


A Morphometric Analysis of Starch Granules from Two Dioscorea Species


Abstract
1. Introduction
Study Species
2. Materials and Methods
2.1. Reference Collection
2.2. Sample Preparation
2.3. Microscopy
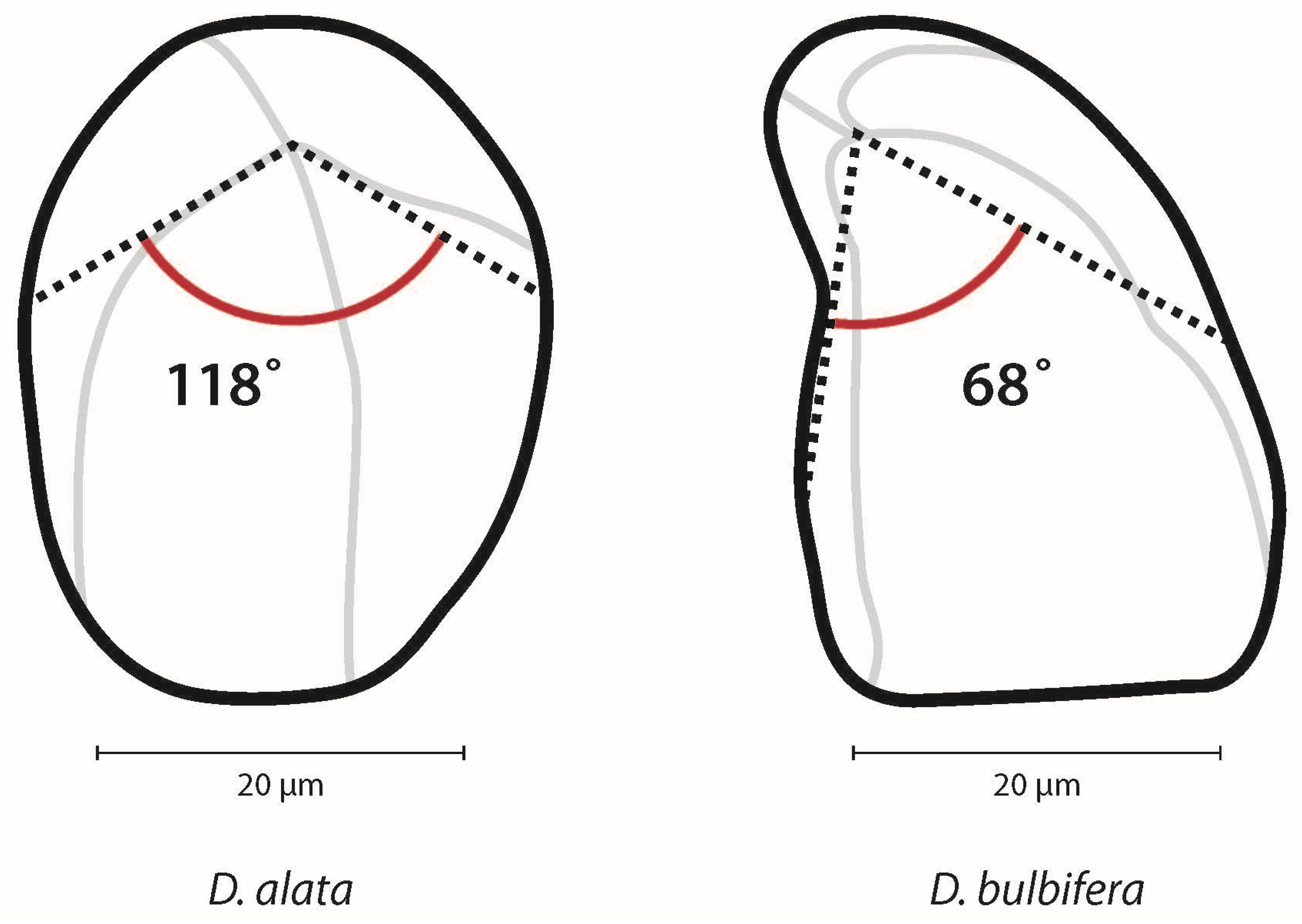
2.4. Granule Measurements and Morphology

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphological Character | Description | Image |
|---|---|---|
| Hilum | The hilum is the point of origin around which layers (lamellae) are deposited [22]. An eccentric hilum is located well outside the geometric center of the granule [21]. | 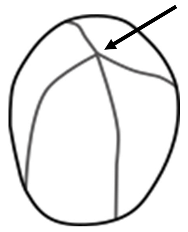 |
| Lamellae | Concentric growth rings of amylase and amylopectin emanating from the hilum [21,22]. Visible under DIC light. | 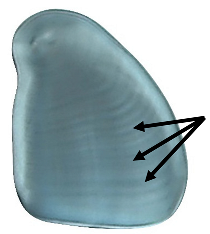 |
| Longitudinal Fissure | Interruption in the starch granule, extending along the long axis of the granule [21,22]. |  |
| Curve | A curve or smooth bend in the granule, usually occurring on the proximal end of the granule close to the hilum. | 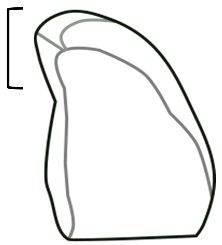 |
| Triangular | Triangular granules possess three well-defined sides [21]. |  |
| Oval | Oval granules have a rounded and slightly elongated outline [21]. |  |
3. Results
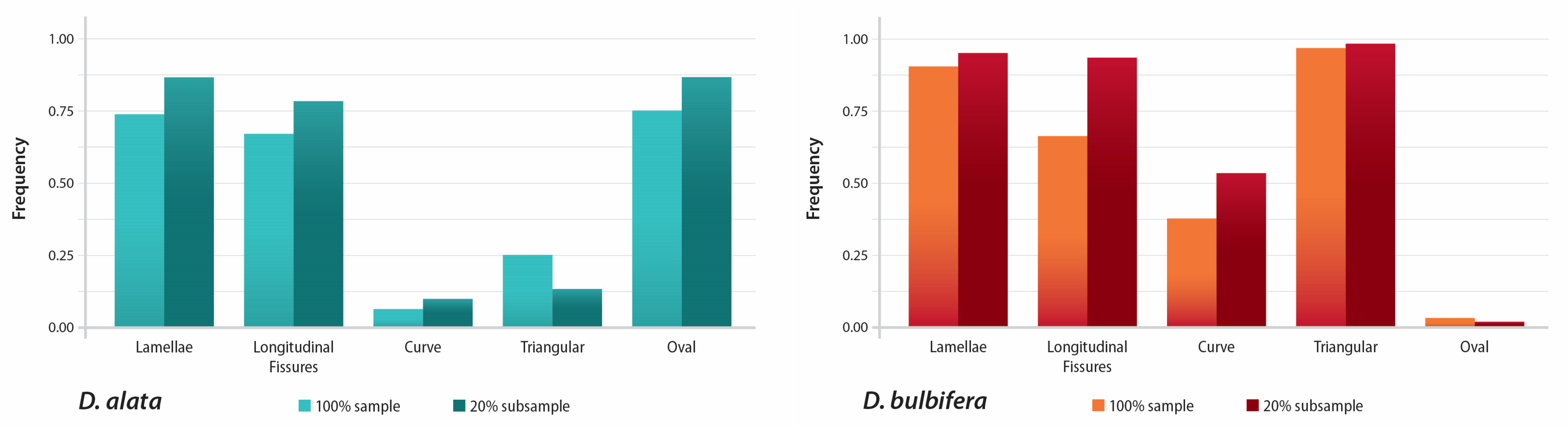
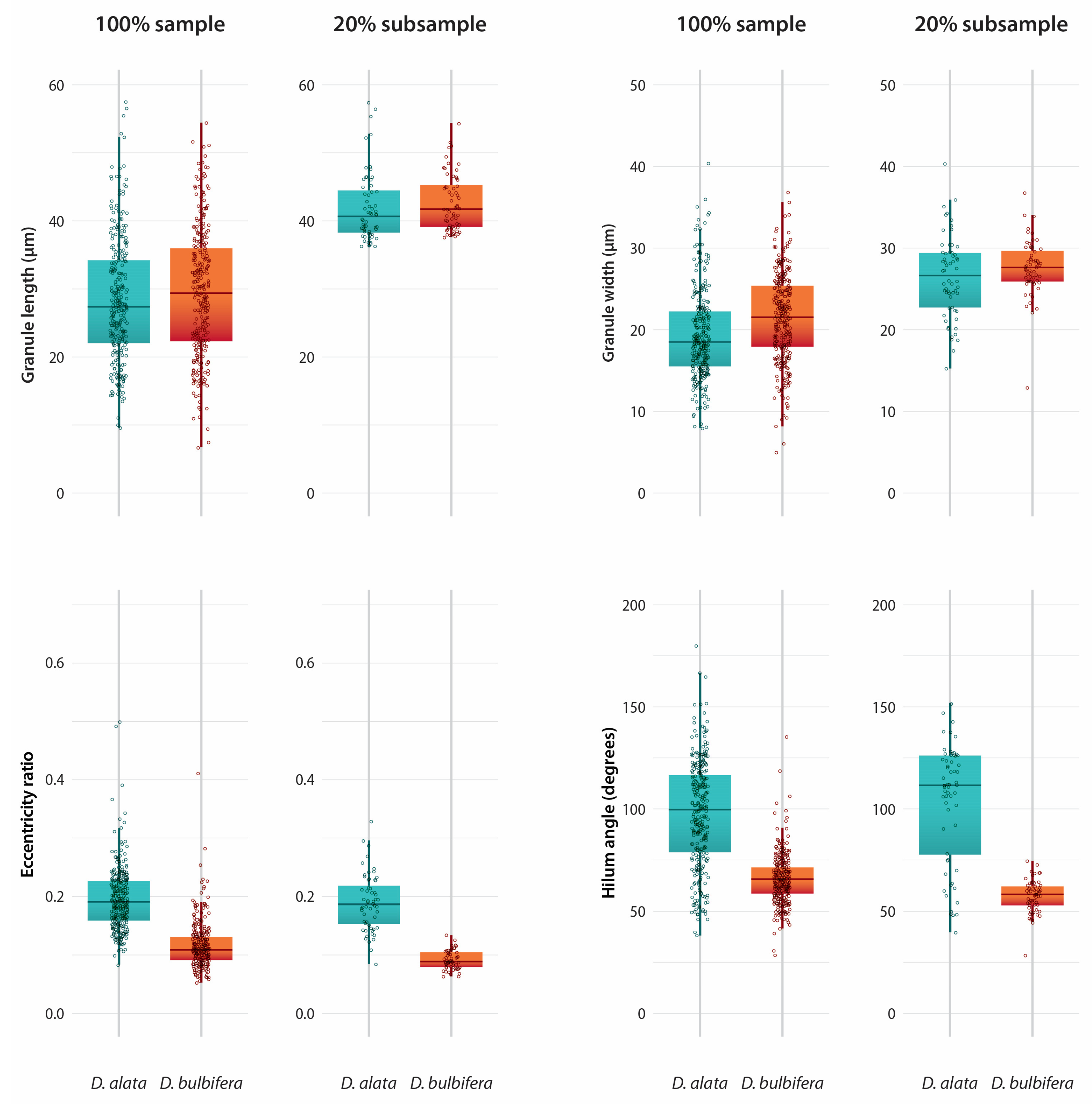
3.1. Granule Characters and Morphology
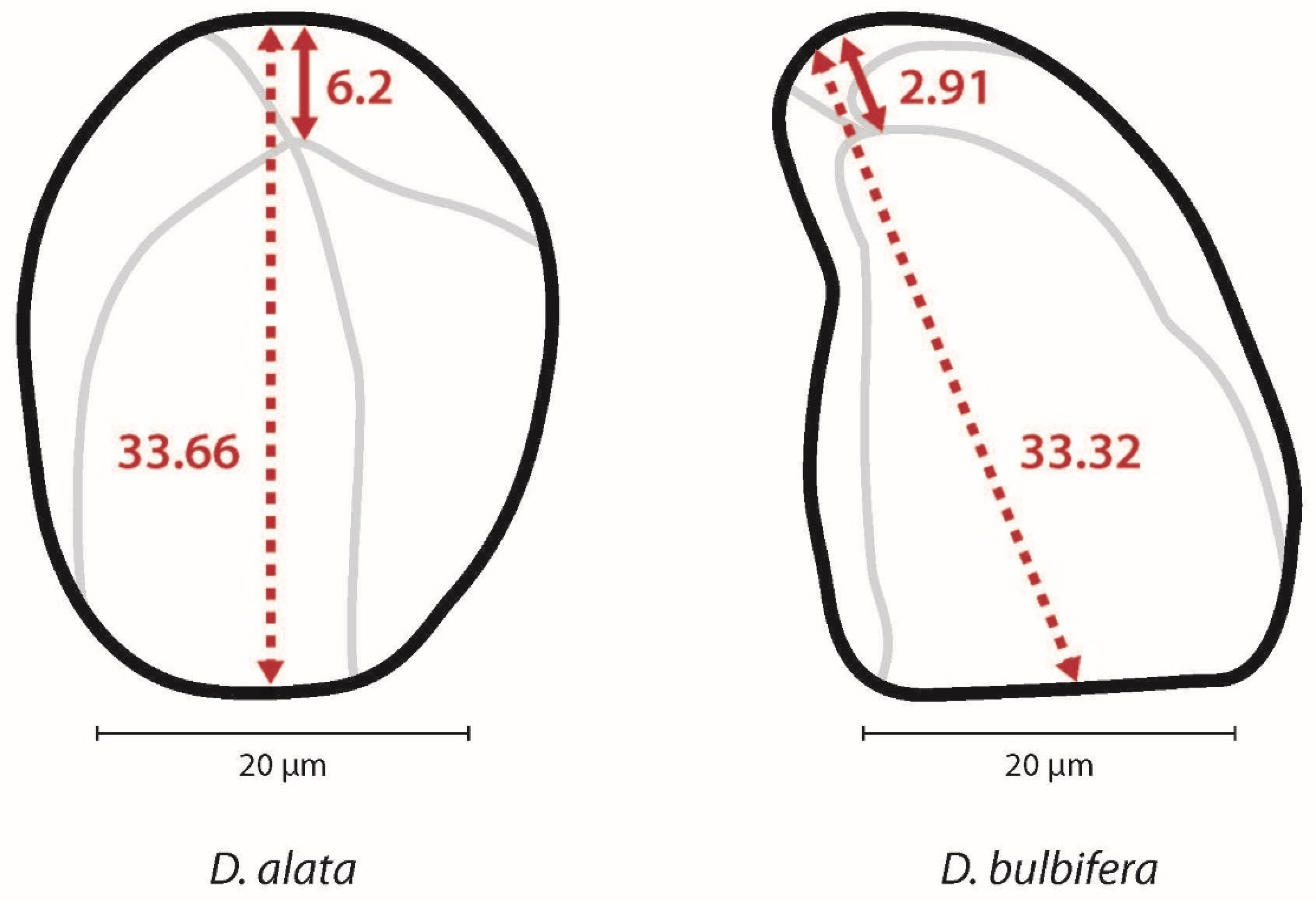
3.2. Dioscorea alata
3.3. Dioscorea bulbifera
3.4. Differences Between Species
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hawkes, K.; O’Connell, J.F.; Blurton Jones, N.G. Hadza Meat Sharing. Evol. Hum. Behav. 2001, 22, 113–142. [Google Scholar] [CrossRef] [PubMed]
- Torrence, R.; Barton, H. Ancient Starch Research, 1st ed.; Routledge: London, UK, 2006. [Google Scholar]
- Allen, M.S.; Ussher, E. Starch Analysis Reveals Prehistoric Plant Translocations and Shell Tool Use, Marquesas Islands, Polynesia. J. Archaeol. Sci. 2013, 40, 2799–2812. [Google Scholar] [CrossRef]
- Fullagar, R.; Field, J.; Denham, T.; Lentfer, C. Early and Mid Holocene Tool-Use and Processing of Taro (Colocasia esculenta), Yam (Dioscorea sp.) and Other Plants at Kuk Swamp in the Highlands of Papua New Guinea. J. Archaeol. Sci. 2006, 33, 595–614. [Google Scholar] [CrossRef]
- Ciofalo, A.J.; Keegan, W.F.; Pateman, M.P.; Pagán-Jiménez, J.R.; Hofman, C.L. Determining Precolonial Botanical Foodways: Starch Recovery and Analysis, Long Island, The Bahamas. J. Archaeol. Sci. Rep. 2018, 21, 305–317. [Google Scholar] [CrossRef]
- Coursey, D.G. The Civilizations of the Yam: Interrelationships of Man and Yams in Africa and the Indo-Pacific Region. Archaeol. Phys. Anthropol. Ocean. 1972, 7, 215–233. [Google Scholar]
- Whistler, A. The Ethnobotany of Tonga: The Plants, Their Tongan Names, and Their Uses; Bishop Museum Bulletins in Botany; Bishop Museum Press: Honolulu, HI, USA, 1991. [Google Scholar]
- Loy, T.H.; Spriggs, M.; Wickler, S. Direct Evidence for Human Use of Plants 28,000 Years Ago: Starch Residues on Stone Artefacts from the Northern Solomon Islands. Antiquity 1992, 66, 898–912. [Google Scholar] [CrossRef]
- Burley, D.V.; Horrocks, M.; Weisler, M.I. Earliest Evidence for Pit Cultivation Provides Insight on the Nature of First Polynesian Settlement. J. Isl. Coast. Archaeol. 2020, 15, 127–147. [Google Scholar] [CrossRef]
- Ussher, E.M. Agriculture in Tongan Prehistory: An Archaeobotanical Perspective. Ph.D. Thesis, Australian National University, Canberra, Australia, 2015. [Google Scholar] [CrossRef]
- Huard, K.; Burley, D. On the question of Anadara antiquata bivalve scrapers in the archaeological record of Tonga. Archaeol. Ocean. 2017, 52, 141–148. [Google Scholar] [CrossRef]
- Ngo Ngwe, M.F.S.; Omokolo, D.N.; Joly, S. Evolution and Phylogenetic Diversity of Yam Species (Dioscorea spp.): Implication for Conservation and Agricultural Practices. PLoS ONE 2015, 10, e0145364. [Google Scholar] [CrossRef] [PubMed]
- Arnau, G.; Bhattacharjee, R.; Mn, S.; Chair, H.; Malapa, R.; Lebot, V.; Abraham, K.; Perrier, X.; Petro, D.; Penet, L.; et al. Understanding the Genetic Diversity and Population Structure of Yam (Dioscorea alata L.) Using Microsatellite Markers. PLoS ONE 2017, 12, e0174150. [Google Scholar] [CrossRef] [PubMed]
- Vaioleti, T. Kahokaho: ‘Ufi from the God/s; Its Future in the Hands of People. Available online: https://www.dvv-international.de/en/adult-education-and-development/editions/aed-792012/rio-20-education-for-sustainability/kahokaho-ufi-from-the-gods-its-future-in-the-hands-of-people (accessed on 19 November 2023).
- Dioscorea Alata—Plant Finder. Available online: https://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?taxonid=279847&isprofile=0&chr=19 (accessed on 4 November 2023).
- Ferdon, E. Early Tonga: As the Explorers Saw It (1616–1810); University of Arizona Press: Tucson, AZ, USA, 1987. [Google Scholar]
- Whistler, W.A. Plants of the Canoe People: An Ethnobotanical Voyage Through Polynesia; National Tropical Botanical Garden: Kalāheo, HI, USA, 2009; ISBN 978-0-915809-00-4. [Google Scholar]
- Cook, J.; King, J. A Voyage to the Pacific Ocean. Undertaken by the Command of His Majesty, for Making Discoveries in the Northern Hemisphere, to Determine the Position and Extent of the West Side of North America; Its Distance from Asia; and the Practicability of a Northern Passage to Europe; G. Nichol & T. Cadell: London, UK, 1784. [Google Scholar]
- Mercader, J.; Abtosway, M.; Bird, R.; Bundala, M.; Clarke, S.; Favreau, J.; Inwood, J.L.; Itambu, M.; Larter, F.; Lee, P.; et al. Morphometrics of Starch Granules From Sub-Saharan Plants and the Taxonomic Identification of Ancient Starch. Front. Earth Sci. 2018, 6, 146. [Google Scholar] [CrossRef]
- Fatokun, O.T. Micrometrics and Morphological Properties of Starch. In Chemical Properties of Starch; Emeje, M., Ed.; IntechOpen: London, UK, 2020; ISBN 978-1-83880-115-1. [Google Scholar]
- ICSN 2011, The International Code for Starch Nomenclature. Available online: https://fossilfarm.org/ICSN/Code.html (accessed on 23 January 2025).
- Reichert, E.T. The Differentiation and Specificity of Starches in Relation to Genera, Species, etc.; Carnegie Institution of Washington: Washington, DC, USA, 1913. [Google Scholar]
- Louderback, L.A.; Herzog, N.M.; Pavlik, B.M. A New Approach for Identifying Starch Granules of Wild Food Plants from Arid Western North America: Identifying Starch Granules from Underground Storage Organs. Starch-Stärke 2017, 69, 1600167. [Google Scholar] [CrossRef]
- Shannon, J.C.; Garwood, D.L.; Boyer, C.D. Genetics and Physiology of Starch Development. In Starch: Chemistry and Technology; Academic Press: Cambridge, MA, USA, 2009. [Google Scholar]
- A Box and Whiskers Plot (in the Style of Tukey)—Geom_boxplot. Available online: https://ggplot2.tidyverse.org/reference/geom_boxplot.html?q=boxplot#null (accessed on 21 October 2024).
- Field, J.; Cosgrove, R.; Fullagar, R.; Lance, B. Starch Residues on Grinding Stones in Private Collections: A Study of Morahs from the Tropical Rainforests of NE Queensland. In Archaeological Science Under a Microscope: Studies in Residue and Ancient DNA Analysis in Honour of Thomas H. Loy; Haslam, M., Robertson, G., Crowther, A., Nugent, S., Kirkwood, L., Eds.; ANU Press: Canberra, Australia, 2009; ISBN 978-1-921536-84-7. [Google Scholar]
- Berenguer, P.; Clavero, C.; Saldarriaga-Córdoba, M.; Rivera-Hutinel, A.; Seelenfreund, D.; Martinsson-Wallin, H.; Castañeda, P.; Seelenfreund, A. Identification of Breadfruit (Artocarpus altilis) and South American Crops Introduced during Early Settlement of Rapa Nui (Easter Island), as Revealed through Starch Analysis. PLoS ONE 2024, 19, e0298896. [Google Scholar] [CrossRef] [PubMed]
- Field, J.H.; Coster, A.C.; Shaw, B.; Hayes, E.; Fullagar, R.; Lovave, M.; Haro, J.; Summerhayes, G.R. Ancient Starch and Usewear Analyses of an Excavated Pestle Fragment from the Upper Kaironk Valley, Madang Province, Papua New Guinea. In Forty Years in the South Seas: Archaeological Perspectives on the Human History of Papua New Guinea and the Western Pacific Region; Ford, A., Shaw, B., Gaffney, D., Eds.; ANU Press: Canberra, Australia, 2024; pp. 391–415. ISBN 978-1-76046-643-5. [Google Scholar]
- Bascom, W.R. Ponapean Prestige Economy. Southwest. J. Anthropol. 1948, 4, 211–221. [Google Scholar] [CrossRef]
- Padhan, B.; Panda, D. Potential of Neglected and Underutilized Yams (Dioscorea spp.) for Improving Nutritional Security and Health Benefits. Front. Pharmacol. 2020, 11, 496. [Google Scholar] [CrossRef] [PubMed]




| Lab Sample Number | Species | Variety/Tongan Name | Island | Context |
|---|---|---|---|---|
| LS1 | D. alata | ‘Ufi Sikalu | Vava’u | Community Garden |
| LS2 | D. alata | ‘Ufi Hawaii (purple var.) | Vava’u | Private plantation |
| LS3 | D. alata | ‘Ufi Hawaii (white var.) | Vava’u | Private plantation |
| LS5 | D. bulbifera | Hoi | Vava’u | Roadside, fallow field |
| LS6 | D. bulbifera | Hoi | Vava’u | Fallow field |
| LS7 | D. bulbifera | Hoi | Vava’u | Roadside |
| Plant Taxon | Granule Length | Eccentricity Ratio (Range) | Angle (Range) | Shape | Lamellae | Longitudinal Fissure | Curved at Proximal End |
|---|---|---|---|---|---|---|---|
| D. alata (100%) (n = 298) | 9.53–57.46 µm | 0.08–0.50 | 38–180° | Oval 75% | 73% | 67% | 6% |
| D. alata (20% subsample) (n = 60) | 36.35–57.46 µm | 0.08–0.33 | 40–152° | Oval 86% | 86% | 78% | 10% |
| D. bulbifera (100%) (n = 301) | 6.65–54.42 µm | 0.05–0.41 | 28–135° | Triangular 96% | 90% | 93% | 37% |
| D. bulbifera (20% subsample) (n = 60) | 37.62–54.42 µm | 0.06–0.13 | 28–75° | Triangular 98% | 95% | 98% | 53% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rickett, S.; Louderback, L.A.; Bell, A.V. A Morphometric Analysis of Starch Granules from Two Dioscorea Species. Plants 2025, 14, 1869. https://doi.org/10.3390/plants14121869
Rickett S, Louderback LA, Bell AV. A Morphometric Analysis of Starch Granules from Two Dioscorea Species. Plants. 2025; 14(12):1869. https://doi.org/10.3390/plants14121869
Chicago/Turabian StyleRickett, Sara, Lisbeth A. Louderback, and Adrian V. Bell. 2025. "A Morphometric Analysis of Starch Granules from Two Dioscorea Species" Plants 14, no. 12: 1869. https://doi.org/10.3390/plants14121869
APA StyleRickett, S., Louderback, L. A., & Bell, A. V. (2025). A Morphometric Analysis of Starch Granules from Two Dioscorea Species. Plants, 14(12), 1869. https://doi.org/10.3390/plants14121869