Emerging Functions for Cell Wall Polysaccharides Accumulated during Eudicot Seed Development



Abstract
1. Introduction
2. General Overview of Polysaccharide Production during Arabidopsis Seed Development
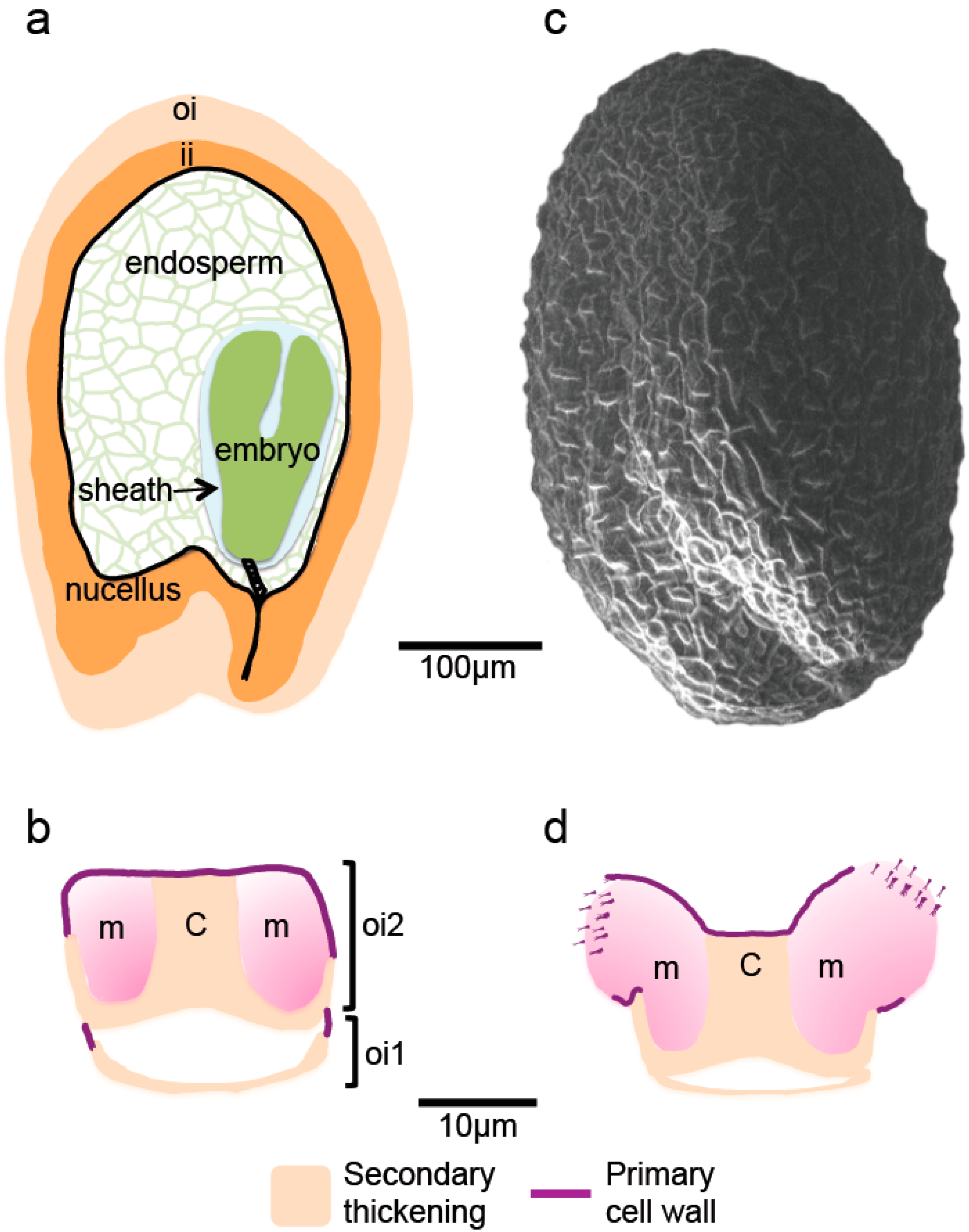
3. Localized Accumulation and Remodeling of Cell Wall Polysaccharides during Eudicot Seed Development
3.1. Distinctive Polysaccharide Accumulation Patterns in the Embryo and Endosperm
3.2. Specialization of the Seed Coat through Polysaccharide Accumulation
4. Functional Roles of Polysaccharides during Seed Development
4.1. Cell Wall Biosynthesis Defects that Induce Embryo Lethality
4.2. Polysaccharides in Seed Cell Division, Expansion and Elimination
4.3. Diverse Functions for Seed Coat Polysaccharides
5. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Wolf, S.; Hematy, K.; Hofte, H. Growth control and cell wall signaling in plants. Annu. Rev. Plant Biol. 2012, 63, 381–407. [Google Scholar] [CrossRef] [PubMed]
- Finch-Savage, W.E.; Bassel, G.W. Seed vigour and crop establishment: Extending performance beyond adaptation. J. Exp. Bot. 2016, 67, 567–591. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Re-constructing our models of cellulose and primary cell wall assembly. Curr. Opin. Plant Biol. 2014, 22, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Pauly, M.; Keegstra, K. Biosynthesis of the plant cell wall matrix polysaccharide xyloglucan. Annu. Rev. Plant Biol. 2016, 67, 235–259. [Google Scholar] [CrossRef] [PubMed]
- Tuomivaara, S.T.; Yaoi, K.; O’Neill, M.A.; York, W.S. Generation and structural validation of a library of diverse xyloglucan-derived oligosaccharides, including an update on xyloglucan nomenclature. Carbohydr. Res. 2015, 402, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Schultink, A.; Liu, L.; Zhu, L.; Pauly, M. Structural diversity and function of xyloglucan sidechain substituents. Plants 2014, 3, 526–542. [Google Scholar] [CrossRef] [PubMed]
- Burton, R.A.; Gidley, M.J.; Fincher, G.B. Heterogeneity in the chemistry, structure and function of plant cell walls. Nat. Chem. Biol. 2010, 6, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Lyczakowski, J.J.; Pereira, C.S.; Kotake, T.; Yu, X.; Li, A.; Mogelsvang, S.; Skaf, M.S.; Dupree, P. The patterned structure of galactoglucomannan suggests it may bind to cellulose in seed mucilage. Plant Physiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Mohnen, D. Pectin structure and biosynthesis. Curr. Opin. Plant Biol. 2008, 11, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Zheng, Y.; Dong, Q.; Fang, J. Structural analysis of a pectic polysaccharide from the leaves of diospyros kaki. Phytochemistry 2004, 65, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Ralet, M.C.; Crepeau, M.J.; Vigouroux, J.; Tran, J.; Berger, A.; Salle, C.; Granier, F.; Botran, L.; North, H.M. Xylans provide the structural driving force for mucilage adhesion to the Arabidopsis seed coat. Plant Physiol. 2016, 171, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Piršelová, B.; Matušíková, I. Callose: The plant cell wall polysaccharide with multiple biological functions. Acta Physiol. Plant. 2013, 35, 635–644. [Google Scholar] [CrossRef]
- Verbančič, J.; Lunn, J.E.; Stitt, M.; Persson, S. Carbon supply and the regulation of cell wall synthesis. Mol. Plant 2018, 11, 75–94. [Google Scholar] [CrossRef] [PubMed]
- Zykwinska, A.W.; Ralet, M.C.; Garnier, C.D.; Thibault, J.F. Evidence for in vitro binding of pectin side chains to cellulose. Plant Physiol. 2005, 139, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J.; Jarvis, M.C. Comparative structure and biomechanics of plant primary and secondary cell walls. Front. Plant Sci. 2012, 3, 204. [Google Scholar] [CrossRef] [PubMed]
- Ten Hove, C.A.; Lu, K.-J.; Weijers, D. Building a plant: Cell fate specification in the early Arabidopsis embryo. Development 2015, 142, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Van Dop, M.; Liao, C.Y.; Weijers, D. Control of oriented cell division in the Arabidopsis embryo. Curr. Opin. Plant Biol. 2015, 23, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Miart, F.; Desprez, T.; Biot, E.; Morin, H.; Belcram, K.; Hofte, H.; Gonneau, M.; Vernhettes, S. Spatio-temporal analysis of cellulose synthesis during cell plate formation in Arabidopsis. Plant J. 2014, 77, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Berger, F. Endosperm: Food for humankind and fodder for scientific discoveries. New Phytol. 2012, 195, 290–305. [Google Scholar] [CrossRef] [PubMed]
- Lafon-Placette, C.; Kohler, C. Embryo and endosperm, partners in seed development. Curr. Opin. Plant Biol. 2014, 17, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Otegui, M.; Staehelin, L.A. Syncytial-type cell plates: A novel kind of cell plate involved in endosperm cellularization of Arabidopsis. Plant Cell 2000, 12, 933–947. [Google Scholar] [CrossRef] [PubMed]
- Otegui, M.S.; Mastronarde, D.N.; Kang, B.H.; Bednarek, S.Y.; Staehelin, L.A. Three-dimensional analysis of syncytial-type cell plates during endosperm cellularization visualized by high resolution electron tomography. Plant Cell 2001, 13, 2033–2051. [Google Scholar] [CrossRef] [PubMed]
- Ingram, G.C. Dying to live: Cell elimination as a developmental strategy in angiosperm seeds. J. Exp. Bot. 2017, 68, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Fiume, E.; Coen, O.; Pechoux, C.; Lepiniec, L.; Magnani, E. Endosperm and nucellus develop antagonistically in Arabidopsis seeds. Plant Cell 2016, 28, 1343–1360. [Google Scholar] [CrossRef] [PubMed]
- Western, T.L.; Skinner, D.J.; Haughn, G.W. Differentiation of mucilage secretory cells of the Arabidopsis seed coat. Plant Physiol. 2000, 122, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Coen, O.; Magnani, E. Seed coat thickness in the evolution of angiosperms. Cell. Mol. Life Sci. 2018, 75, 2509–2518. [Google Scholar] [CrossRef] [PubMed]
- Samuels, A.L.; Giddings, T.H., Jr.; Staehelin, L.A. Cytokinesis in tobacco BY-2 and root tip cells: A new model of cell plate formation in higher plants. J. Cell Biol. 1995, 130, 1345–1357. [Google Scholar] [CrossRef] [PubMed]
- Thiele, K.; Wanner, G.; Kindzierski, V.; Jurgens, G.; Mayer, U.; Pachl, F.; Assaad, F.F. The timely deposition of callose is essential for cytokinesis in Arabidopsis. Plant J. 2009, 58, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Otegui, M.S. Endosperm cell walls: Formation, composition, and functions. In Endosperm: Developmental and Molecular Biology; Olsen, O.-A., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 159–177. [Google Scholar]
- Vanzin, G.F.; Madson, M.; Carpita, N.C.; Raikhel, N.V.; Keegstra, K.; Reiter, W.D. The mur2 mutant of Arabidopsis thaliana lacks fucosylated xyloglucan because of a lesion in fucosyltransferase AtFUT1. Proc. Natl. Acad. Sci. USA 2002, 99, 3340–3345. [Google Scholar] [CrossRef] [PubMed]
- Sechet, J.; Frey, A.; Effroy-Cuzzi, D.; Berger, A.; Perreau, F.; Cueff, G.; Charif, D.; Rajjou, L.; Mouille, G.; North, H.M.; et al. Xyloglucan metabolism differentially impacts the cell wall characteristics of the endosperm and embryo during Arabidopsis seed germination. Plant Physiol. 2016, 170, 1367–1380. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; Cornuault, V.; Manfield, I.W.; Ralet, M.C.; Knox, J.P. Multi-scale spatial heterogeneity of pectic rhamnogalacturonan I (RG-I) structural features in tobacco seed endosperm cell walls. Plant J. 2013, 75, 1018–1027. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Valderrama, J.E.; Jimenez-Duran, K.; Zuniga-Sanchez, E.; Salazar-Iribe, A.; Marquez-Guzman, J.; Gamboa-deBuen, A. Degree of pectin methyl esterification in endosperm cell walls is involved in embryo bending in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2018, 495, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Müller, K.; Levesque-Tremblay, G.; Bartels, S.; Weitbrecht, K.; Wormit, A.; Usadel, B.; Haughn, G.; Kermode, A.R. Demethylesterification of cell wall pectins in Arabidopsis plays a role in seed germination. Plant Physiol. 2013, 161, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Levesque-Tremblay, G.; Muller, K.; Mansfield, S.D.; Haughn, G.W. Highly methyl esterified seeds is a pectin methyl esterase involved in embryo development. Plant Physiol. 2015, 167, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Saez-Aguayo, S.; Ralet, M.C.; Berger, A.; Botran, L.; Ropartz, D.; Marion-Poll, A.; North, H.M. PECTIN METHYLESTERASE INHIBITOR6 promotes Arabidopsis mucilage release by limiting methylesterification of homogalacturonan in seed coat epidermal cells. Plant Cell 2013, 25, 308–323. [Google Scholar] [CrossRef] [PubMed]
- Fourquin, C.; Beauzamy, L.; Chamot, S.; Creff, A.; Goodrich, J.; Boudaoud, A.; Ingram, G. Mechanical stress mediated by both endosperm softening and embryo growth underlies endosperm elimination in Arabidopsis seeds. Development 2016, 143, 3300–3305. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; Dekkers, B.J.; Steinbrecher, T.; Walsh, C.T.; Bacic, A.; Bentsink, L.; Leubner-Metzger, G.; Knox, J.P. Distinct cell wall architectures in seed endosperms in representatives of the Brassicaceae and Solanaceae. Plant Physiol. 2012, 160, 1551–1566. [Google Scholar] [CrossRef] [PubMed]
- Naran, R.; Chen, G.; Carpita, N.C. Novel rhamnogalacturonan I and arabinoxylan polysaccharides of flax seed mucilage. Plant Physiol. 2008, 148, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Arsovski, A.A.; Popma, T.M.; Haughn, G.W.; Carpita, N.C.; McCann, M.C.; Western, T.L. AtBXL1 encodes a bifunctional beta-d-xylosidase/alpha-l-arabinofuranosidase required for pectic arabinan modification in Arabidopsis mucilage secretory cells. Plant Physiol. 2009, 150, 1219–1234. [Google Scholar] [CrossRef] [PubMed]
- Dean, G.H.; Zheng, H.; Tewari, J.; Huang, J.; Young, D.S.; Hwang, Y.T.; Western, T.L.; Carpita, N.C.; McCann, M.C.; Mansfield, S.D.; et al. The Arabidopsis MUM2 gene encodes a beta-galactosidase required for the production of seed coat mucilage with correct hydration properties. Plant Cell 2007, 19, 4007–4021. [Google Scholar] [CrossRef] [PubMed]
- Macquet, A.; Ralet, M.C.; Loudet, O.; Kronenberger, J.; Mouille, G.; Marion-Poll, A.; North, H.M. A naturally occurring mutation in an Arabidopsis accession affects a beta-d-galactosidase that increases the hydrophilic potential of rhamnogalacturonan I in seed mucilage. Plant Cell 2007, 19, 3990–4006. [Google Scholar] [CrossRef] [PubMed]
- Hall, Q.; Cannon, M.C. The cell wall hydroxyproline-rich glycoprotein RSH is essential for normal embryo development in Arabidopsis. Plant Cell 2002, 14, 1161–1172. [Google Scholar] [CrossRef] [PubMed]
- Cannon, M.C.; Terneus, K.; Hall, Q.; Tan, L.; Wang, Y.; Wegenhart, B.L.; Chen, L.; Lamport, D.T.A.; Chen, Y.; Kieliszewski, M.J. Self-assembly of the plant cell wall requires an extensin scaffold. Proc. Natl. Acad. Sci. USA 2008, 105, 2226–2231. [Google Scholar] [CrossRef] [PubMed]
- Goubet, F.; Barton, C.J.; Mortimer, J.C.; Yu, X.; Zhang, Z.; Miles, G.P.; Richens, J.; Liepman, A.H.; Seffen, K.; Dupree, P. Cell wall glucomannan in Arabidopsis is synthesised by CSLA glycosyltransferases, and influences the progression of embryogenesis. Plant J. 2009, 60, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Goubet, F.; Misrahi, A.; Park, S.K.; Zhang, Z.; Twell, D.; Dupree, P. Atcsla7, a cellulose synthase-like putative glycosyltransferase, is important for pollen tube growth and embryogenesis in Arabidopsis. Plant Physiol. 2003, 131, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Andujar, C.; Pluskota, W.E.; Bassel, G.W.; Asahina, M.; Pupel, P.; Nguyen, T.T.; Takeda-Kamiya, N.; Toubiana, D.; Bai, B.; Gorecki, R.J.; et al. Mechanisms of hormonal regulation of endosperm cap-specific gene expression in tomato seeds. Plant J. 2012, 71, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Voiniciuc, C.; Schmidt, M.H.; Berger, A.; Yang, B.; Ebert, B.; Scheller, H.V.; North, H.M.; Usadel, B.; Gunl, M. MUCILAGE-RELATED10 produces galactoglucomannan that maintains pectin and cellulose architecture in Arabidopsis seed mucilage. Plant Physiol. 2015, 169, 403–420. [Google Scholar] [CrossRef] [PubMed]
- Macquet, A.; Ralet, M.C.; Kronenberger, J.; Marion-Poll, A.; North, H.M. In situ, chemical and macromolecular study of the composition of Arabidopsis thaliana seed coat mucilage. Plant Cell Physiol. 2007, 48, 984–999. [Google Scholar] [CrossRef] [PubMed]
- Gillmor, C.S.; Poindexter, P.; Lorieau, J.; Palcic, M.M.; Somerville, C. Alpha-glucosidase I is required for cellulose biosynthesis and morphogenesis in Arabidopsis. J. Cell Biol. 2002, 156, 1003–1013. [Google Scholar] [CrossRef] [PubMed]
- Stork, J.; Harris, D.; Griffiths, J.; Williams, B.; Beisson, F.; Li-Beisson, Y.; Mendu, V.; Haughn, G.; Debolt, S. CELLULOSE SYNTHASE9 serves a nonredundant role in secondary cell wall synthesis in Arabidopsis epidermal testa cells. Plant Physiol. 2010, 153, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Mendu, V.; Griffiths, J.S.; Persson, S.; Stork, J.; Downie, A.B.; Voiniciuc, C.; Haughn, G.W.; DeBolt, S. Subfunctionalization of cellulose synthases in seed coat epidermal cells mediates secondary radial wall synthesis and mucilage attachment. Plant Physiol. 2011, 157, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Barnes, W.J.; Anderson, C.T. Cytosolic invertases contribute to cellulose biosynthesis and influence carbon partitioning in seedlings of Arabidopsis thaliana. Plant J. 2018, 94, 956–974. [Google Scholar] [CrossRef] [PubMed]
- Haigler, C.H.; Betancur, L.; Stiff, M.R.; Tuttle, J.R. Cotton fiber: A powerful single-cell model for cell wall and cellulose research. Front. Plant Sci. 2012, 3, 104. [Google Scholar] [CrossRef] [PubMed]
- Phan, J.L.; Tucker, M.R.; Khor, S.F.; Shirley, N.; Lahnstein, J.; Beahan, C.; Bacic, A.; Burton, R.A. Differences in glycosyltransferase family 61 accompany variation in seed coat mucilage composition in Plantago spp. J. Exp. Bot. 2016, 67, 6481–6495. [Google Scholar] [CrossRef] [PubMed]
- McCartney, L.; Ormerod, A.P.; Gidley, M.J.; Knox, J.P. Temporal and spatial regulation of pectic (1→4)-beta-d-galactan in cell walls of developing pea cotyledons: Implications for mechanical properties. Plant J. 2000, 22, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Buckeridge, M.S. Seed cell wall storage polysaccharides: Models to understand cell wall biosynthesis and degradation. Plant Physiol. 2010, 154, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Pauly, M.; Keegstra, K. Plant cell wall polymers as precursors for biofuels. Curr. Opin. Plant Biol. 2010, 13, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Cholewa, E.; Mohamed, T.; Peterson, C.A.; Gijzen, M. Cracks in the palisade cuticle of soybean seed coats correlate with their permeability to water. Ann. Bot. 2004, 94, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Western, T.L. The sticky tale of seed coat mucilages: Production, genetics, and role in seed germination and dispersal. Seed Sci. Res. 2012, 22, 1–25. [Google Scholar] [CrossRef]
- Yu, L.; Shi, D.; Li, J.; Kong, Y.; Yu, Y.; Chai, G.; Hu, R.; Wang, J.; Hahn, M.G.; Zhou, G. CELLULOSE SYNTHASE-LIKE A2, a glucomannan synthase, is involved in maintaining adherent mucilage structure in Arabidopsis seed. Plant Physiol. 2014, 164, 1842–1856. [Google Scholar] [CrossRef] [PubMed]
- Rautengarten, C.; Usadel, B.; Neurnetzler, L.; Hartmann, J.; Buessis, D.; Altmann, T. A subtilisin-like serine protease essential for mucilage release from Arabidopsis seed coats. Plant J. 2008, 54, 466–480. [Google Scholar] [CrossRef] [PubMed]
- Kunieda, T.; Shimada, T.; Kondo, M.; Nishimura, M.; Nishitani, K.; Hara-Nishimura, I. Spatiotemporal secretion of PEROXIDASE36 is required for seed coat mucilage extrusion in Arabidopsis. Plant Cell 2013, 25, 1355–1367. [Google Scholar] [CrossRef] [PubMed]
- Cocuron, J.-C.; Lerouxel, O.; Drakakaki, G.; Alonso, A.P.; Liepman, A.H.; Keegstra, K.; Raikhel, N.; Wilkerson, C.G. A gene from the cellulose synthase-like C family encodes a β-1,4 glucan synthase. Proc. Natl. Acad. Sci. USA 2007, 104, 8550. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Mortimer, J.C.; Davis, J.; Dupree, P.; Keegstra, K. Identification of an additional protein involved in mannan biosynthesis. Plant J. 2013, 73, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.K.; Johnson, N.; Wilkerson, C.G. Discovery of diversity in xylan biosynthetic genes by transcriptional profiling of a heteroxylan containing mucilaginous tissue. Front. Plant Sci. 2013, 4, 183. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, Y.; Kato, K.; Ogawa-Ohnishi, M.; Tsuruhama, K.; Kajiura, H.; Yagyu, K.; Takeda, A.; Takeda, Y.; Kunieda, T.; Hara-Nishimura, I.; et al. Pectin RG-I rhamnosyltransferases represent a novel plant-specific glycosyltransferase family. Nat. Plants 2018, 4, 669–676. [Google Scholar] [CrossRef] [PubMed]
- North, H.M.; Berger, A.; Saez-Aguayo, S.; Ralet, M.C. Understanding polysaccharide production and properties using seed coat mutants: Future perspectives for the exploitation of natural variants. Ann. Bot. 2014, 114, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Persson, S.; Paredez, A.; Carroll, A.; Palsdottir, H.; Doblin, M.; Poindexter, P.; Khitrov, N.; Auer, M.; Somerville, C.R. Genetic evidence for three unique components in primary cell-wall cellulose synthase complexes in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 15566–15571. [Google Scholar] [CrossRef] [PubMed]
- Liepman, A.; Cavalier, D. The cellulose synthase-like A and cellulose synthase-like C families: Recent advances and future perspectives. Front. Plant Sci. 2012, 3, 109. [Google Scholar] [CrossRef] [PubMed]
- Liepman, A.H.; Wilkerson, C.G.; Keegstra, K. Expression of cellulose synthase-like (Csl) genes in insect cells reveals that CslA family members encode mannan synthases. Proc. Natl. Acad. Sci. USA 2005, 102, 2221–2226. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Liu, L.; Lee, E.; Han, X.; Rim, Y.; Chu, H.; Kim, S.W.; Sack, F.; Kim, J.Y. The Arabidopsis callose synthase gene GSL8 is required for cytokinesis and cell patterning. Plant Physiol. 2009, 150, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Johnston, N.; Talideh, E.; Mitchell, S.; Jeffree, C.; Goodrich, J.; Ingram, G. The endosperm-specific ZHOUPI gene of Arabidopsis thaliana regulates endosperm breakdown and embryonic epidermal development. Development 2008, 135, 3501–3509. [Google Scholar] [CrossRef] [PubMed]
- Denay, G.; Creff, A.; Moussu, S.; Wagnon, P.; Thevenin, J.; Gerentes, M.F.; Chambrier, P.; Dubreucq, B.; Ingram, G. Endosperm breakdown in Arabidopsis requires heterodimers of the basic helix-loop-helix proteins ZHOUPI and INDUCER OF CBP EXPRESSION 1. Development 2014, 141, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Xing, Q.; Creff, A.; Waters, A.; Tanaka, H.; Goodrich, J.; Ingram, G.C. ZHOUPI controls embryonic cuticle formation via a signalling pathway involving the subtilisin protease abnormal LEAF-SHAPE1 and the receptor kinases GASSHO1 and GASSHO2. Development 2013, 140, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Moussu, S.; Doll, N.M.; Chamot, S.; Brocard, L.; Creff, A.; Fourquin, C.; Widiez, T.; Nimchuk, Z.L.; Ingram, G. ZHOUPI and KERBEROS mediate embryo/endosperm separation by promoting the formation of an extracuticular sheath at the embryo surface. Plant Cell 2017, 29, 1642–1656. [Google Scholar] [CrossRef] [PubMed]
- Penfield, S. Seed dormancy and germination. Curr. Biol. 2017, 27, R874–R878. [Google Scholar] [CrossRef] [PubMed]
- Mohamed-Yasseen, Y.; Barringer, S.A.; Splittstoesser, W.E.; Costanza, S. The role of seed coats in seed viability. Bot. Rev. 1994, 60, 426–439. [Google Scholar] [CrossRef]
- Creff, A.; Brocard, L.; Ingram, G. A mechanically sensitive cell layer regulates the physical properties of the Arabidopsis seed coat. Nat. Commun. 2015, 6, 6382. [Google Scholar] [CrossRef] [PubMed]
- Donohue, K.; Polisetty, C.R.; Wender, N.J. Genetic basis and consequences of niche construction: Plasticity-induced genetic constraints on the evolution of seed dispersal in Arabidopsis thaliana. Am. Nat. 2005, 165, 537–550. [Google Scholar] [PubMed]
- Betancur, L.; Singh, B.; Rapp, R.A.; Wendel, J.F.; Marks, M.D.; Roberts, A.W.; Haigler, C.H. Phylogenetically distinct cellulose synthase genes support secondary wall thickening in Arabidopsis shoot trichomes and cotton fiber. J. Integr. Plant Biol. 2010, 52, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Saez-Aguayo, S.; Rondeau-Mouro, C.; Macquet, A.; Kronholm, I.; Ralet, M.C.; Berger, A.; Salle, C.; Poulain, D.; Granier, F.; Botran, L.; et al. Local evolution of seed flotation in Arabidopsis. PLoS Genet. 2014, 10, e1004221. [Google Scholar] [CrossRef] [PubMed]
- Di Marsico, A.; Scrano, L.; Amato, M.; Gamiz, B.; Real, M.; Cox, L. Mucilage from seeds of chia (Salvia hispanica L.) used as soil conditioner; effects on the sorption-desorption of four herbicides in three different soils. Sci. Total Environ. 2018, 625, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, J.; Galloway, A.F.; Nikolopoulos, G.; Field, K.J.; Knox, P. A quantitative method for the high throughput screening for the soil adhesion properties of plant and microbial polysaccharides and exudates. Plant Soil 2018, 428, 57–65. [Google Scholar] [CrossRef]
- Barnes, W.J.; Anderson, C.T. Release, recycle, rebuild: Cell-wall remodeling, autodegradation, and sugar salvage for new wall biosynthesis during plant development. Mol. Plant 2018, 11, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Engelsdorf, T.; Gigli-Bisceglia, N.; Veerabagu, M.; McKenna, J.F.; Vaahtera, L.; Augstein, F.; Van der Does, D.; Zipfel, C.; Hamann, T. The plant cell wall integrity maintenance and immune signaling systems cooperate to control stress responses in Arabidopsis thaliana. Sci. Signal. 2018, 11, eaao3070. [Google Scholar] [CrossRef] [PubMed]
- Munz, E.; Rolletschek, H.; Oeltze-Jafra, S.; Fuchs, J.; Guendel, A.; Neuberger, T.; Ortleb, S.; Jakob, P.M.; Borisjuk, L. A functional imaging study of germinating oilseed rape seed. New Phytol. 2017, 216, 1181–1190. [Google Scholar] [CrossRef] [PubMed]
- Gierlinger, N. New insights into plant cell walls by vibrational microspectroscopy. Appl. Spectrosc. Rev. 2018, 53, 517–551. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Polysaccharide | Species | Tissue | Seed Developmental Stage when Detected | Known Function [Reference] | |
|---|---|---|---|---|---|
| Pectin | HG | Arabidopsis | Embryo | Bent cotyledon, mature seed | Control of cell expansion by the degree of pectin methylesterification [34,35] |
| Low- and de-esterified HG | Arabidopsis | Endosperm | Bent cotyledon, mature seed | Modulation of the elasticity of the endosperm and its ability to be degraded [33,35] | |
| Limited HG demethylesterification | Arabidopsis | Seed coat epidermal cells | Throughout development | Localized fragmentation of primary cell wall [36] | |
| Arabinan-rich RGI | Arabidopsis | Endosperm | Throughout development | Controlling endosperm breakdown [37] | |
| Arabinan-rich RGI | Arabidopsis Cress Tobacco | Endosperm | Mature seed | Mechanical resistance against radicle protrusion [32,38] | |
| RGI | Arabidopsis Flaxseed | Seed coat epidermal cells | From linear cotyledon to mature seed | Mucilage, reduction of seed buoyancy [39,40,41,42] | |
| Extensin/pectin interaction | Arabidopsis | Cytokinesis, cell plate formation [43,44] | |||
| Mannan | Arabidopsis | Embryo | Involved in embryogenesis [45,46] | ||
| Arabidopsis Tobacco Tomato | Endosperm | Mature seed | Formation of endosperm cell walls (embryo lethal mutants) [46]. Rigidity of endosperm tissues, degraded to allow radicle protrusion [38,47] | ||
| GGM | Arabidopsis | Seed coat epidermal cells | Mature seed | Mucilage structure [48] | |
| Callose | Arabidopsis | Embryo | Transiently present in dividing cells | Cytokinesis, cell plate formation [28] | |
| Arabidopsis | Endosperm | Throughout development | Cytokinesis, cell plate formation [21,22] | ||
| Arabidopsis | Seed coat epidermal cells | Mature seed | Columella formation [49] | ||
| Soybean | Sclereids | ||||
| Xyloglucan | Galactosylated | Arabidopsis | Endosperm | Before endosperm cellularization | |
| Galactosylated/Fucosylated | Arabidopsis | Embryo, seed coat | Before endosperm cellularization | ||
| Galactosylated/Fucosylated | Arabidopsis | Embryo Endosperm Seed coat epidermal cells | From bent cotyledon to mature seed | Mechanical resistance of endosperm against radicle protrusion [31] | |
| Cellulose | Arabidopsis | Embryo | All seed stages | Orientation of anisotropic cell elongation and organ expansion [50] | |
| Arabidopsis | Endosperm | Low throughout development | |||
| Arabidopsis | Seed coat (outer integument and columella) | Throughout development | Mechanical resistance and columella formation [51,52,53] | ||
| Arabidopsis | Seed coat epidermal cells | Mature seed | Attachment of mucilage pectin to seed coat [11,14] | ||
| Cotton | Seed trichome | Throughout development | Cotton fiber [54] | ||
| Xylan | Arabidopsis | Seed coat epidermal cells | Mature seed | Attachment of mucilage pectin to seed coat [11] | |
| Heteroxylan | Psyllium | Seed coat epidermal cells | Mature seed | Mucilage [55] | |
| AX | Flaxseed | Seed coat epidermal cells | Mature seed | Mucilage [39] | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sechet, J.; Marion-Poll, A.; North, H.M. Emerging Functions for Cell Wall Polysaccharides Accumulated during Eudicot Seed Development. Plants 2018, 7, 81. https://doi.org/10.3390/plants7040081
Sechet J, Marion-Poll A, North HM. Emerging Functions for Cell Wall Polysaccharides Accumulated during Eudicot Seed Development. Plants. 2018; 7(4):81. https://doi.org/10.3390/plants7040081
Chicago/Turabian StyleSechet, Julien, Annie Marion-Poll, and Helen M. North. 2018. "Emerging Functions for Cell Wall Polysaccharides Accumulated during Eudicot Seed Development" Plants 7, no. 4: 81. https://doi.org/10.3390/plants7040081
APA StyleSechet, J., Marion-Poll, A., & North, H. M. (2018). Emerging Functions for Cell Wall Polysaccharides Accumulated during Eudicot Seed Development. Plants, 7(4), 81. https://doi.org/10.3390/plants7040081