; Kim, H.J.; Nam, H.G.; Lim, P.O. Plant leaf senescence and death—Regulation by multiple layers of control and implications for aging in general. J. Cell Sci. 2013, 126, 4823. [Google Scholar] [CrossRef]
- Huang, X.; Li, J.; Bao, F.; Zhang, X.; Yang, S. A gain-of-function mutation in the Arabidopsis disease resistance gene RPP4 confers sensitivity to low temperature. Plant Physiol. 2010, 154, 796–809. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Hua, J. A haplotype-specific resistance gene regulated by BONZAI1 mediates temperature-dependent growth control in Arabidopsis. Plant Cell 2004, 16, 1060–1071. [Google Scholar] [CrossRef] [PubMed]
- Coll, N.S.; Epple, P.; Dangl, J.L. Programmed cell death in the plant immune system. Cell Death Differ. 2011, 18, 1247. [Google Scholar] [CrossRef] [PubMed]
- Overmyer, K.; Brosché, M.; Kangasjärvi, J. Reactive oxygen species and hormonal control of cell death. Trends Plant Sci. 2003, 8, 335–342. [Google Scholar] [CrossRef]
- Jajic, I.; Sarna, T.; Strzalka, K. Senescence, stress, and reactive oxygen species. Plants 2015, 4, 393–411. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Rizhsky, L.; Shulaev, V.; Mittler, R. Measuring programmed cell death in plants. In Apoptosis Methods and Protocols; Brady, H.J.M., Ed.; Humana Press: Totowa, NJ, USA, 2004; pp. 179–189. [Google Scholar]
- Zhang, H.K.; Zhang, X.; Mao, B.Z.; Li, Q.; He, Z.H. Alpha-picolinic acid, a fungal toxin and mammal apoptosis-inducing agent, elicits hypersensitive-like response and enhances disease resistance in rice. Cell Res. 2004, 14, 27–33. [Google Scholar] [CrossRef]
- Chandru, H.K.; Kim, E.; Kuk, Y.; Cho, K.; Han, O. Kinetics of wound-induced activation of antioxidative enzymes in Oryza sativa: Differential activation at different growth stages. Plant Sci. 2003, 164, 935–941. [Google Scholar] [CrossRef]
- Yamamoto, E.; Takashi, T.; Morinaka, Y.; Lin, S.; Wu, J.; Matsumoto, T.; Kitano, H.; Matsuoka, M.; Ashikari, M. Gain of deleterious function causes an autoimmune response and Bateson–Dobzhansky–Muller incompatibility in rice. Mol. Genet. Genomics 2010, 283, 305–315. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | RAP-DB locus ID a | CGSNL Gene Name b | Description |
|---|---|---|---|
| PR1A | Os07g0129200 | PATHOGENESIS-RELATED GENE 1A | Similar to Pathogenesis-related protein PR1a |
| PR1B | Os01g0382000 | PATHOGENESIS-RELATED GENE 1B | Similar to Pathogenesis-related protein PRB1-2 precursor |
| Gns5 | Os01g0940700 | Similar to Glucan endo-1,3-beta-glucosidase GII precursor | |
| Gns2 | Os01g0944900 | Similar to Glucan endo-1,3-beta-D-glucosidase | |
| OsEGL2 | Os01g0942300 | Similar to Beta glucanase precursor | |
| PR4 | Os11g0592200 | PATHOGENESIS-RELATED GENE 4 | Similar to Chitin-binding allergen Bra r 2 |
| CHT9 | Os05g0399400 | CHITINASE 9 | Chitinase 9 |
| CHT11 | Os03g0132900 | CHITINASE 11 | Similar to Chitinase 11 |
| RIXI | Os11g0701800 | Chitinase III C10701-rice (Class III chitinase homologue) | |
| ACO2 | Os09g0451000 | AMINOCYCLOPROPANE-1-CARBOXYLIC ACID OXIDASE 2 | Similar to 1-aminocyclopropane-1-carboxylase 1 |
| PDC1 | Os05g0469600 | PYRUVATE DECARBOXYLASE 1 | Similar to Pyruvate decarboxylase |
| PSAF | Os03g0778100 | PHOTOSYSTEM I SUBUNIT | Similar to Photosystem-1 F subunit |
| LHCB | Os03g0592500 | Similar to Photosystem II type II chlorophyll a/b binding protein | |
| OsRbcL | Os06g0598500 | Similar to Ribulose bisphosphate carboxylase large chain precursor | |
| Fd1 | Os08g0104600 | Ferredoxin I, chloroplast precursor | |
| Actin | Os05g0438800 | Similar to Actin1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shiragaki, K.; Iizuka, T.; Ichitani, K.; Kuboyama, T.; Morikawa, T.; Oda, M.; Tezuka, T. HWA1- and HWA2-Mediated Hybrid Weakness in Rice Involves Cell Death, Reactive Oxygen Species Accumulation, and Disease Resistance-Related Gene Upregulation. Plants 2019, 8, 450. https://doi.org/10.3390/plants8110450
Shiragaki K, Iizuka T, Ichitani K, Kuboyama T, Morikawa T, Oda M, Tezuka T. HWA1- and HWA2-Mediated Hybrid Weakness in Rice Involves Cell Death, Reactive Oxygen Species Accumulation, and Disease Resistance-Related Gene Upregulation. Plants. 2019; 8(11):450. https://doi.org/10.3390/plants8110450
Chicago/Turabian StyleShiragaki, Kumpei, Takahiro Iizuka, Katsuyuki Ichitani, Tsutomu Kuboyama, Toshinobu Morikawa, Masayuki Oda, and Takahiro Tezuka. 2019. "HWA1- and HWA2-Mediated Hybrid Weakness in Rice Involves Cell Death, Reactive Oxygen Species Accumulation, and Disease Resistance-Related Gene Upregulation" Plants 8, no. 11: 450. https://doi.org/10.3390/plants8110450
APA StyleShiragaki, K., Iizuka, T., Ichitani, K., Kuboyama, T., Morikawa, T., Oda, M., & Tezuka, T. (2019). HWA1- and HWA2-Mediated Hybrid Weakness in Rice Involves Cell Death, Reactive Oxygen Species Accumulation, and Disease Resistance-Related Gene Upregulation. Plants, 8(11), 450. https://doi.org/10.3390/plants8110450