
Assessing the Impact of Environmental and Management Variables on Mountain Meadow Yield and Feed Quality Using a Random Forest Model


Abstract
1. Introduction
2. Results
2.1. Data Results
2.2. Random Forest Models
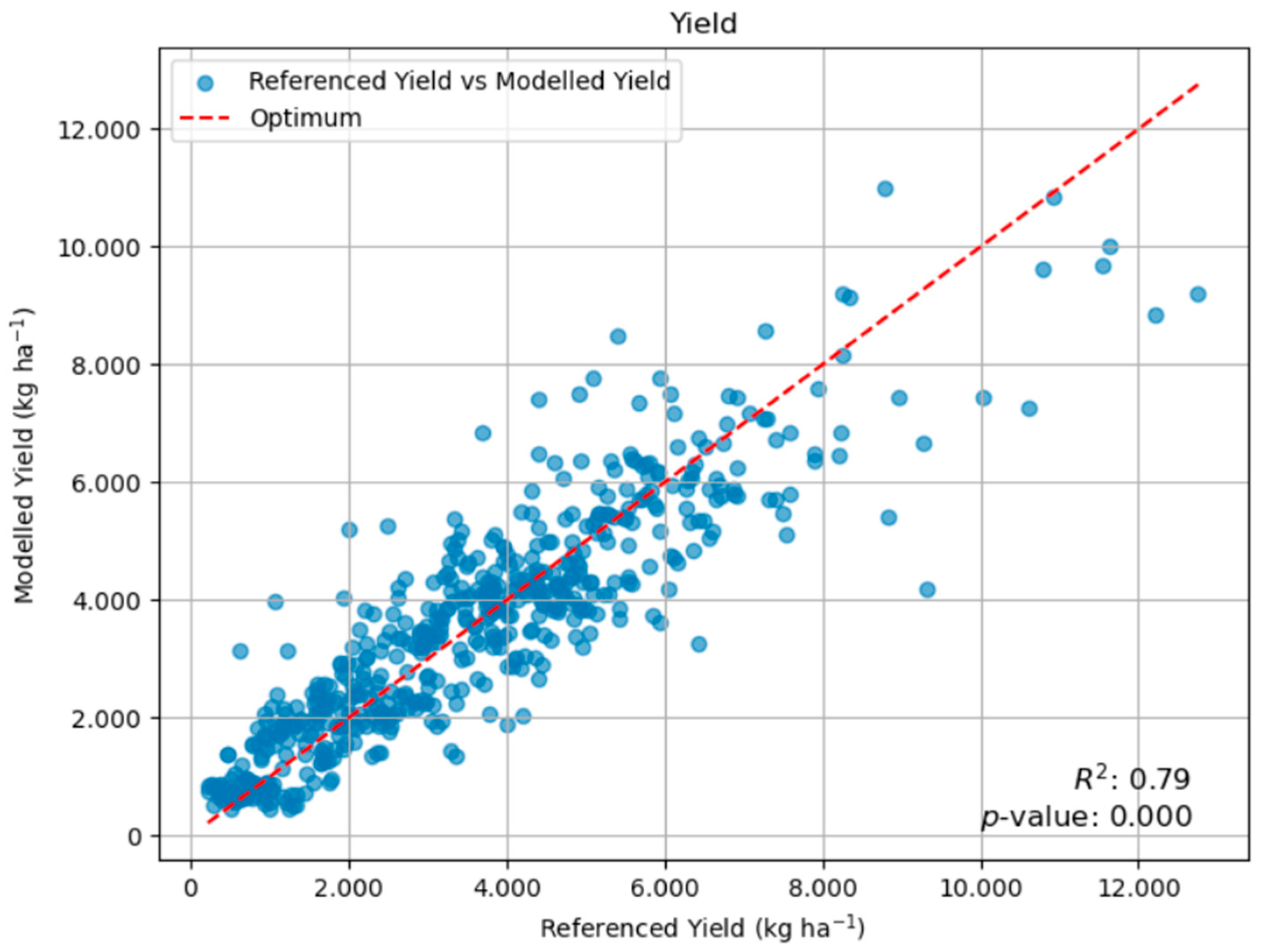
2.2.1. Yield Model
2.2.2. Protein Model
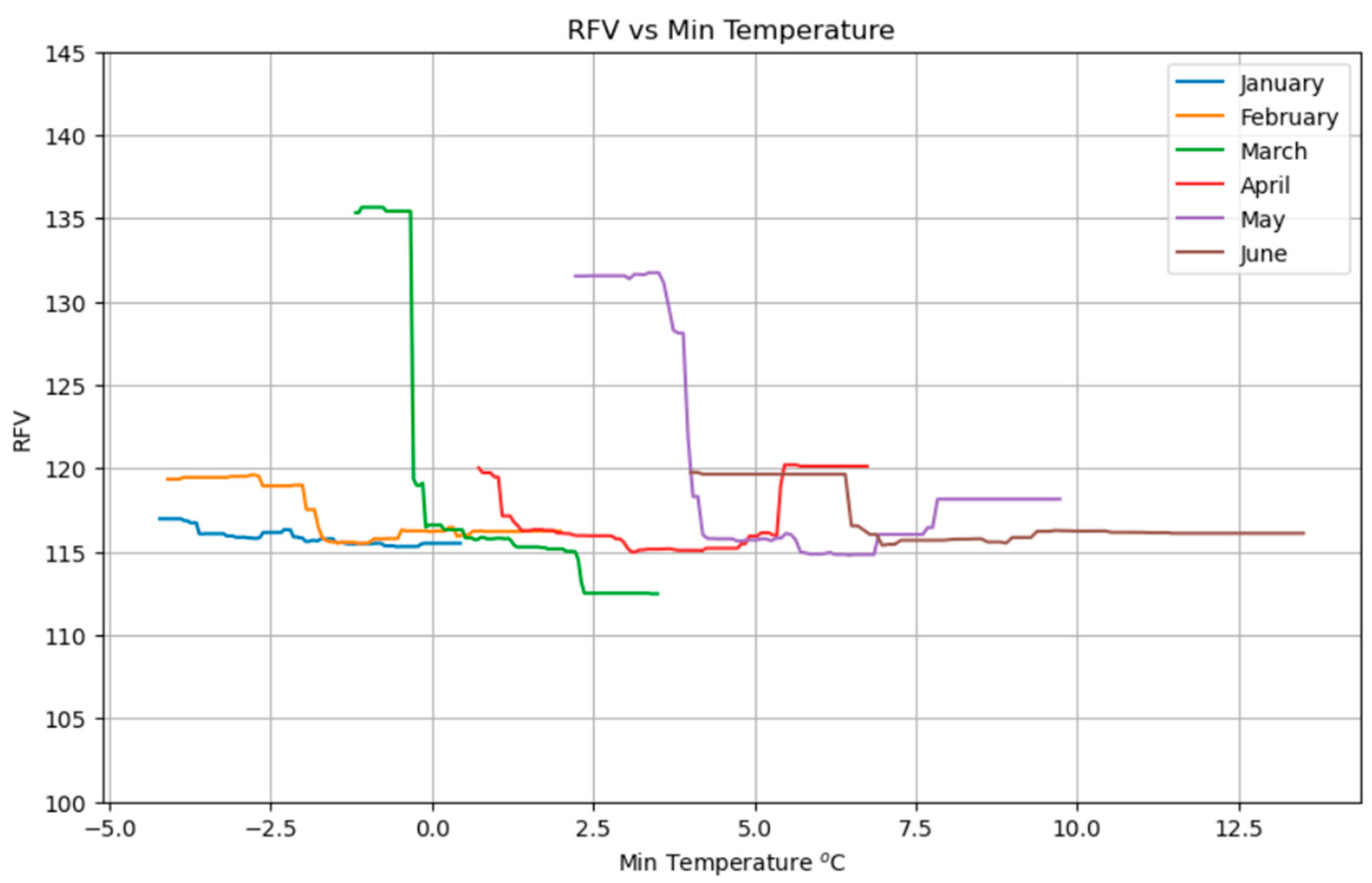
2.2.3. RFV Model
2.3. Variable Impact
2.3.1. Yield Model Variable Impact
2.3.2. Protein Model Variable Impact
2.3.3. RFV Model Variable Impact
3. Discussion
4. Materials and Methods
4.1. Study Area and Experimental Design
4.2. Farmer Surveys
4.3. Vegetation Sampling
4.4. Climatic Data Acquisition
4.5. Flora Inventory and Biodiversity Metrics
4.6. Meadow Type
4.7. Biomass Production and Dry Matter Determination
4.8. Forage Quality Analysis
4.9. Statistical Analysis and Modeling
- -
- Yield model: n_estimators = 158, max_depth = 14, min_samples_split = 5, min_samples_leaf = 1, random_state = 42
- -
- Protein model: n_estimators = 222, max_depth = 13, min_samples_split = 6, min_samples_leaf = 1, random_state = 42
- -
- RFV model: n_estimators = 198, max_depth = 27, min_samples_split = 5, min_samples_leaf = 1, random_state = 42
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| RFV | Relative Feed Value |
| LU | Livestock Units |
| RMSE | Root Mean Square Error |
| DM | Dry Matter |
| CP | Crude Protein |
| ADF | Acid Detergent Fiber |
| NDF | Neutral Detergent Fiber |
References
- Mountain Meadow Management Boosts Resilience to Climate Extremes—Innovations Report. Available online: https://www.innovations-report.com/agriculture-environment/ecology-the-environment-and-conservation/management-of-mountain-meadows-influences-resilience-to-climate-extremes/ (accessed on 15 May 2025).
- Álvarez, J.; Afif, E.; Díaz, T.E.; García, L.; Oliveira, J.A. Effects of Management Practices on Soil Properties and Plant Nutrition in Hay Meadows in Picos de Europa. Environments 2021, 8, 38. [Google Scholar] [CrossRef]
- Chocarro, C.; Reiné, R.J. El cultivo de los prados en el Pirineo. In Pastos del Pirineo; Consejo Superior de Investigaciones Científicas, CSIC: Madrid, Spain, 2008; pp. 141–158. ISBN 978-84-00-08614-5. [Google Scholar]
- Boob, M.; Truckses, B.; Seither, M.; Elsäßer, M.; Thumm, U.; Lewandowski, I. Management Effects on Botanical Composition of Species-Rich Meadows within the Natura 2000 Network. Biodivers. Conserv. 2019, 28, 729–750. [Google Scholar] [CrossRef]
- Dengler, J.; Janišová, M.; Török, P.; Wellstein, C. Biodiversity of Palaearctic Grasslands: A Synthesis. Agric. Ecosyst. Environ. 2014, 182, 1–14. [Google Scholar] [CrossRef]
- Humbert, J.-Y.; Pellet, J.; Buri, P.; Arlettaz, R. Does Delaying the First Mowing Date Benefit Biodiversity in Meadowland? Env. Evid. 2012, 1, 9. [Google Scholar] [CrossRef]
- Ascaso, J.; Reiné, R.; Barrantes, O. Evolution of Hay Meadows between 1956, 1986, and 2016 and Its Relation to the Characteristics and Location of the Parcels in the Valley of the River Esera (Pyrenees, Spain). Agronomy 2020, 10, 329. [Google Scholar] [CrossRef]
- Bowskill, V.; Bhagwat, S.; Gowing, D. Depleting Soil Nutrients through Frequency and Timing of Hay Cutting on Floodplain Meadows for Habitat Restoration and Nutrient Neutrality. Biol. Conserv. 2023, 283, 110140. [Google Scholar] [CrossRef]
- Fu, L.; Bo, T.; Du, G.; Zheng, X. Modeling the Responses of Grassland Vegetation Coverage to Grazing Disturbance in an Alpine Meadow. Ecol. Model. 2012, 247, 221–232. [Google Scholar] [CrossRef]
- Bagcilar, S.H.; Reed, C.C.; Poulson, S.R.; Verburg, P.S.J.; Sullivan, B.W. Does Montane Meadow Restoration Influence the Mineral Association and Stability of Soil Carbon? Biogeochemistry 2024, 167, 1089–1105. [Google Scholar] [CrossRef]
- Blackburn, D.A.; Oliphant, A.J.; Davis, J.D. Carbon and Water Exchanges in a Mountain Meadow Ecosystem, Sierra Nevada, California. Wetlands 2021, 41, 39. [Google Scholar] [CrossRef]
- Guasconi, D. The Hidden Half of the Meadow: Interactions Between Drought, Soil Carbon, Roots and Soil Microbial Communities. Doctoral Dissertation, Department of Physical Geography, Stockholm University, Stockholm, Sweden, 2024. [Google Scholar]
- Ascaso, J.; Reiné, R. Temporal Variations in the Production—Quality and Optimal Cutting Date of Hay Meadows in the Central Pyrenees (Spain). Agronomy 2022, 12, 918. [Google Scholar] [CrossRef]
- Reiné, R.; Barrantes, O.; Chocarro, C.; Juárez-Escario, A.; Broca, A.; Maestro, M.; Ferrer, C. Pyrenean Meadows in Natura 2000 Network: Grass Production and Plant Biodiversity Conservation. Span. J. Agric. Res. 2014, 12, 61–77. [Google Scholar] [CrossRef]
- Jones, J.A.; Hutchinson, R.; Moldenke, A.; Pfeiffer, V.; Helderop, E.; Thomas, E.; Griffin, J.; Reinholtz, A. Landscape Patterns and Diversity of Meadow Plants and Flower-Visitors in a Mountain Landscape. Landsc. Ecol. 2019, 34, 997–1014. [Google Scholar] [CrossRef]
- Gabrovska-Evstatieva, K.; Evstatiev, B. Overview of Methods and Technologies Used for Smart Management of Pastures and Meadows. AIP Conf. Proc. 2022, 2570, 040011. [Google Scholar] [CrossRef]
- Barrachina, M.; Cristóbal, J.; Tulla, A.F. Estimating Above-Ground Biomass on Mountain Meadows and Pastures through Remote Sensing. Int. J. Appl. Earth Obs. Geoinf. 2015, 38, 184–192. [Google Scholar] [CrossRef]
- Baghdadi, N.; El Hajj, M.; Zribi, M.; Bousbih, S. Calibration of the Water Cloud Model at C-Band for Winter Crop Fields and Grasslands. Remote Sens. 2017, 9, 969. [Google Scholar] [CrossRef]
- Ben Touhami, H.; Lardy, R.; Barra, V.; Bellocchi, G. Screening Parameters in the Pasture Simulation Model Using the Morris Method. Ecol. Model. 2013, 266, 42–57. [Google Scholar] [CrossRef]
- Aggarwal, P.K.; Kalra, N. Analyzing the Limitations Set by Climatic Factors, Genotype, and Water and Nitrogen Availability on Productivity of Wheat II. Climatically Potential Yields and Management Strategies. Field Crops Res. 1994, 38, 93–103. [Google Scholar] [CrossRef]
- Jing, Q.; Jégo, G.; Bélanger, G.; Chantigny, M.H.; Rochette, P. Simulation of Water and Nitrogen Balances in a Perennial Forage System Using the STICS Model. Field Crops Res. 2017, 201, 10–18. [Google Scholar] [CrossRef]
- Best, E.P.H.; Jacobs, F.H.H. Production, Nutrient Availability, and Elemental Balances of Two Meadows Affected by Different Fertilization and Water Table Regimes in The Netherlands. Plant Ecol. 2001, 155, 61–73. [Google Scholar] [CrossRef]
- Campdelacreu Rocabruna, P.; Domene, X.; Matteazzi, A.; Figl, U.; Fundneider, A.; Fernández-Martínez, M.; Venir, E.; Robatscher, P.; Preece, C.; Peñuelas, J.; et al. Effect of Organic Fertilisation on Soil Phosphatase Activity, Phosphorus Availability and Forage Yield in Mountain Permanent Meadows. Agric. Ecosyst. Environ. 2024, 368, 109006. [Google Scholar] [CrossRef]
- Comakli, B.; Mentese, O.; Koc, A. Nitrogen Fertilizing and Pre-Anthesis Cutting Stage Improve Dry Matter Production, Protein Content and Botanical Composition in Meadows. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2005, 55, 125–130. [Google Scholar] [CrossRef]
- Dolezal, J.; Lanta, V.; Mudrak, O.; Leps, J. Seasonality Promotes Grassland Diversity: Interactions with Mowing, Fertilization and Removal of Dominant Species. J. Ecol. 2019, 107, 203–215. [Google Scholar] [CrossRef]
- Wezel, A.; Stöckli, S.; Tasser, E.; Nitsch, H.; Vincent, A. Good Pastures, Good Meadows: Mountain Farmers’ Assessment, Perceptions on Ecosystem Services, and Proposals for Biodiversity Management. Sustainability 2021, 13, 5609. [Google Scholar] [CrossRef]
- Jarne, A.; Usón, A.; Reiné, R. Predictive Production Models for Mountain Meadows: A Review. Agronomy 2024, 14, 830. [Google Scholar] [CrossRef]
- Bala, A. Machine Learning Approaches for Crop Yield Prediction-Review. Int. J. Comput. Eng. Technol. 2020, 11, 23–27. [Google Scholar]
- De Rosa, D.; Basso, B.; Fasiolo, M.; Friedl, J.; Fulkerson, B.; Grace, P.R.; Rowlings, D.W. Predicting Pasture Biomass Using a Statistical Model and Machine Learning Algorithm Implemented with Remotely Sensed Imagery. Comput. Electron. Agric. 2021, 180, 105880. [Google Scholar] [CrossRef]
- Sanchez-Ruiz, S.; Chiesi, M.; Fibbi, L.; Carrara, A.; Maselli, F.; Gilabert, M.A. Optimized Application of Biome-BGC for Modeling the Daily GPP of Natural Vegetation over Peninsular Spain. J. Geophys. Res. Biogeosciences 2018, 123, 531–546. [Google Scholar] [CrossRef]
- Wätzold, F.; Drechsler, M.; Johst, K.; Mewes, M.; Sturm, A. A Novel, Spatiotemporally Explicit Ecological-Economic Modeling Procedure for the Design of Cost-Effective Agri-Environment Schemes to Conserve Biodiversity. Am. J. Agric. Econ. 2016, 98, 489–512. [Google Scholar] [CrossRef]
- Pruchniewicz, D. Abandonment of Traditionally Managed Mesic Mountain Meadows Affects Plant Species Composition and Diversity. Basic Appl. Ecol. 2017, 20, 10–18. [Google Scholar] [CrossRef]
- Bogireddy, S.R.; Murari, H. Enhancing Crop Yield Prediction through Random Forest Classifier: A Comprehensive Approach. In Proceedings of the 2024 5th International Conference on Smart Electronics and Communication (ICOSEC), Tiruchirappalli, India, 18–20 September 2024; pp. 1663–1668. [Google Scholar]
- Liakos, K.G.; Busato, P.; Moshou, D.; Pearson, S.; Bochtis, D. Machine Learning in Agriculture: A Review. Sensors 2018, 18, 2674. [Google Scholar] [CrossRef]
- Serpa-Imbett, C.M.; Gómez-Palencia, E.L.; Medina-Herrera, D.A.; Mejía-Luquez, J.A.; Martínez, R.R.; Burgos-Paz, W.O.; Aguayo-Ulloa, L.A. In-Field Forage Biomass and Quality Prediction Using Image and VIS-NIR Proximal Sensing with Machine Learning and Covariance-Based Strategies for Livestock Management in Silvopastoral Systems. AgriEngineering 2025, 7, 111. [Google Scholar] [CrossRef]
- Sahu, S.; Chawla, M.; Khare, N. An Efficient Analysis of Crop Yield Prediction Using Hadoop Framework Based on Random Forest Approach. In Proceedings of the 2017 International Conference on Computing, Communication and Automation (ICCCA), Greater Noida, India, 5–6 May 2017; pp. 53–57. [Google Scholar]
- Elavarasan, D.; Vincent, P.M.D.R. A Reinforced Random Forest Model for Enhanced Crop Yield Prediction by Integrating Agrarian Parameters. J. Ambient. Intell. Hum. Comput. 2021, 12, 10009–10022. [Google Scholar] [CrossRef]
- Burdett, H.; Wellen, C. Statistical and Machine Learning Methods for Crop Yield Prediction in the Context of Precision Agriculture. Precis. Agric. 2022, 23, 1553–1574. [Google Scholar] [CrossRef]
- Kroes, J.G.; Supit, I. Impact Analysis of Drought, Water Excess and Salinity on Grass Production in The Netherlands Using Historical and Future Climate Data. Agric. Ecosyst. Environ. 2011, 144, 370–381. [Google Scholar] [CrossRef]
- Ludewig, K.; Donath, T.W.; Zelle, B.; Eckstein, R.L.; Mosner, E.; Otte, A.; Jensen, K. Effects of Reduced Summer Precipitation on Productivity and Forage Quality of Floodplain Meadows at the Elbe and the Rhine River. PLoS ONE 2015, 10, e0124140. [Google Scholar] [CrossRef]
- Fiorelli, J.-L.; Drouet, J.-L.; Duretz, S.; Gabrielle, B.; Cellier, P.; Blanfort, V.; Capitaine, M.; Boisdon, I.; Soussana, J.-F. A Whole Farm Approach Based on Farm Management Practices Data Coupled with Crop and Grassland Mechanistic Models. In Proceedings of the Colloque International Scientifique et Professionnel: Agriculture Biologique et Changement Climatique, Clermont Ferrand, France, 17–18 April 2008. [Google Scholar]
- Aghajanzadeh-Darzi, P.; Martin, R.; Laperche, S.; Jayet, P.-A. Climate Change Impacts on European Agriculture Revisited: Adding the Economic Dimension of Grasslands. Reg. Environ. Chang. 2017, 17, 261–272. [Google Scholar] [CrossRef]
- Optimal Base Temperature for Computing Growing Degree-Day Sums to Predict Forage Quality of Mountain Permanent Meadow in South Tyrol. Available online: https://bia.unibz.it/esploro/outputs/journalArticle/Optimal-base-temperature-for-computing-growing/991006484894901241 (accessed on 20 May 2025).
- Graux, A.-I.; Bellocchi, G.; Lardy, R.; Soussana, J.-F. Ensemble Modelling of Climate Change Risks and Opportunities for Managed Grasslands in France. Agric. For. Meteorol. 2013, 170, 114–131. [Google Scholar] [CrossRef]
- Duffkova, R.; Kvívtek, T. Effect of Cutting Regime on Soil Physical Properties of Wet Thistle Meadows. Soil Water Res. 2009, 4, 104–115. [Google Scholar] [CrossRef]
- Radkowski, A.; Radkowska, I.; Godyn, D. Effects of Fertilization with an Amino Acid Preparation on the Dry Matter Yield and Chemical Composition of Meadow Plants. J. Elem. 2018, 23, 947–958. [Google Scholar] [CrossRef]
- Reiné, R.; Ascaso, J.; Barrantes, O. Nutritional Quality of Plant Species in Pyrenean Hay Meadows of High Diversity. Agronomy 2020, 10, 883. [Google Scholar] [CrossRef]
- Stewart, G.B.; Pullin, A.S. The Relative Importance of Grazing Stock Type and Grazing Intensity for Conservation of Mesotrophic ‘Old Meadow’ Pasture. J. Nat. Conserv. 2008, 16, 175–185. [Google Scholar] [CrossRef]
- Donovan, M.; Monaghan, R. Impacts of Grazing on Ground Cover, Soil Physical Properties and Soil Loss via Surface Erosion: A Novel Geospatial Modelling Approach. J. Environ. Manag. 2021, 287, 112206. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Gong, Y.; Li, Y.; Liu, S.; Yu, Z.; Zhao, R. Does Grazing Exclusion Enhance Grassland Restoration? Evidence from Northern China. Ecol. Indic. 2023, 149, 110166. [Google Scholar] [CrossRef]
- Wang, S.; Duan, J.; Xu, G.; Wang, Y.; Zhang, Z.; Rui, Y.; Luo, C.; Xu, B.; Zhu, X.; Chang, X.; et al. Effects of Warming and Grazing on Soil N Availability, Species Composition, and ANPP in an Alpine Meadow. Ecology 2012, 93, 2365–2376. [Google Scholar] [CrossRef]
- Oles, K.M.; Weixelman, D.A.; Lile, D.F.; Tate, K.W.; Snell, L.K.; Roche, L.M. Riparian Meadow Response to Modern Conservation Grazing Management. Environ. Manag. 2017, 60, 383–395. [Google Scholar] [CrossRef]
- Pavlů, V.; Hejcman, M.; Pavlů, L.; Gaisler, J.; Nežerková, P. Effect of Continuous Grazing on Forage Quality, Quantity and Animal Performance. Agric. Ecosyst. Environ. 2006, 113, 349–355. [Google Scholar] [CrossRef]
- Follett, R.H.; Westfall, D.G.; Shanahan, J.F.; Lybecker, D.W. Nitrogen Fertilization of Mountain Meadows. J. Prod. Agric. 1995, 8, 239–243. [Google Scholar] [CrossRef]
- Gasarch, E.I.; Seastedt, T.R. Plant Community Response to Nitrogen and Phosphorus Enrichment Varies across an Alpine Tundra Moisture Gradient. Plant Ecol. Divers. 2015, 8, 739–749. [Google Scholar] [CrossRef]
- Tan, S.S.; Smeins, F.E. Predicting Grassland Community Changes with an Artificial Neural Network Model. Ecol. Model. 1996, 84, 91–97. [Google Scholar] [CrossRef]
- Linn, J.G.; Martin, N.P. Forage Quality Tests and Interpretation (Revised 1989); University of Minnesota. Agricultural Extension Service: St. Paul, MN, USA, 1989. [Google Scholar]
- JupyterLab Documentation—JupyterLab 4.5.0a0 Documentation. Available online: https://jupyterlab.readthedocs.io/en/latest/ (accessed on 25 May 2025).
- Scikit-Learn: Machine Learning in Python—Scikit-Learn 1.6.1 Documentation. Available online: https://scikit-learn.org/stable/ (accessed on 25 May 2025).
- Botero-Valencia, J.; García-Pineda, V.; Valencia-Arias, A.; Valencia, J.; Reyes-Vera, E.; Mejia-Herrera, M.; Hernández-García, R. Machine Learning in Sustainable Agriculture: Systematic Review and Research Perspectives. Agriculture 2025, 15, 377. [Google Scholar] [CrossRef]
- Pugh, N.A.; Young, A.; Ojha, M.; Emendack, Y.; Sanchez, J.; Xin, Z.; Puppala, N. Yield Prediction in a Peanut Breeding Program Using Remote Sensing Data and Machine Learning Algorithms. Front. Plant Sci. 2024, 15, 1339864. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2019 | 2020 | 2022 | 2023 | 2024 | ||
|---|---|---|---|---|---|---|
| January | Average temperature °C | 2.89 | 3.58 | 3.61 | 1.64 | 3.92 |
| Maximum temperature °C | 7.38 | 8.22 | 9.60 | 6.80 | 8.74 | |
| Minimum temperature °C | −1.63 | −1.06 | −2.37 | −2.42 | −0.86 | |
| Cumulated rain mm | 78.15 | 95.92 | 33.87 | 93.06 | 64.96 | |
| February | Average temperature °C | 5.40 | 6.35 | 5.64 | 3.43 | 5.59 |
| Maximum temperature °C | 12.04 | 12.56 | 11.69 | 9.47 | 10.69 | |
| Minimum temperature °C | −1.24 | 0.19 | −0.41 | −2.59 | 0.47 | |
| Rain mm | 48.00 | 5.42 | 30.76 | 42.38 | 108.14 | |
| March | Average temperature °C | 6.95 | 6.16 | 5.74 | 7.33 | 6.25 |
| Maximum temperature °C | 13.46 | 10.72 | 9.67 | 13.46 | 11.44 | |
| Minimum temperature °C | 0.47 | 1.61 | 1.81 | 1.22 | 1.08 | |
| Rain mm | 14.77 | 176.71 | 81.60 | 25.18 | 228.37 | |
| April | Average temperature °C | 7.48 | 8.72 | 7.78 | 9.60 | 8.40 |
| Maximum temperature °C | 12.50 | 12.82 | 13.30 | 16.02 | 14.26 | |
| Minimum temperature °C | 2.49 | 4.63 | 2.25 | 3.20 | 2.57 | |
| Rain mm | 157.89 | 118.72 | 111.70 | 20.78 | 60.23 | |
| May | Average temperature °C | 10.12 | 12.68 | 13.24 | 11.16 | 10.36 |
| Maximum temperature °C | 16.16 | 18.43 | 19.56 | 17.15 | 15.99 | |
| Minimum temperature °C | 4.06 | 6.94 | 6.92 | 5.13 | 4.78 | |
| Rain mm | 87.43 | 123.40 | 31.31 | 70.61 | 100.35 | |
| June | Average temperature °C | 15.07 | 12.91 | 16.91 | 14.79 | 14.27 |
| Maximum temperature °C | 21.78 | 18.13 | 23.54 | 20.18 | 20.46 | |
| Minimum temperature °C | 8.36 | 7.65 | 10.29 | 9.40 | 8.10 | |
| Rain mm | 0.49 | 137.36 | 65.91 | 174.22 | 91.70 | |
| Annual fertilization | N kg ha−1 | 95.20 | 101.24 | 99.59 | 91.57 | 91.57 |
| P kg ha−1 | 114.72 | 137.46 | 125.47 | 116.15 | 116.15 | |
| K kg ha−1 | 71.74 | 73.68 | 77.31 | 70.05 | 70.05 | |
| Annual stocking rate | Livestock Units (LUs ha−1) | 0.38 | 0.34 | 0.36 | 0.40 | 0.40 |
| Cutting date | Day of year | 178.62 | 160.17 | 156.52 | 153.71 | 155.79 |
| Yield | kg ha−1 | 1997.31 | 4862.54 | 4684.55 | 2792.58 | 6273.54 |
| Crude protein | % | 12.05 | 10.24 | 11.14 | 14.08 | 11.94 |
| RFV | RFV units | 123.42 | 107.12 | 112.71 | 126.91 | 115.28 |
| Yield | Protein | RFV | ||
|---|---|---|---|---|
| January | Average temperature °C | 0.006 | 0.033 | 0.052 |
| Maximum temperature °C | 0.018 | 0.018 | 0.028 | |
| Minimum temperature °C | 0.011 | 0.012 | 0.011 | |
| Rain mm | 0.013 | 0.013 | 0.008 | |
| February | Average temperature °C | 0.007 | 0.020 | 0.025 |
| Maximum temperature °C | 0.008 | 0.011 | 0.020 | |
| Minimum temperature °C | 0.037 | 0.021 | 0.029 | |
| Rain mm | 0.016 | 0.045 | 0.008 | |
| March | Average temperature °C | 0.007 | 0.023 | 0.008 |
| Maximum temperature °C | 0.041 | 0.009 | 0.008 | |
| Minimum temperature °C | 0.012 | 0.083 | 0.134 | |
| Rain mm | 0.430 | 0.021 | 0.008 | |
| April | Average temperature °C | 0.006 | 0.012 | 0.008 |
| Maximum temperature °C | 0.015 | 0.013 | 0.018 | |
| Minimum temperature °C | 0.016 | 0.024 | 0.013 | |
| Rain mm | 0.010 | 0.021 | 0.014 | |
| May | Average temperature °C | 0.004 | 0.017 | 0.019 |
| Maximum temperature °C | 0.006 | 0.008 | 0.023 | |
| Minimum temperature °C | 0.006 | 0.053 | 0.078 | |
| Rain mm | 0.007 | 0.006 | 0.007 | |
| June | Average temperature °C | 0.010 | 0.009 | 0.013 |
| Maximum temperature °C | 0.011 | 0.019 | 0.020 | |
| Minimum temperature °C | 0.006 | 0.010 | 0.007 | |
| Rain mm | 0.007 | 0.021 | 0.005 | |
| Fertilization | N kg ha−1 | 0.005 | 0.003 | 0.011 |
| P kg ha−1 | 0.006 | 0.003 | 0.011 | |
| K kg ha−1 | 0.020 | 0.008 | 0.005 | |
| Livestock load | LUs | 0.015 | 0.054 | 0.040 |
| Cutting date | Day of year | 0.194 | 0.366 | 0.344 |
| Biodiversity | Shannon Index | 0.025 | 0.040 | 0.021 |
| Meadow type | 0.014 | 0.003 | 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jarne, A.; Usón, A.; Reiné, R. Assessing the Impact of Environmental and Management Variables on Mountain Meadow Yield and Feed Quality Using a Random Forest Model. Plants 2025, 14, 2150. https://doi.org/10.3390/plants14142150
Jarne A, Usón A, Reiné R. Assessing the Impact of Environmental and Management Variables on Mountain Meadow Yield and Feed Quality Using a Random Forest Model. Plants. 2025; 14(14):2150. https://doi.org/10.3390/plants14142150
Chicago/Turabian StyleJarne, Adrián, Asunción Usón, and Ramón Reiné. 2025. "Assessing the Impact of Environmental and Management Variables on Mountain Meadow Yield and Feed Quality Using a Random Forest Model" Plants 14, no. 14: 2150. https://doi.org/10.3390/plants14142150
APA StyleJarne, A., Usón, A., & Reiné, R. (2025). Assessing the Impact of Environmental and Management Variables on Mountain Meadow Yield and Feed Quality Using a Random Forest Model. Plants, 14(14), 2150. https://doi.org/10.3390/plants14142150