
Overexpression of Peony PoWOX1 Promotes Callus Induction and Root Development in Arabidopsis thaliana

 , ,
, ,
Abstract
1. Introduction
2. Results
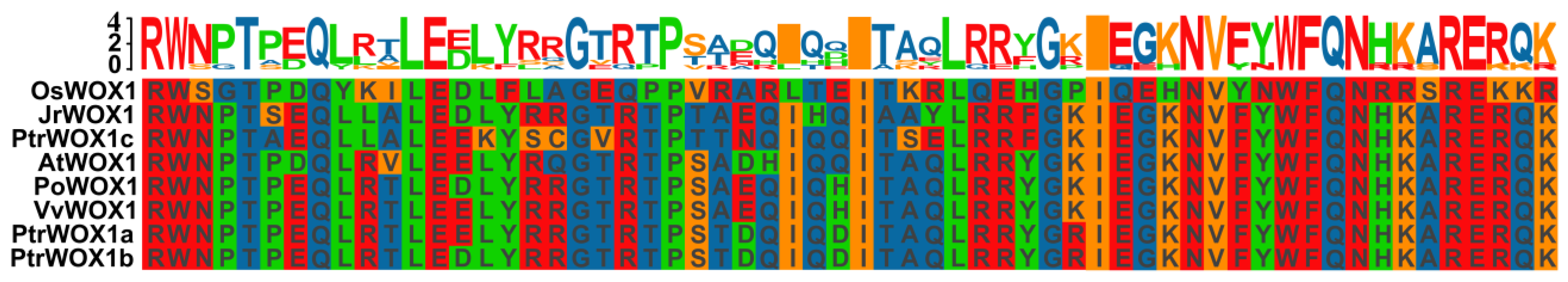
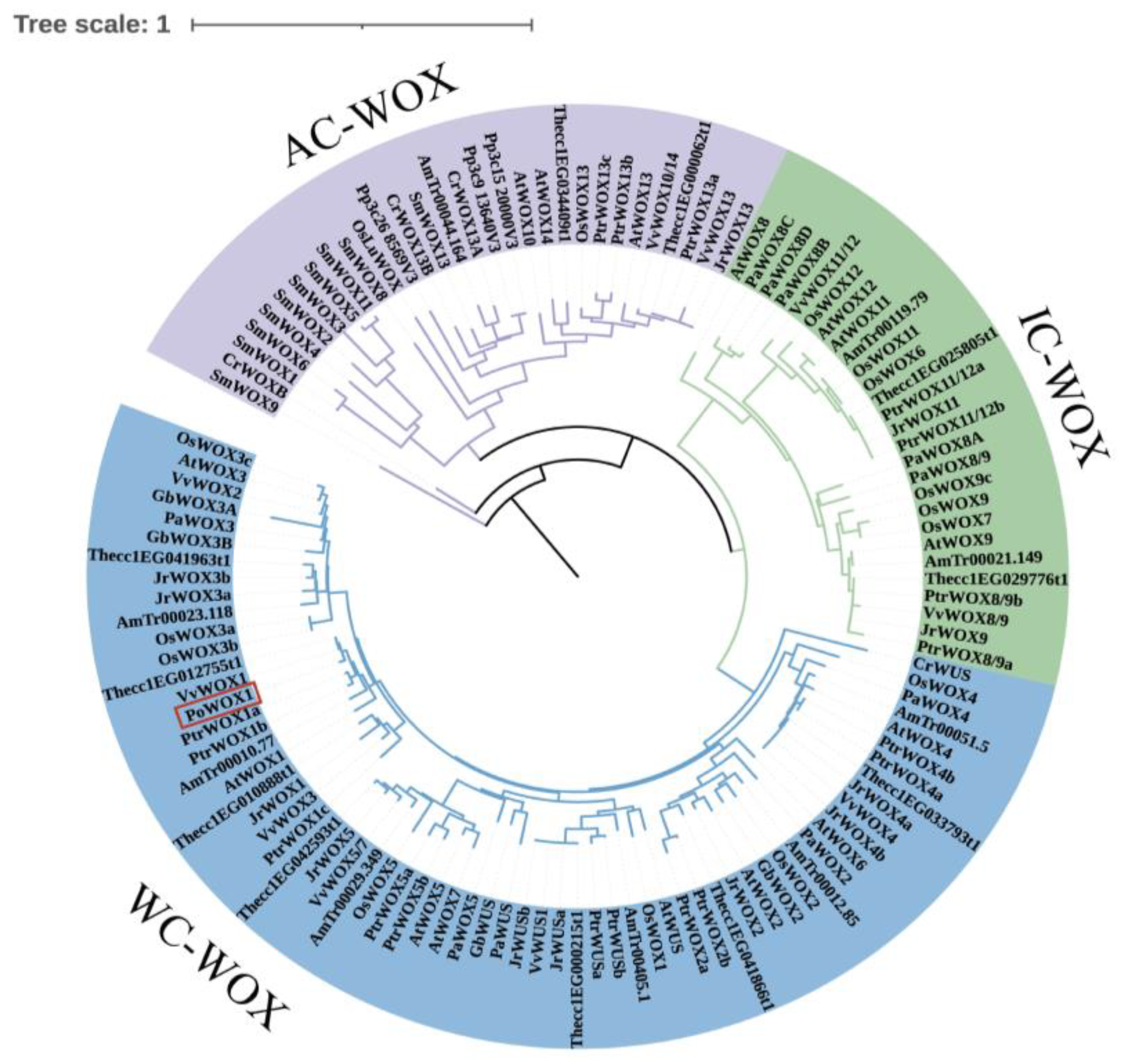
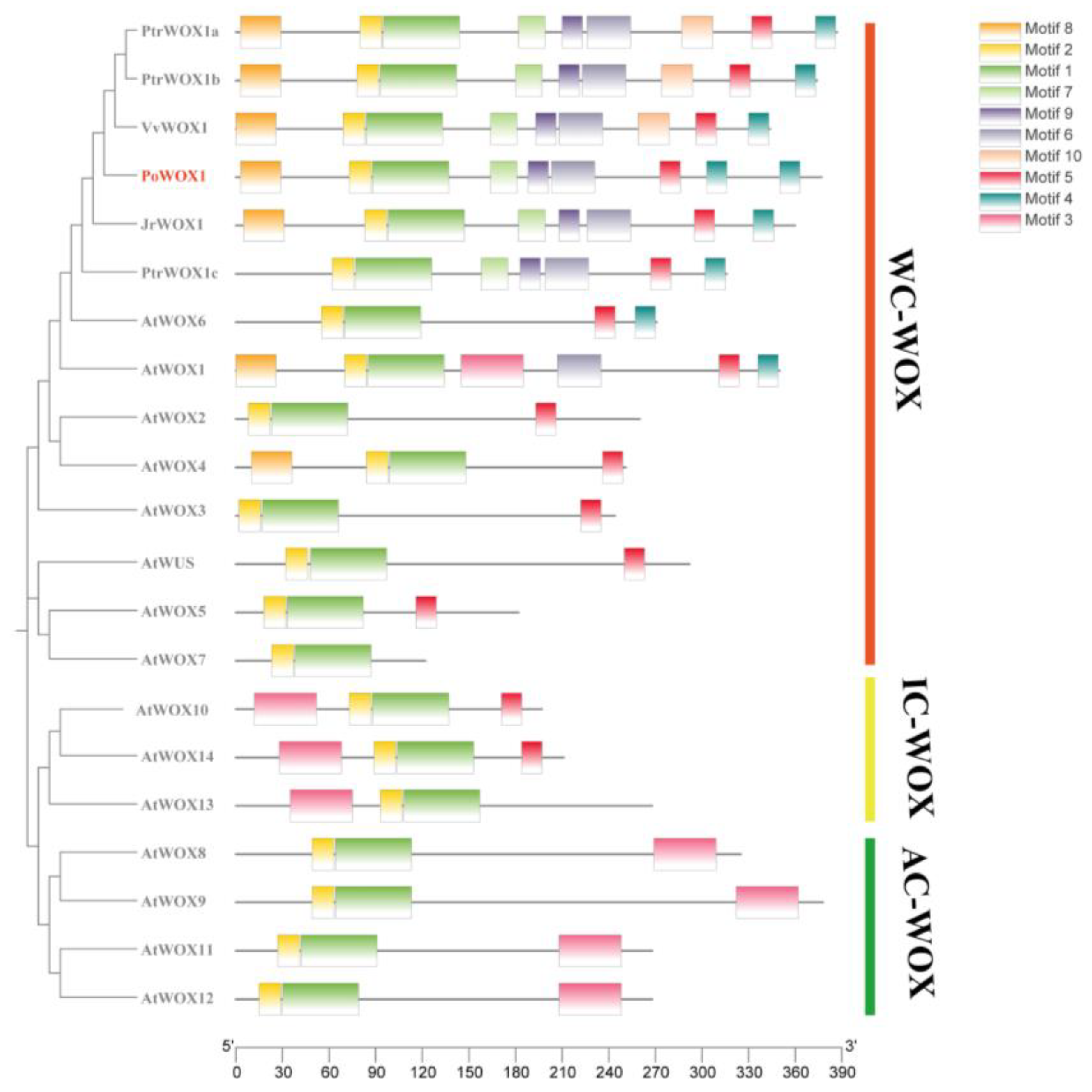
2.1. Bioinformatic Analysis of PoWOX1
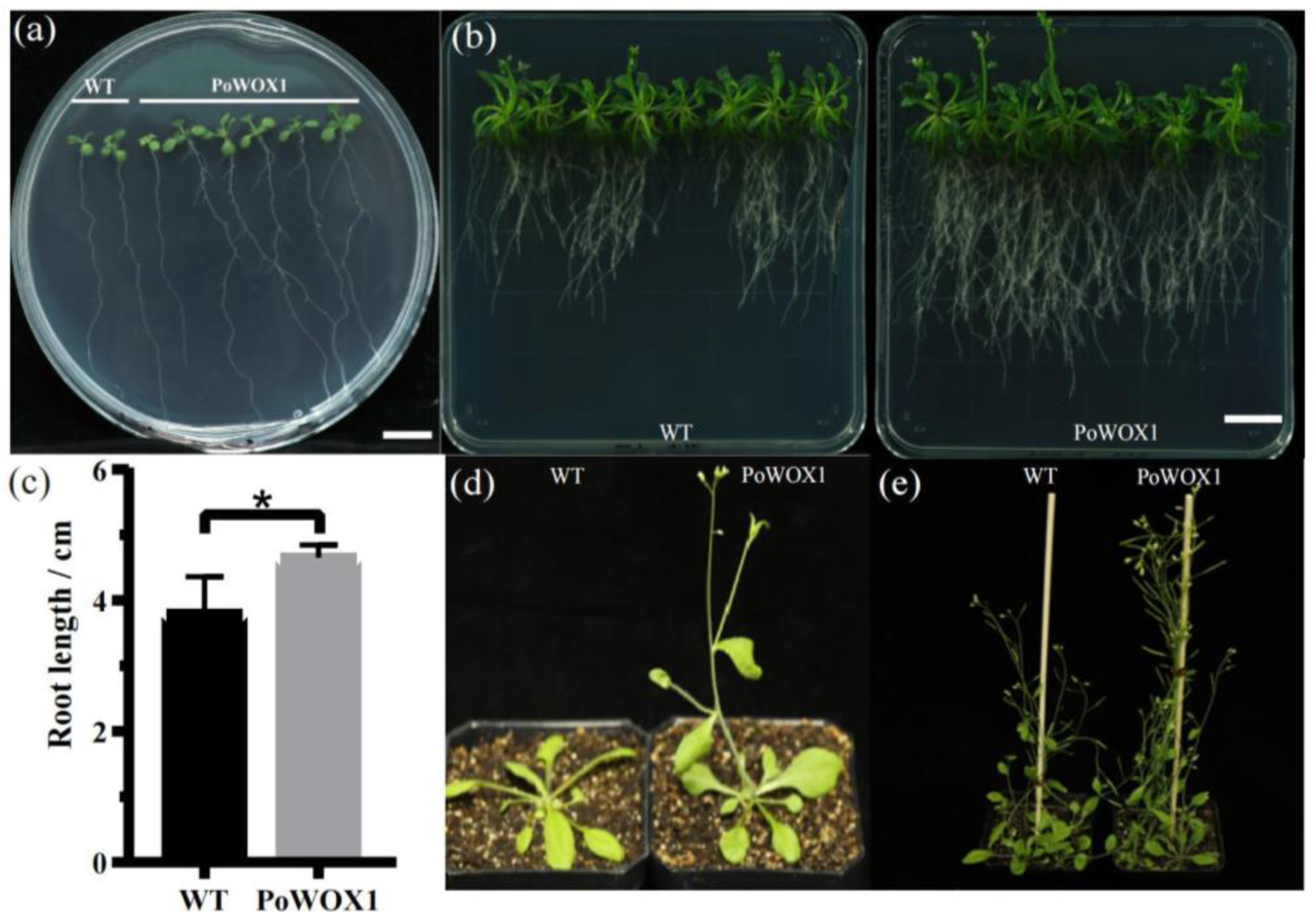
2.2. Phenotypic Analysis of Arabidopsis Plants Overexpressing PoWOX1
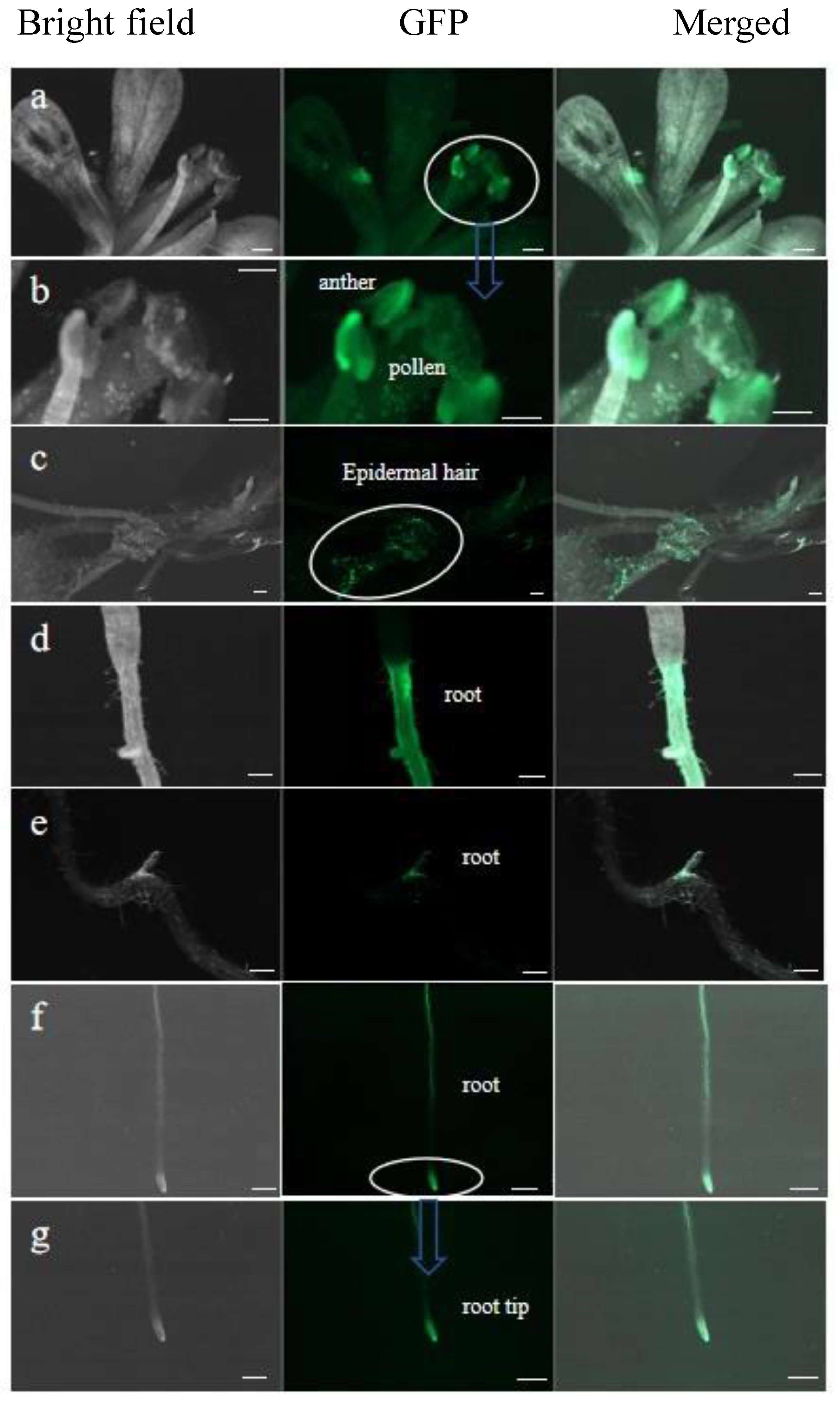
2.3. Assessment of PoWOX1 Expression in Arabidopsis Transformants
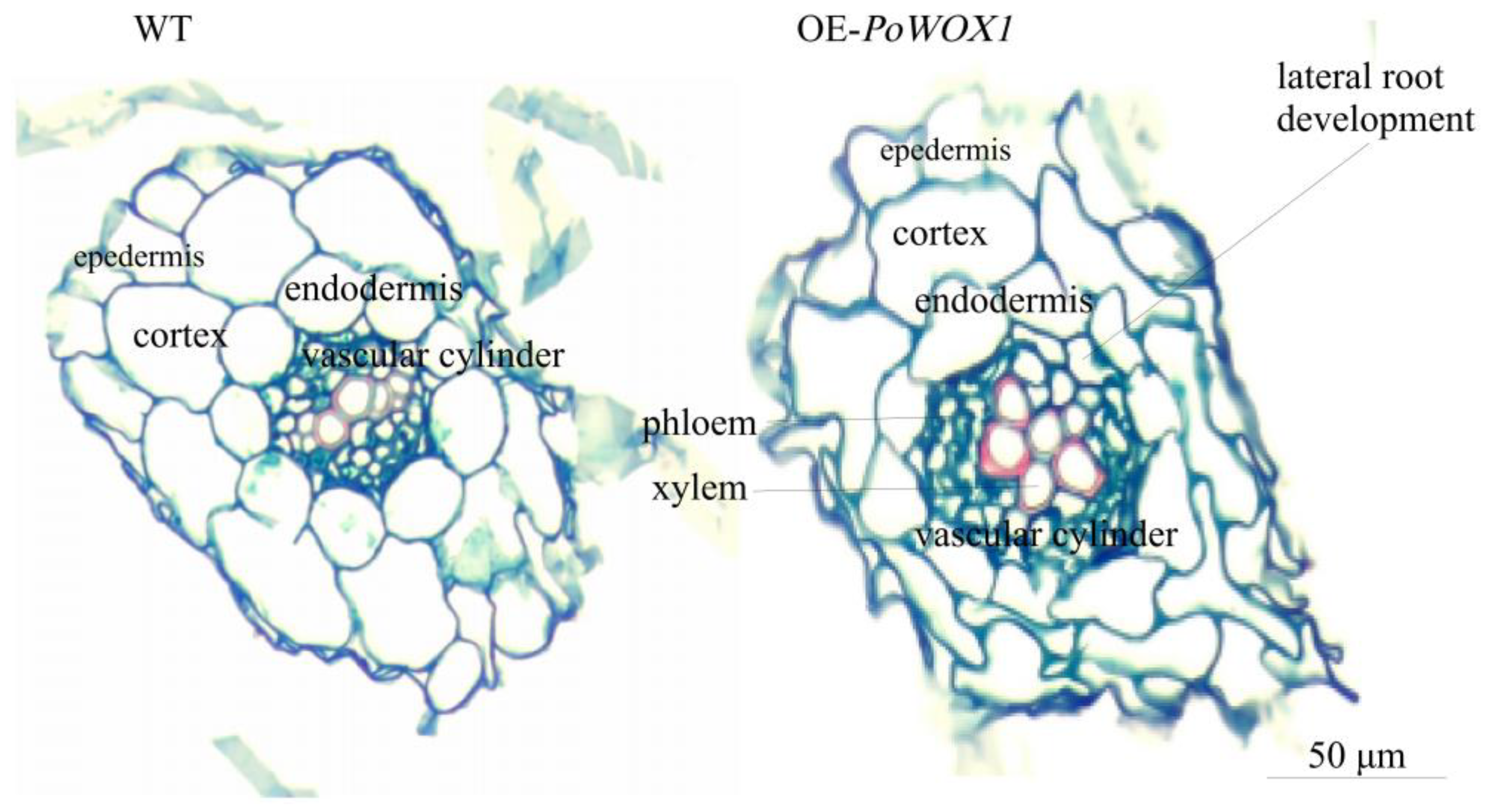
2.4. Analysis of Arabidopsis Roots Overexpressing PoWOX1
2.5. Callus Tissue Induction and Differentiation from the Leaves and Roots of PoWOX1-Overexpressing Arabidopsis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Cultivation Conditions
4.2. Construction of the Plant Expression Vector and Transformation of Agrobacterium Cells
4.3. Generation of Arabidopsis Thaliana Overexpressing PoWOX1
4.4. Phenotypic Analysis of Roots and Leaves Differentiated from Arabidopsis Healing Tissue
4.5. Histological Analysis of Arabidopsis Roots
4.6. RNA Extraction and Real-Time PCR
4.7. Sequence Alignment and Phylogenetic Tree Generation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gao, J.; Xue, J.Q.; Xue, Y.Q.; Liu, R.; Ren, X.X.; Wang, S.L.; Zhang, X.X. Transcriptome sequencing and identification of key callus browning-related genes from petiole callus of tree peony (Paeonia suffruticosa cv. Kao) cultured on media with three browning inhibitors. Plant Physiol. Biochem. PPB 2020, 149, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Dong, C.L.; Xue, Z.Y.; Jin, Q.J.; Xu, Y.C. De novo transcriptome sequencing and discovery of genes related to copper tolerance in Paeonia ostii. Gene 2016, 576, 126–135. [Google Scholar] [CrossRef]
- Zhang, H.F.; Li, X.F.; Wu, K.; Wang, M.K.; Liu, P.; Wang, X.S.; Deng, R.X. Antioxidant Activities and Chemical Constituents of Flavonoids from the Flower of Paeonia ostii. Molecules 2016, 22, 5. [Google Scholar] [CrossRef]
- Yang, Y.; Sun, M.; Li, S.S.; Chen, Q.H.; Teixeira da Silva, J.A.; Wang, A.J.; Yu, X.N.; Wang, L.S. Germplasm resources and genetic breeding of Paeonia: A systematic review. Hortic. Res. 2020, 7, 107. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liu, Z.Y.; Lyu, M.L.; Yuan, Y.; Wu, B.H. Characterization of JsWOX1 and JsWOX4 during Callus and Root Induction in the Shrub Species Jasminum sambac. Plants 2019, 8, 79. [Google Scholar] [CrossRef]
- Du, L.; Kang, X.l.; Guo, H.r.; Zhu, Z.f.; Wu, R.; Yuan, M.j.; Ding, C.Y. Cotyledonary somatic embryo is one kind of intermediate material similar to callus in the process of in vitro tissue culture from Rosa hybrida ‘John F. Kennedy’. BMC Genom. 2024, 25, 362. [Google Scholar] [CrossRef]
- Zhang, K.L.; Yao, L.J.; Zhang, Y.; Baskin, J.M.; Baskin, C.C.; Xiong, Z.M.; Tao, J. A review of the seed biology of Paeonia species (Paeoniaceae), with particular reference to dormancy and germination. Planta 2019, 249, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Yang, Y.M.; Chen, J.C.; Lin, X.L.; Zhang, H.; Wang, H.; Wang, H.Z.; Bie, X.M.; Jiang, J.F.; Feng, X.Q.; et al. Dynamic chromatin regulatory programs during embryogenesis of hexaploid wheat. Genome Biol. 2023, 24, 7. [Google Scholar] [CrossRef]
- Hesami, M.; Pepe, M.; de Ronne, M.; Yoosefzadeh-Najafabadi, M.; Adamek, K.; Torkamaneh, D.; Jones, A.M.P. Transcriptomic Profiling of Embryogenic and Non-Embryogenic Callus Provides New Insight into the Nature of Recalcitrance in Cannabis. Int. J. Mol. Sci. 2023, 24, 14625. [Google Scholar] [CrossRef]
- Ramakrishnan, M.; Zhou, M.B.; Ceasar, S.A.; Ali, D.J.; Maharajan, T.; Vinod, K.K.; Sharma, A.; Ahmad, Z.; Wei, Q. Epigenetic modifications and miRNAs determine the transition of somatic cells into somatic embryos. Plant Cell Rep. 2023, 42, 1845–1873. [Google Scholar] [CrossRef]
- Joshi, S.; Hill, K.; Chakrabarti, M.; Perry, S.E. Regulatory mechanisms of the LBD40 transcription factor in Arabidopsis thaliana somatic embryogenesis. Plant Direct 2023, 7, e547. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.Y.; Kagale, S.; Ferrie, A.M.R. Multifaceted roles of transcription factors during plant embryogenesis. Front. Plant Sci. 2023, 14, 1322728. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Ryan, L.; Sardesai, N.; Wu, E.; Lenderts, B.; Lowe, K.; Che, P.; Anand, A.; Worden, A.; van Dyk, D.; et al. Leaf transformation for efficient random integration and targeted genome modification in maize and sorghum. Nat. Plants 2023, 9, 255–270. [Google Scholar] [CrossRef] [PubMed]
- Gaj, M.D.; Zhang, S.; Harada, J.J.; Lemaux, P.G. Leafy cotyledon genes are essential for induction of somatic embryogenesis of Arabidopsis. Planta 2005, 222, 977–988. [Google Scholar] [CrossRef]
- Schmidt, E.D.; Guzzo, F.; Toonen, M.A.; de Vries, S.C. A leucine-rich repeat containing receptor-like kinase marks somatic plant cells competent to form embryos. Development 1997, 124, 2049–2062. [Google Scholar] [CrossRef]
- Song, J.P.; Xie, X.; Chen, C.; Shu, J.; Thapa, R.K.; Nguyen, V.; Bian, S.M.; Kohalmi, S.E.; Marsolais, F.; Zou, J.T.; et al. LEAFY COTYLEDON1 expression in the endosperm enables embryo maturation in Arabidopsis. Nat. Commun. 2021, 12, 3963. [Google Scholar] [CrossRef]
- Srinivasan, C.; Liu, Z.; Heidmann, I.; Supena, E.D.; Fukuoka, H.; Joosen, R.; Lambalk, J.; Angenent, G.; Scorza, R.; Custers, J.B.; et al. Heterologous expression of the BABY BOOM AP2/ERF transcription factor enhances the regeneration capacity of tobacco (Nicotiana tabacum L.). Planta 2007, 225, 341–351. [Google Scholar] [CrossRef]
- Lee, H.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. Arabidopsis LEAFY COTYLEDON1 represents a functionally specialized subunit of the CCAAT binding transcription factor. Proc. Natl. Acad. Sci. USA 2003, 100, 2152–2156. [Google Scholar] [CrossRef]
- Jacobson, S.; Bondarchuk, N.; Nguyen, T.A.; Canada, A.; McCord, L.; Artlip, T.S.; Welser, P.; Klocko, A.L. Apple CRISPR-Cas9-A Recipe for Successful Targeting of AGAMOUS-like Genes in Domestic Apple. Plants 2023, 12, 3693. [Google Scholar] [CrossRef]
- Xu, L.; Liu, Y.; Zhang, J.J.; Wu, W.H.; Hao, Z.D.; He, S.C.; Li, Y.R.; Shi, J.S.; Chen, J.H. Genomic survey and expression analysis of LcARFs reveal multiple functions to somatic embryogenesis in Liriodendron. BMC Plant Biol. 2024, 24, 94. [Google Scholar] [CrossRef]
- Wang, F.X.; Shang, G.D.; Wu, L.Y.; Xu, Z.G.; Zhao, X.Y.; Wang, J.W. Chromatin Accessibility Dynamics and a Hierarchical Transcriptional Regulatory Network Structure for Plant Somatic Embryogenesis. Dev. Cell 2020, 54, 742–757.e748. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Niu, H.H.; Li, C.; Shen, G.Y.; Liu, X.F.; Weng, Y.Q.; Wu, T.; Li, Z. WUSCHEL-related homeobox1 (WOX1) regulates vein patterning and leaf size in Cucumis sativus. Hortic. Res. 2020, 7, 182. [Google Scholar] [CrossRef]
- Kamiya, N.; Nagasaki, H.; Morikami, A.; Sato, Y.; Matsuoka, M. Isolation and characterization of a rice WUSCHEL-type homeobox gene that is specifically expressed in the central cells of a quiescent center in the root apical meristem. Plant J. Cell Mol. Biol. 2003, 35, 429–441. [Google Scholar] [CrossRef]
- Mayer, K.F.; Schoof, H.; Haecker, A.; Lenhard, M.; Jürgens, G.; Laux, T. Role of WUSCHEL in regulating stem cell fate in the Arabidopsis shoot meristem. Cell 1998, 95, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Graaff, E.; Laux, T.; Rensing, S.A. The WUS homeobox-containing (WOX) protein family. Genome Biol. 2009, 10, 248. [Google Scholar]
- Wang, C.Q.; Zhao, B.L.; He, L.L.; Zhou, S.L.; Liu, Y.; Zhao, W.Y.; Guo, S.Q.; Wang, R.R.; Bai, Q.Z.; Li, Y.H.; et al. The WOX family transcriptional regulator SlLAM1 controls compound leaf and floral organ development in Solanum lycopersicum. J. Exp. Bot. 2021, 72, 1822–1835. [Google Scholar] [CrossRef]
- Haecker, A.; Gross-Hardt, R.; Geiges, B.; Sarkar, A.; Breuninger, H.; Herrmann, M.; Laux, T. Expression dynamics of WOX genes mark cell fate decisions during early embryonic patterning in Arabidopsis thaliana. Development 2004, 131, 657–668. [Google Scholar] [CrossRef]
- Dolzblasz, A.; Nardmann, J.; Clerici, E.; Causier, B.; van der Graaff, E.; Chen, J.; Davies, B.; Werr, W.; Laux, T. Stem Cell Regulation by Arabidopsis WOX Genes. Mol. Plant 2016, 9, 1028–1039. [Google Scholar] [CrossRef] [PubMed]
- Tadege, M.; Lin, H.; Bedair, M.; Berbel, A.; Wen, J.Q.; Rojas, C.M.; Niu, L.F.; Tang, Y.H.; Sumner, L.; Ratet, P.; et al. STENOFOLIA regulates blade outgrowth and leaf vascular patterning in Medicago truncatula and Nicotiana sylvestris. Plant Cell 2011, 23, 2125–2142. [Google Scholar] [CrossRef]
- Lou, X.Y.; Wang, J.G.; Wang, G.Q.; He, D.; Shang, W.Q.; Song, Y.L.; Wang, Z.; He, S.L. Genome-Wide Analysis of the WOX Family and Its Expression Pattern in Root Development of Paeonia ostii. Int. J. Mol. Sci. 2024, 25, 7668. [Google Scholar] [CrossRef]
- Jha, P.; Ochatt, S.J.; Kumar, V. WUSCHEL: A master regulator in plant growth signaling. Plant Cell Rep. 2020, 39, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Niu, L.F.; McHale, N.A.; Ohme-Takagi, M.; Mysore, K.S.; Tadege, M. Evolutionarily conserved repressive activity of WOX proteins mediates leaf blade outgrowth and floral organ development in plants. Proc. Natl. Acad. Sci. USA 2013, 110, 366–371. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Wu, R.H.; Qin, G.J.; Chen, Z.L.; Gu, H.Y.; Qu, L.J. Over-expression of WOX1 leads to defects in meristem development and polyamine homeostasis in Arabidopsis. J. Integr. Plant Biol. 2011, 53, 493–506. [Google Scholar] [CrossRef]
- Nakata, M.; Matsumoto, N.; Tsugeki, R.; Rikirsch, E.; Laux, T.; Okada, K. Roles of the middle domain-specific WUSCHEL-RELATED HOMEOBOX genes in early development of leaves in Arabidopsis. Plant Cell 2012, 24, 519–535. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Mo, Y.J.; Israeli, A.; Wang, Q.Q.; Yifhar, T.; Ori, N.; Jiao, Y.L. Leaflet initiation and blade expansion are separable in compound leaf development. Plant J. Cell Mol. Biol. 2020, 104, 1073–1087. [Google Scholar] [CrossRef]
- Gao, B.; Wen, C.; Fan, L.S.; Kou, Y.P.; Ma, N.; Zhao, L.J. A Rosa canina WUSCHEL-related homeobox gene, RcWOX1, is involved in auxin-induced rhizoid formation. Plant Mol. Biol. 2014, 86, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Z.; Yuan, X.; Zhao, Y.G.; Wang, X.H.; Lu, L.; Wang, H.J.; Li, Y.M.; Gao, J.; Wang, L.M.; Zhang, H.C. Identification of ARF Genes and Elucidation of the Regulatory Effects of PsARF16a on the Dormancy of Tree Peony Plantlets. Genes 2024, 15, 666. [Google Scholar] [CrossRef]
- Song, Y.L.; Wang, J.G.; Zhu, J.L.; Shang, W.Q.; Jia, W.; Sun, Y.K.; He, S.L.; Yang, X.T.; Wang, Z. Functional Analysis of the PoSERK-Interacting Protein PorbcL in the Embryogenic Callus Formation of Tree Peony (Paeonia ostii T. Hong et J. X. Zhang). Plants 2024, 13, 2697. [Google Scholar] [CrossRef]
- Ge, Y.C.; Liu, J.; Zeng, M.H.; He, J.F.; Qin, P.; Huang, H.; Xu, L. Identification of WOX Family Genes in Selaginella kraussiana for Studies on Stem Cells and Regeneration in Lycophytes. Front. Plant Sci. 2016, 7, 93. [Google Scholar] [CrossRef]
- Nakata, M.T.; Tameshige, T.; Takahara, M.; Mitsuda, N.; Okada, K. The functional balance between the WUSCHEL-RELATED HOMEOBOX1 gene and the phytohormone auxin is a key factor for cell proliferation in Arabidopsis seedlings. Plant Biotechnol. 2018, 35, 141–154. [Google Scholar] [CrossRef]
- Ikeda, M.; Mitsuda, N.; Ohme-Takagi, M. Arabidopsis WUSCHEL is a bifunctional transcription factor that acts as a repressor in stem cell regulation and as an activator in floral patterning. Plant Cell 2009, 21, 3493–3505. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.H.; Jiang, S.J.; Jian, J.B.; Liu, M.Y.; Yue, Z.; Xu, J.B.; Li, J.; Xu, C.Y.; Lin, L.H.; Jing, Y.; et al. Genomic basis of the giga-chromosomes and giga-genome of tree peony Paeonia ostii. Nat. Commun. 2022, 13, 7328. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, W.B.; Chang, Y.T.; Ma, Y.J.; Deng, Y.Y.; Zhang, N.; Bai, Y.W.; Jiang, Z.H.; Hu, T. Cloning, Characterization, and Expression Pattern Analysis of the BBM Gene in Tree Peony (Paeonia ostii). Forests 2023, 15, 36. [Google Scholar] [CrossRef]
- Sang, Y.L.; Lin, J.C.; Xian, S.Z. Plant stem cells and de novo organogenesis. New Phytol. 2018, 218, 1334–1339. [Google Scholar] [CrossRef]
- Sugimoto, K.; Gordon, S.P.; Meyerowitz, E.M. Regeneration in plants and animals: Dedifferentiation, transdifferentiation, or just differentiation? Trends Cell Biol. 2011, 21, 212–218. [Google Scholar] [CrossRef]
- Zhang, Y.; Mitsuda, N.; Yoshizumi, T.; Horii, Y.; Oshima, Y.; Ohme-Takagi, M.; Matsui, M.; Kakimoto, T. Two types of bHLH transcription factor determine the competence of the pericycle for lateral root initiation. Nat. Plants 2021, 7, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Deyhle, F.; Sarkar, A.K.; Tucker, E.J.; Laux, T. WUSCHEL regulates cell differentiation during anther development. Dev. Biol. 2007, 302, 154–159. [Google Scholar] [CrossRef]
- Xia, M.S.; Zhang, W.B.; Chang, Y.T.; Ma, Y.J.; Deng, Y.Y.; Fan, K.K.; Zhang, X.; Jiang, Z.H.; Hu, T. A Preliminary Investigation on the Functional Validation and Interactions of PoWOX Genes in Peony (Paeonia ostii). Horticulturae 2022, 8, 266. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Primer Sequence (5′-3′) | Purpose |
|---|---|---|
| HPT-F | GGTCGCGGAGGCTATGGATGC | PCR identification of positive seedlings |
| HPT-R | GCTTCTGCGGGCGATTTGTGT | |
| Q-PoWOX1-F | CGTTGGCGGCAATGAAGAAGAATC | Real-time PCR |
| Q-PoWOX1-R | GGCAATTAGGAGGACTCAAGTTGGTAT | |
| Q-AtActin-F | GGTATGGGTCAGAAAGATGCT | |
| Q-AtActin-R | CGTTGTAGAAAGTGTGATGCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Hu, T.; Chang, Y.; Xia, M.; Ma, Y.; Deng, Y.; Jiang, Z.; Zhang, W. Overexpression of Peony PoWOX1 Promotes Callus Induction and Root Development in Arabidopsis thaliana. Plants 2025, 14, 1857. https://doi.org/10.3390/plants14121857
Zhang X, Hu T, Chang Y, Xia M, Ma Y, Deng Y, Jiang Z, Zhang W. Overexpression of Peony PoWOX1 Promotes Callus Induction and Root Development in Arabidopsis thaliana. Plants. 2025; 14(12):1857. https://doi.org/10.3390/plants14121857
Chicago/Turabian StyleZhang, Xue, Tao Hu, Yanting Chang, Mengsi Xia, Yanjun Ma, Yayun Deng, Zehui Jiang, and Wenbo Zhang. 2025. "Overexpression of Peony PoWOX1 Promotes Callus Induction and Root Development in Arabidopsis thaliana" Plants 14, no. 12: 1857. https://doi.org/10.3390/plants14121857
APA StyleZhang, X., Hu, T., Chang, Y., Xia, M., Ma, Y., Deng, Y., Jiang, Z., & Zhang, W. (2025). Overexpression of Peony PoWOX1 Promotes Callus Induction and Root Development in Arabidopsis thaliana. Plants, 14(12), 1857. https://doi.org/10.3390/plants14121857