
Analysis of Meiotic Behavior and 2n Pollen Formation Frequency in Triploid Hybrids of Chinese Jujube

Abstract
1. Introduction
2. Results
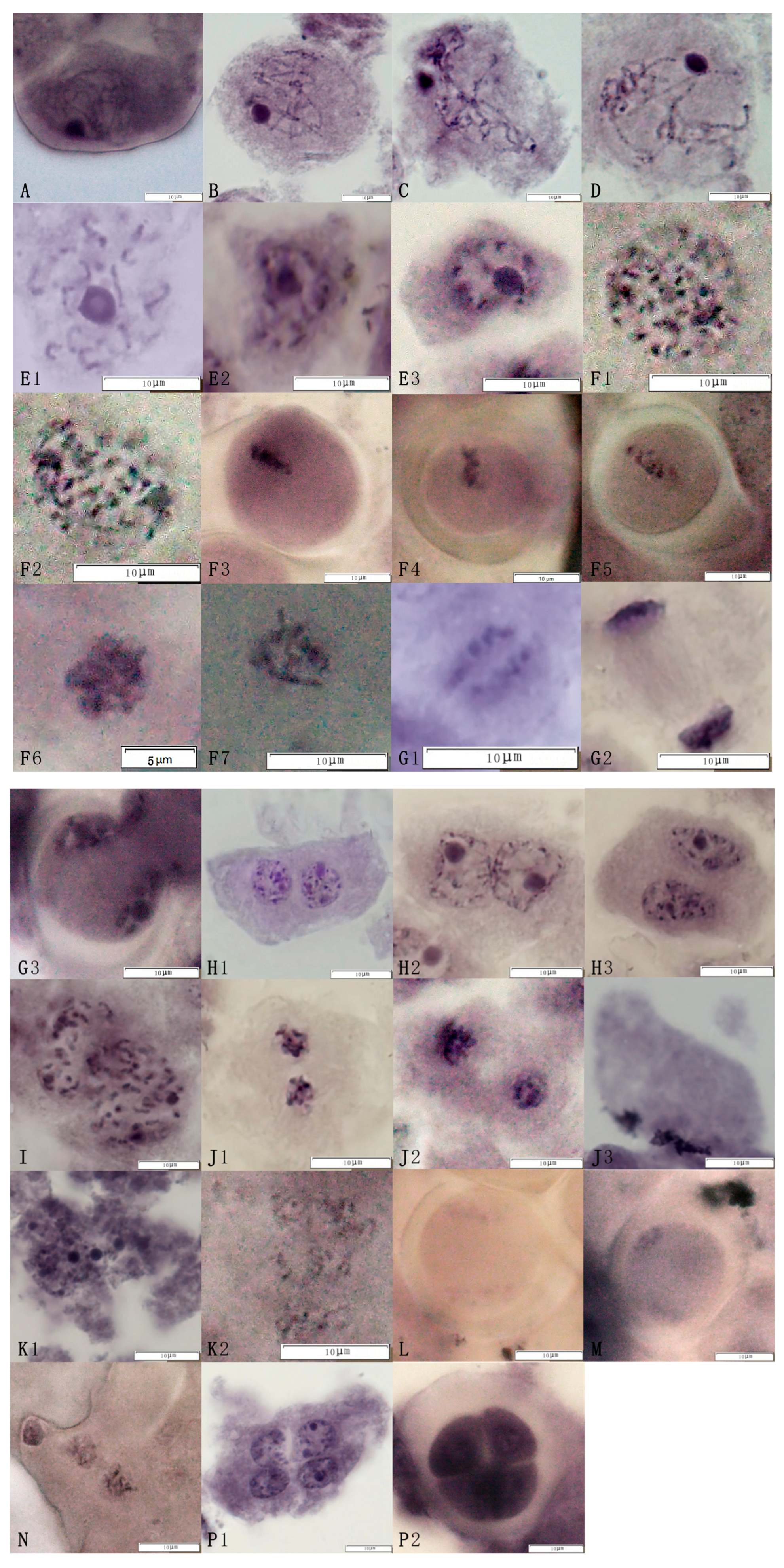
2.1. The Analysis of Meiosis and the Abnormal Behavior of Pollen Mother Cells of the Triploid Germplasm
2.1.1. Correspondence Between Flower Bud Size and Meiotic Period
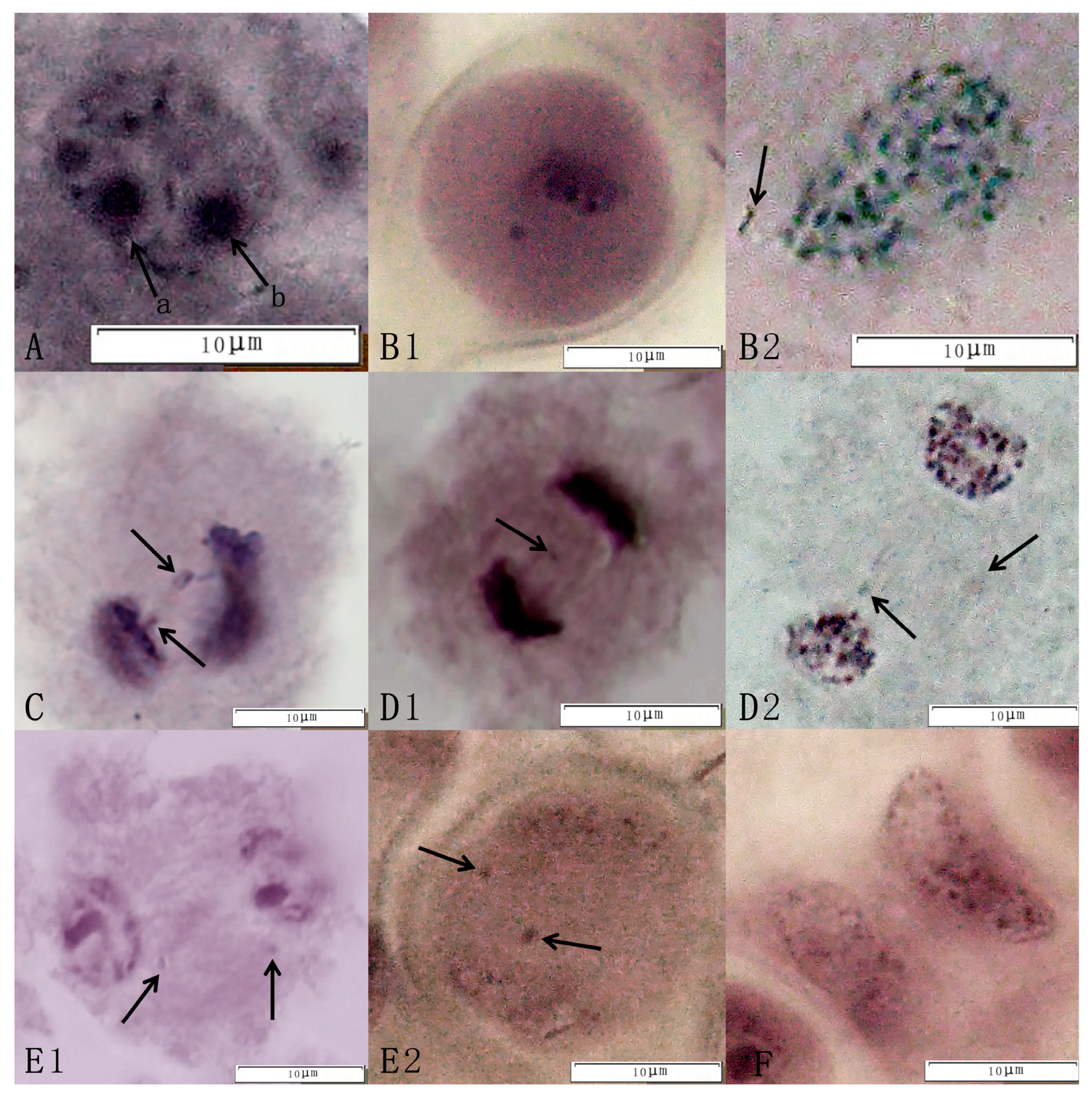
2.1.2. The Process of Meiosis
2.1.3. Statistics of Abnormal Meiotic Behavior
2.1.4. Determination of the Chromosome Number of Progeny Q161 of the Jujube Superior Line
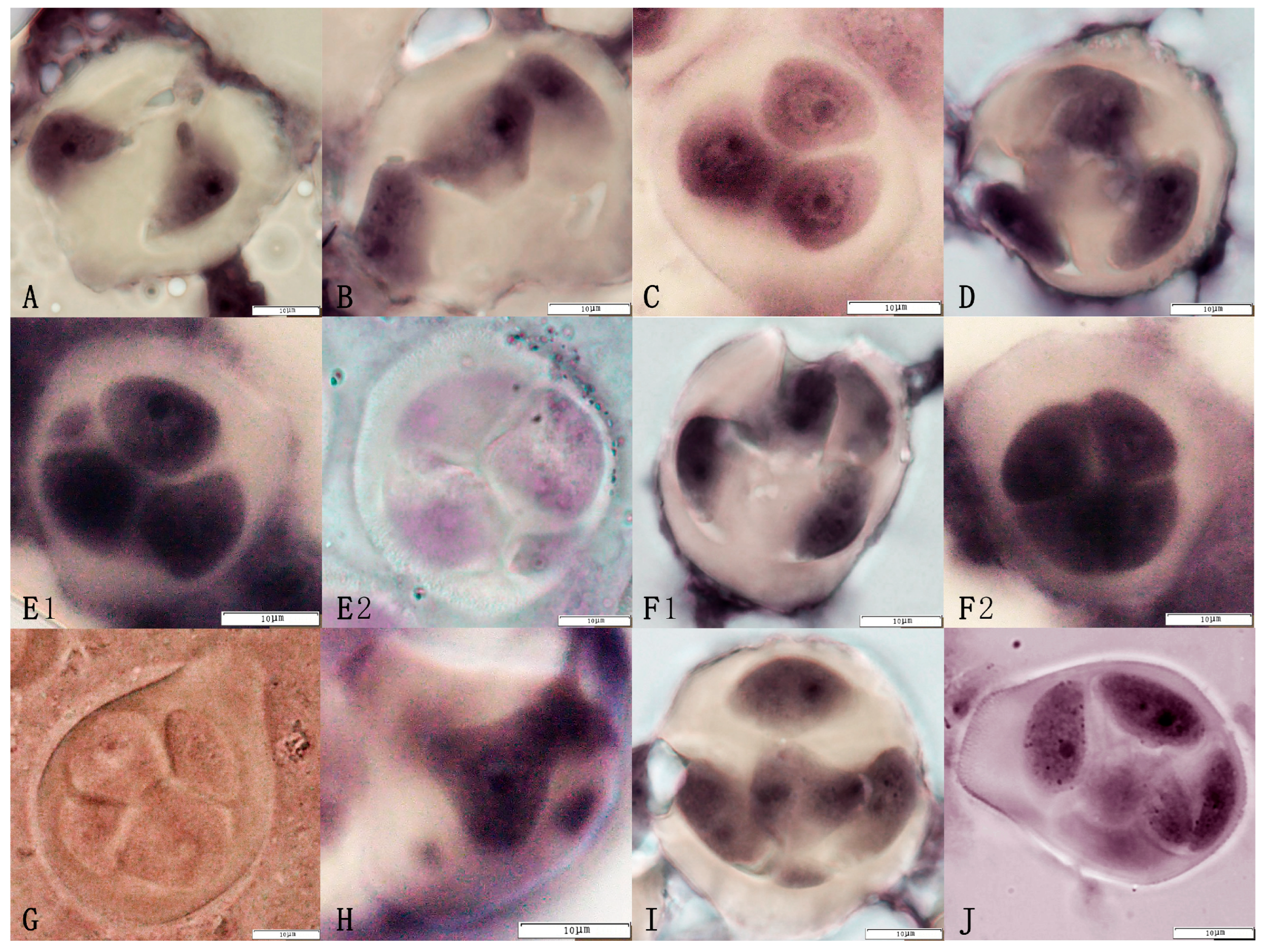
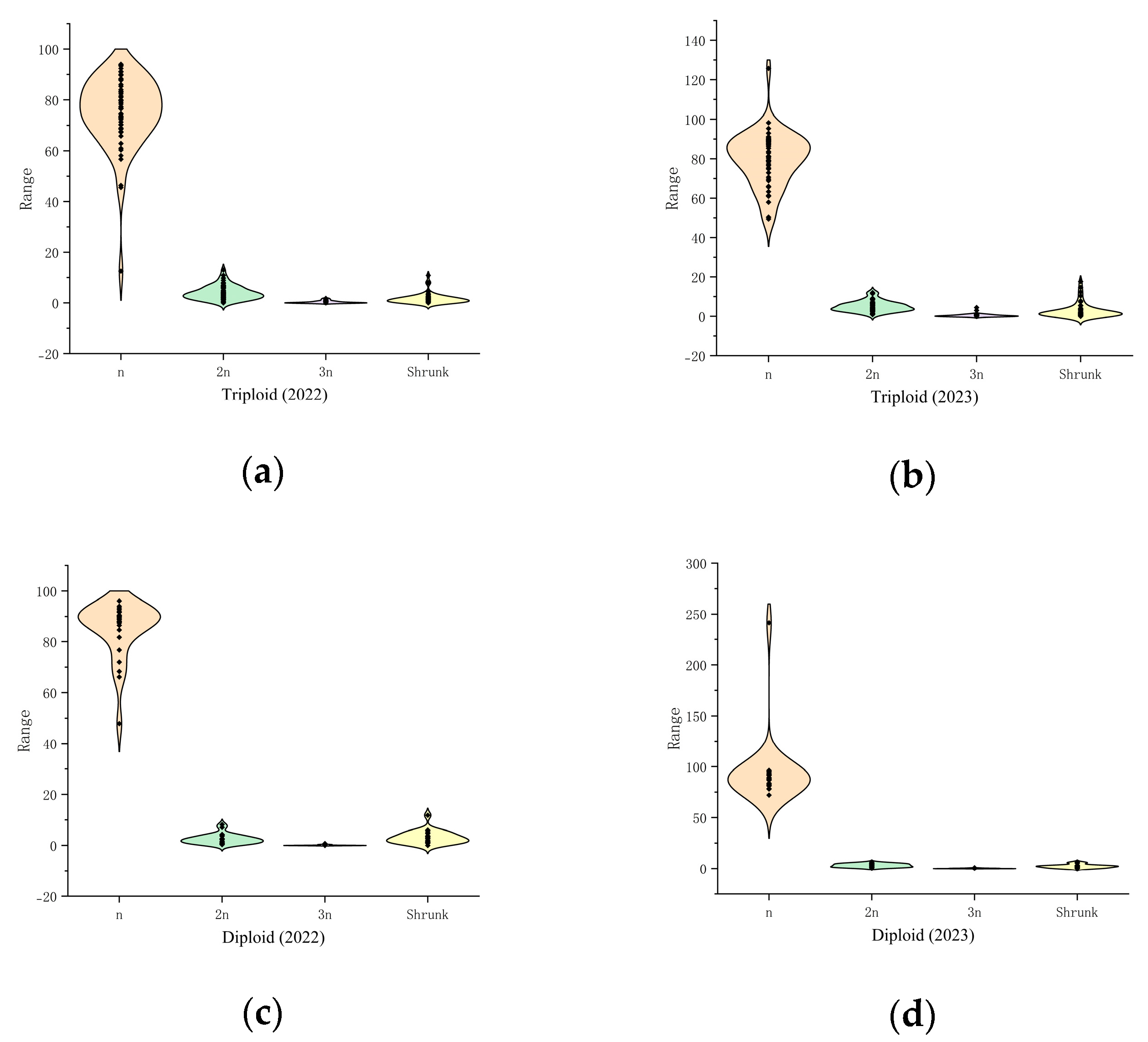
2.2. Natural 2n Pollen Distribution of the Triploid Germplasm
2.3. Analysis of Pollen Traits in Jujube Triploid Hybrid Progeny Clusters
2.4. Characterization of the Distribution of Natural 2n Pollen in Jujube Date Triploid Hybrid Progeny Clusters
2.5. The High 2n Pollen Line Germplasm Screening
3. Discussion
3.1. Analysis of Abnormal Meiotic Behavior in Triploids
3.2. Chromosome Number and Karyotype Analysis of Q161
3.3. Analysis of Natural 2n Pollen Occurrence in Triploid Hybrid Progeny
3.4. Comparative Analysis of Pollen Characteristics and Genetic Variation in Triploid Hybrid Progeny
4. Conclusions
5. Materials and Methods
5.1. Materials
5.2. Methods
5.2.1. Observations of Meiosis in Microspore Mother Cells
5.2.2. Determination of Pollen Traits
5.2.3. Determination of Pollen Type
5.2.4. Scanning Electron Microscopy of Pollen
5.3. Data Processing
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Xie, W.J.; Chen, X.L.; Shi, G.C.; Zhang, P. Flower Development and Nectar Secretion of Ziziphus jujuba ‘Dongzao’ Under Facility Cultivation. Acta Hortic. Sin. 2024, 51, 2735–2742. [Google Scholar]
- Liu, M.J.; Wang, J.R.; Liu, P.; Zhao, J.; Zhao, Z.H.; Dai, L.; Li, X.S.; Liu, Z.G. Historical Achievements and Frontier Advances in the Production ano Research of Chinese Jujube (Ziziphus jujuba) in China. Acta Hortic. Sin. 2015, 42, 1683–1698. [Google Scholar]
- Tang, H.X.; Pei, G.Y.; Zhang, Q.; Wang, Z.T. Location and Analysis of Quantitative Trait Loci in Chinese Jujube Fruit. Acta Hortic. Sin. 2023, 50, 754–764. [Google Scholar]
- Xin, Z.; Bing, B.Z.; Zhi, L.; Yuan, Y.; Zhao, Z.H.; Liu, M.J. Composition Analysis and Nutritional Value Evaluation of Amino Acids in the Fruit of 161 Jujube Cultivars. Plants 2023, 12, 1744. [Google Scholar] [CrossRef]
- Xie, S.P.; Xie, K.D.; Xia, Q.M.; Xiao, G.A.; Zhou, R.; Zhang, C.L.; Zheng, H.; Wu, X.M.; Guo, W.W. Efficient exploration and SSR identification of 53 doubled diploid seedlings from six local Citrus cultivars and germplasm resources. J. Fruit Sci. 2022, 39, 1–9. [Google Scholar]
- Tan, F.Q.; Zhang, M.; Xie, K.D.; Fan, Y.J.; Song, X.; Wang, R.; Wu, X.M.; Zhang, H.Y.; Guo, W.W. Polyploidy remodels fruit metabolism by modifying carbon source utilization and metabolic flux in Ponkan mandarin (Citrus reticulata Blanco). Plant Sci. 2019, 289, 110276. [Google Scholar] [CrossRef]
- Zhi, W.; Guang, M.H.; Zuo, Z.L.; Zhong, C.H.; Yao, X.H. Characterizing Tetraploid Populations of Actinidia chinensis for Kiwifruit Genetic Improvement. Plants 2022, 11, 1154. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xie, S.P.; Xiao, G.A.; Xiao, G.A.; Zhou, R.; Wu, X.M.; Wu, Q.; Deng, J.R.; Ao, Y.J.; Liu, G.P. Efficient exploration and SSR identification of autotetraploids from the seedlings of thirteen apomictic Citrus genotypes. J. Fruit Sci. 2023, 40, 2297–2306. [Google Scholar]
- Thomas, P.T. A Useful Abnormality of the Pollen in a Pear. Nature 1942, 149, 168–169. [Google Scholar] [CrossRef]
- Dewitte, A.; Eeckhaut, T.; Van Huylenbroeck, J.; Van Bockstaele, E. Meiotic aberrations during 2n pollen formation in Begonia. Heredity 2010, 104, 215–223. [Google Scholar] [CrossRef]
- Wang, J.; Li, D.; Shang, F.; Kang, X. High temperature-induced production of unreduced pollen and its cytological effects in Populus. Sci. Rep. 2017, 7, 5281. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huo, B.; Liu, W.; Li, D.; Liao, L. Abnormal meiosis in an intersectional allotriploid of Populus L. and segregation of ploidy levels in 2x × 3x progeny. PLoS ONE 2017, 12, e0181767. [Google Scholar] [CrossRef]
- Barke, B.H.; Karbstein, K.; Daubert, M.; Hörandl, E. The relation of meiotic behaviour to hybridity, polyploidy and apomixis in the Ranunculus auricomus complex (Ranunculaceae). BMC Plant Biol. 2020, 20, 523. [Google Scholar] [CrossRef]
- Ting, W.; Rui, D.J.; Shu, H.Y. Research Advances and Prospects om Pjalaenopsis Polyploid Breeding. Acta Hortic. Sin. 2022, 49, 448–462. [Google Scholar]
- Schindfessel, C.; Jin, C.; Geelen, D. Heat induced male meiotic restitution and sexual polyploidisation through natural variation of the Arabidopsis cyclin TAM/CYCA1. bioRxiv 2023. [Google Scholar]
- Yi, X.H.; Peng, C.Y.; Qian, Z.; Yue, Z.Y.; Piao, M.L.; Gao, X.; Jia, G.X. Effective Induction of Fertile Gametes in Oriental × Trumpet (OT) Lily by High Temperatures. Plants 2023, 12, 1563. [Google Scholar] [CrossRef]
- Deng, H.; Zhang, X.; Sheng, S.; Huang, Y.X.; Ye, Z.Q.; Zhang, T.T.; Liu, X.; Han, Z.Q. High temperature treatment induced production of unreduced 2n pollen in Camellia oleifera. Hortic. Plant J. 2024, 10, 879–896. [Google Scholar] [CrossRef]
- Hui, M.L.; Ji, C.G.; Hai, X.; Mao, X.; Li, S.; Zhang, H.; Hu, G.; Liu, G.; Fu, J. Production of Triploid Germplasm by Inducing 2n Pollen in Longan. Horticulturae 2022, 8, 437. [Google Scholar] [CrossRef]
- Shao, F.; Wang, S.; Liu, Z.; Hong, R.Y.; Zhao, T.J. Pollination, Fertilization, and Embryo Development in Southern China Fresh-eating Jujube. HortScience 2020, 55, 1315–1322. [Google Scholar] [CrossRef]
- Zhi, Y.; Chuan, J.Z.; Yang, F.Y.; Dong, M.Y.; Wang, Z.L.; Yan, F.F.; Wu, C.Y.; Wang, J.R.; Liu, M.J.; Lin, M.J. Genetic analysis of mined models of fruit sugar-acid fractions in a cross between jujube (Ziziphus jujuba Mill.) and wild jujube (Z. acido jujuba). Front. Plant Sci. 2023, 14, 1181903. [Google Scholar] [CrossRef]
- Xu, X.G.; Chen, L.; Peng, J.Y. Meiotic Observations on the Pollen Mother Cells of Ziziphus spinosus Hu. and Ziziphus jujuba Mill. ‘Zanhuang Dazao’. Acta Hortic. Sin. 2009, 36, 1127–1133. [Google Scholar]
- Lu, Y.; Xue, Z.H.; Wu, G.E.; Liu, P.; Liu, M.J. Abnormal Meiosis Behaviors of Triploid and Tetraploid Chinese Jujube. Acta Hortic. Sin. 2018, 45, 659–668. [Google Scholar]
- Lu, Y.; Hu, L.; Wang, L.H.; Liu, P.; Liu, M.J. Meiosis and 2n Pollen Formation of Ziziphus jujuba ‘Wuhefeng’ After Treated with Colchicine. Acta Hortic. Sin. 2017, 44, 11–22. [Google Scholar]
- Rouiss, H.; Cuenca, J.; Navarro, L.; Ollitrault, P.; Aleza, P. Tetraploid citrus progenies arising from FDR and SDR unreduced pollen in 4x X 2x hybridizations. Tree Genet. Genomes 2017, 13, 1–14. [Google Scholar] [CrossRef]
- Liu, X.S.; Chen, L.; Wang, J.X.; Li, L.; Peng, J.Y. Discovery and Identification of Natural Triploid Ploidy of Chinese Jujube Cultivar‘Pingguozao’. Acta Hortic. Sin. 2013, 40, 426–432. [Google Scholar]
- Kang, X.Y. Study on Mechanism of Pollen Abortion in Chinese White Poplar (Populus tomentosa Carr.). Sci. Silvae Sin. 2001, 37, 35–39+145. [Google Scholar]
- Park, H.R.; Park, J.E.; Kim, J.H.; Shin, H.; Yu, S.H.; Son, S.; Yi, G.; Lee, S.S.; Kim, H.H.; Huh, J.H. Meiotic Chromosome Stability and Suppression of Crossover Between Non-homologous Chromosomes in x Brassicoraphanus, an Intergeneric Allotetraploid Derived From a Cross Between Brassica rapa and Raphanus sativus. Front Plant Sci. 2020, 11, 851. [Google Scholar] [CrossRef]
- Cheng, Z.; Song, W.; Zhang, X. Genic male and female sterility in vegetable crops. Hortic Res. 2022, 10, uhac232. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yang, F.; Fernández-Jiménez, N.; Tučková, M.; Vrána, J.; Cápal, P.; Díaz, M.; Pradillo, M.; Pecinka, A. Defects in meiotic chromosome segregation lead to unreduced male gametes in Arabidopsis SMC5/6 complex mutants. Plant Cell. 2021, 33, 3104–3119. [Google Scholar] [CrossRef]
- Li, Z.F.; Wang, J.; Hu, Y.H.; Liu, Q.H.; Qin, J. Karyotype Analysis and Cytology Observation on Meiosis of Paeonia ostii ‘Phoenix White’. Acta Hortic. Sin. 2007, 411–416. [Google Scholar]
- Zhao, Y.L.; Maize, Z. Perennis allotetraploid as a bridge to transferexogenous genetic material to maize. Master’s Thesis, Sichuan Agricultural University, Ya’an, China, 2019. [Google Scholar]
- Li, D.L.; Shang, G.J.; Tian, J.; Song, L.J.; Liu, C.H.; Li, Y.C.; Kang, X.Y.; Wang, J. Meiotic chromosome behavior of pollen mother cells and pollen variation intriploid hybrid between section Tacamahaca and sect. Aigeiros of Populus. J. Beijing For. Univ. 2019, 41, 75–82. [Google Scholar]
- Li, X.; Niu, X.X.; Bao, Y.C. Observation on meiosis of pollen mother cells of super-male plants in Asparagus officinalis L. China Cucurbits Veg. 2024, 37, 95–99. [Google Scholar]
- Dündar, G.; Teranishi, M.; Hidema, J. Autophagy-deficient Arabidopsis mutant atg5, which shows ultraviolet-B sensitivity, cannot remove ultraviolet-B-induced fragmented mitochondria. Photochem. Photobiol. Sci. 2020, 19, 1717–1729. [Google Scholar] [CrossRef]
- Shao, Y.Z.; Jing, L.; Lu, M.; Wang, H.L.; Zhou, H.; Ni, E.D.; Jiang, D.G.; Liu, Z.L.; Zhuang, C.X. OsAGO2 controls ROS production and the initiation of tapetal PCD by epigenetically regulating OsHXK1 expression in rice anthers. Proc. Natl. Acad. Sci. USA 2019, 116, 7549–7558. [Google Scholar]
- Leroux, C.; Bouton, S.; Kiefer-Meyer, M.C.; Fabrice, T.N.; Mareck, A.; Guénin, S.; Fournet, F.; Ringli, C.; Pelloux, J.; Driouich, A.; et al. PECTIN METHYLESTERASE48 is involved in Arabidopsis pollen grain germination. Plant Physiol. 2015, 167, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Chen, L. Cytological Studies on Ziziphus jujuba Mill. and Ziziphus spinosus. Master’s Thesis, Hebei Agricultural University, Baoding, China, 2010. [Google Scholar]
- Liu, M.J.; Liu, P.; Jiang, H.G.; Dai, L.; Wu, G.E.; Liu, Z.G. A New Tetraploidy Table Chinese Jujube Cultivar ‘Chenguang’. Acta Hortic. Sin. 2010, 37, 1539–1540. [Google Scholar]
- Shi, Q.H. In vivo Polyploid Induction via Callus in Chinese Jujube and Sour Jujube. PhD’s Thesis, Hebei Agricultural University, Baoding, China, 2010. [Google Scholar]
- Stebbins, G.L. Chromosomal Evolution in Higher Plants; Edward Amold: London, UK, 1971. [Google Scholar]
- Qu, Z.; Wang, Y.; Wen, Z. Karyotype Analysis of Triploid Zanhuang Jujube (Ziziphus jujuba Mill.). Hebei Fruits 1990, 23–25. [Google Scholar] [CrossRef]
- Chen, Y.L. Karyotype Analysis of Wild Jujube (Ziziphus jujuba var. spinosa) and Jinsixiaozao Jujube (Ziziphus jujuba ‘Jinsixiaozao’). China Fruits 1988, 37–38+63. [Google Scholar] [CrossRef]
- Yang, M.; Han, L.; Zhang, S.; Dai, L.; Li, B.; Han, S.; Zhao, J.; Liu, P.; Zhao, Z.; Liu, M. Insights into the evolution and spatial chromosome architecture of jujube from an updated gapless genome assembly. Plant Commun. 2023, 4, 100662. [Google Scholar] [CrossRef]
- Yin, Q.; Pan, Z.; Li, Y.; Xiong, H.; Joseph, M.; Yuan, D.Y.; Zou, F. Pollen and Floral Organ Morphology of 18 Oil-Tea Genotypes and Its Systematic Significance. Horticulturae 2024, 10, 524. [Google Scholar] [CrossRef]
- Lysak, M.A.; Berr, A.; Pecinka, A.; Schmidt, R.; McBreen, K.; Schubert, I. Mechanisms of chromosome number reduction in Arabidopsis thaliana and related Brassicaceae species. Proc. Natl. Acad. Sci. USA 2006, 103, 5224–5229. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cao, L.; Guo, Y.; Li, Z.; Niu, H.H. Enhancer of Shoot Regeneration 2 (ESR2) regulates pollen maturation and vitality in watermelon (Citrullus lanatus). J. Integr. Agric. 2024, 23, 3506–3521. [Google Scholar] [CrossRef]
- Liu, W.G. Gentic Variation and Adverse Stress Resistance Mechanism of Different Ploidy Watermelon (Citrullus lanatus). Ph.D. Thesis, Northwest A&F University, Xianyang, China, 2003. [Google Scholar]
- Zeng, D.; Guan, J.; Luo, J.; Zhao, L.B.; Li, Y.Z.; Chen, W.S.; Zhang, L.Q.; Ning, S.Z.; Yuan, Z.W.; Li, A.L.; et al. A transcriptomic view of the ability of nascent hexaploid wheat to tolerate aneuploidy. BMC Plant Biol. 2020, 20, 97. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.G.; Wang, M.Y.; Zhi, H. Observation and Comparison on Pollen Morphology of Different Ploidy Watermelon. Acta Hortic. Sin. 2003, 30, 328–330. [Google Scholar]
- Hu, G.; Gao, C.; Fan, X.; Gong, W.F.; Yuan, D.Y. Pollination Compatibility and Xenia in Camellia oleifera. HortScience 2020, 55, 898–905. [Google Scholar] [CrossRef]
- Ahmed, H.A.; Akdogan, G.; Özcan, S.F.; Barpete, S. Effect of Parental Genotypes and Their Reciprocal Crosses on Haploid Plant Production by Anther Culture and Confirmation of Double Haploids by Flow Cytometry in Bread Wheat. J. Agric. Sci. 2022, 28, 363–371. [Google Scholar] [CrossRef]
- Scheschke, K. The Need for Agricultural Innovation to Sustainably Feed the World by 2050. Resource 2016, 23, 35. [Google Scholar]
- Ghaleb, M.A.A.; Li, C.; Shahid, M.Q.; Yu, H.; Liang, J.; Chen, R.; Wu, J.; Liu, X. Heterosis analysis and underlying molecular regulatory mechanism in a wide-compatible neo-tetraploid rice line with long panicles. BMC Plant Biol. 2020, 20, 83. [Google Scholar] [CrossRef]
- Wang, D.; Ran, Y. Improving main agronomic characters of dihaploids with 2n gametes by means of sexual hybridisation and anther culture of potato (Solarium tuberosum). N. Z. J. Crop Hortic. Sci. 2000, 28, 1–8. [Google Scholar] [CrossRef]
- Yu, H.; Lin, T.; Meng, X.; Du, H.; Zhang, J.; Liu, G.; Chen, M.; Jing, Y.; Kou, L.; Li, X.; et al. A route to de novo domestication of wild allotetraploid rice. Cell 2021, 184, 1156–1170.E14. [Google Scholar] [CrossRef]
- Kang, X.; Wei, H. Breeding polyploid Populus: Progress and perspective. For. Res. 2022, 2, 4. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Xiang, S.Q.; Liang, G.L. Advances in Polyploid Breeding of Citrus. South China Fruits 2003, 16–19. [Google Scholar]
- Li, Z.; Zhao, Y.; Cheng, X.; Kong, B.; Sang, Y.R.; Zhou, Q.; Wu, J.; Zhang, P.D. High Temperature-Induced Spindle Destabilization Results in Aborted Pollen Production in Populus. Forests 2023, 14, 797. [Google Scholar] [CrossRef]
- Zhang, P.D. Research progress on the mechanism of 2n gamete occurrence in plants. J. Beijing For. Univ. 2023, 45, 1–8. [Google Scholar]
- Mullualem, D.; Tsega, A.; Mengie, T.; Fentie, D.; Kassa, Z.; Fassil, A.; Wondaferew, D.; Gelaw, T.A.; Astatkie, T. Genotype-by-environment interaction and stability analysis of grain yield of bread wheat (Triticum aestivum L.) genotypes using AMMI and GGE biplot analyses. Heliyon 2024, 10, e32918. [Google Scholar] [CrossRef]
- Yi, L.P.; Bao, J.K.; Wu, C.Y.; Wang, J.R.; Liu, M.J.; Yan, F.F. Genetic Variation of Flower Traits in F1 of Male Sterile Jujube ‘JMS2’ × ‘Jiaocheng 5’. Acta Agric. Boreali-Occident. Sin. 2023, 32, 1913–1921. [Google Scholar]
- Liu, J.X.; Liu, P.; Minavar, Y.; Wang, J.R.; Liu, M.J.; Yan, F.F. Comparison of Phenotypic Characters between Diploid and Autotetraploid of Chinese Jujube in Southern Xinjiang. Acta Agric. Boreali-Occident. Sin. 2022, 31, 595–602. [Google Scholar]
- Yi, H.L. Study on Heredity and Variation of Citrus Sexual Progenies Using Allotetraploid Somatic Hybrid as the Pollen Parent. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2005. [Google Scholar]
- Zhang, R.; Zhou, L.; Li, Y.; Ma, H.H.; Li, Y.W.; Ma, Y.Z.; Lv, R.J.; Yang, J.; Wang, W.R.; Alifu, A.; et al. Rapid Identification of Pollen- and Anther-Specific Genes in Response to High-Temperature Stress Based on Transcriptome Profiling Analysis in Cotton. Int. J. Mol. Sci. 2022, 23, 3378. [Google Scholar] [CrossRef]
- Tan, Z.; Yang, J.; Li, Q.; Su, F.; Yang, T.; Wang, W.; Aierxi, A.; Zhang, X.; Yang, W.; Kong, J.; et al. PollenDetect: An Open-Source Pollen Viability Status Recognition System Based on Deep Learning Neural Networks. Int. J. Mol. Sci. 2022, 23, 13469. [Google Scholar] [CrossRef]
- Yan, F.F.; Wang, J.R.; Feng, Y.F.; Lin, M.J.; Wu, C.Y.; Liu, M.J. Establishment and application of large-scale hybrid creation system in Chinese jujube. J. Fruit Sci. 2020, 37, 929–938. [Google Scholar]
- Du, W.W.; Duan, Q.; Yang, N.; Cai, J.J.; Lin, P.S.; Ma, L.L.; Wang, X.L.; Jia, W.J.; Cui, G.F. Screening of Test Methods and Determination of Pollen Viability of 30 Varieties of Foliage Begonia. S. China J. Agric. Sci. 2021, 34, 1521–1527. [Google Scholar]
- Peng, B. Study on 2n in Chinese Jujube. Master’s Thesis, Hebei Agricultural University, Baoding, China, 2008. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microspore Blastogenesis | Range of Flower Bud Diameters/mm | Morphological Characteristics of Flower Buds |
|---|---|---|
| Pollen mother cell period | 1.3–2.1 | Dark green; pollen outline gradually clearing; sepal slits gradually evident |
| Preliminary I | 2.1–2.8 | Dark green to yellowish green; slightly enlarged; sepal slits gradually evident |
| Terminal II | 2.8–3.1 | Yellowish-green; volume expansion evident; sepal slits largely evident |
| Mononuclear pollen | 3.1–3.9 | Yellowish-green; reaching maximum size, at large bud stage; sepals about to unfurl |
| Type | Quantity/pc | Proportion/% |
|---|---|---|
| Dyad | 14 | 4.60% |
| Trisomy | 23 | 7.50% |
| Trisomy + micronucleus | 6 | 1.90% |
| Tetrads | 237 | 77.20% |
| Tetrads + micronuclei | 21 | 6.80% |
| Pentads | 4 | 1.30% |
| Pentads + micronuclei | 1 | 0.30% |
| Hexads | 1 | 0.30% |
| Serial No. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Length (μm) | 2.430 | 2.94 | 1.765 | 1.625 | 1.408 | 1.188 | 1.801 | 1.487 | 0.828 | 1.411 | 1.241 | 1.908 |
| Serial No. | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 |
| Length (μm) | 2.789 | 1.885 | 1.252 | 2.266 | 2.056 | 1.160 | 1.503 | 2.331 | 1.686 | 1.916 | 1.173 | 1.687 |
| Serial No. | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 |
| Length (μm) | 3.059 | 1.877 | 2.735 | 0.941 | 0.998 | 1.603 | 1.955 | 1.486 | 1.213 | 1.206 | 1.211 | 2.74 |
| Pollen Traits | 2022 | 2023 |
|---|---|---|
| Pollen activity (%) | 39.81 ± 3.30 | 22.10 ± 6.39 |
| Pollen amount (%) | 461.11 ± 101.84 | 1522.22 ± 139.77 |
| Pollen diameter (μm) | 23.61 ± 4.05 | 22.40 ± 3.52 |
| Shrunk pollen (%) | 0.33 | 1.00 |
| n pollen rate (%) | 71.33 | 83.67 |
| 2n pollen rate (%) | 4.00 | 4.67 |
| 3n pollen rate (%) | 0.67 | 0.33 |
| Traits | Parents and Ploidy | Pollen Activity/% | Pollen Quantity/Grain | ||
|---|---|---|---|---|---|
| 2022 | 2023 | 2022 | 2023 | ||
| Mean ± SD | DZ (2X) | 16.56 ± 8.90 b | 16.89 ± 3.17 a | 2427.78 ± 233.53 a | 1155.56 ± 108.44 ab |
| CG (4X) | 38.60 ± 13.17 b | 33.68 ± 4. 46 b | 1694.45 ± 353.29 b | 1305.56 ± 635.16 b | |
| Triploid progeny (3X) | 30.45 ± 9.04 a | 23.83 ± 4.62 c | 942.53 ± 763.14 b | 1438.96 ± 755.84 ab | |
| Diploid progeny (2X) | 33.66 ± 5.73 a | 29.21 ± 5.38 d | 925.37 ± 551.20 b | 2118.60 ± 823.72 a | |
| Traits | Ploidy | Years | CV/% | Variation Range | MPH/% | RH/% | RL/% |
|---|---|---|---|---|---|---|---|
| Pollen activity/% | Triploid progeny | 2022 | 29.69 | 15.13–46.90 | 10.41 | 24.49 | 4.08 |
| 2023 | 19.39 | 13.40–35.52 | −5.77 | 2.13 | 8.51 | ||
| Diploid progeny | 2022 | 17.02 | 22.7–44.53 | 22.04 | 30.77 | 0 | |
| 2023 | 18.42 | 23.23–40.41 | 15.50 | 21.74 | 0 | ||
| Pollen quantity/grain | Triploid progeny | 2022 | 80.97 | 0.00–37.50 | −47.39 | 4.92 | 68. 85 |
| 2023 | 52.53 | 0.00–3377.78 | −4.07 | 33.33 | 40.74 | ||
| Diploid progeny | 2022 | 59.57 | 16.67–1888.89 | −48.35 | 0 | 60.00 | |
| 2023 | 38.88 | 0.00–4416.67 | 41.24 | 75.00 | 12.50 |
| Years | Shrunk Rate/% | n Pollen Rate/% | 2n Pollen Rate/% | 3n and Above Pollen Rate/% | ||
|---|---|---|---|---|---|---|
| Parents | 2022 | DZ (2X) | 2.00 | 92.67 | 5.33 | 0.00 |
| CG (4X) | 0.00 | 50.00 | 49.00 | 1.00 | ||
| 2023 | DZ (2X) | 9.33 | 89.00 | 1.67 | 0.00 | |
| CG (4X) | 0.00 | 50.67 | 46.67 | 2.67 | ||
| Hybrid progeny | 2022 | Triploid progeny | 1.75 ± 2.12 * | 74.16 ± 14.10 ** | 22.52 ± 11.59 ** | 1.57 ± 4.88 |
| Diploid progeny | 3.06 ± 2.49 | 85.64 ± 10.35 | 11.10 ± 10.47 | 0.19 ± 0.67 | ||
| 2023 | Triploid progeny | 3.00 ± 4.06 | 89.13 ± 7.70 ** | 7.64 ± 7.82 ** | 0.23 ± 0.48 ** | |
| Diploid progeny | 2.17 ± 1.60 | 95.39 ± 2.48 | 2.44 ± 2.35 | 0.00 ± 0.00 |
| Years | Type | Diploid | Triploid | ||||
|---|---|---|---|---|---|---|---|
| Minimum Rate/% | Maximum Rate/% | CV/% | Minimum Rate/% | Maximum Rate/% | CV/% | ||
| 2022 | Shrunk pollen rate | 0.00 | 11.82 | 81.37 | 0.00 | 10.85 | 121.14 |
| n pollen rate | 47.91 | 96.07 | 12.09 | 12.55 | 94.00 | 19.01 | |
| 2n pollen rate | 0.44 | 46.25 | 94.32 | 5.00 | 52.16 | 51.47 | |
| 3n and above pollen rate | 0.00 | 2.92 | 352.63 | 0.00 | 35.29 | 310.83 | |
| 2023 | Shrunk pollen rate | 0.00 | 6.33 | 73.73 | 0.00 | 17.67 | 135.33 |
| n pollen rate | 91.33 | 98.67 | 2.60 | 69.17 | 99.13 | 8.64 | |
| 2n pollen rate | 0.00 | 8.33 | 96.31 | 0.00 | 28.95 | 102.36 | |
| 3n and above pollen rate | 0.00 | 0.00 | 0.00 | 0.00 | 2.64 | 208.70 | |
| Name | 2n Pollen Rate/% | Pollen Activity/% | Pollen Amount/Grain |
|---|---|---|---|
| Q20 | 12.26 | 19.61 | 1094.44 |
| Q35 | 8.05 | 29.55 | 636.11 |
| Q36 | 9.17 | 21.30 | 950.00 |
| Q40 | 5.96 | 24.04 | 697.22 |
| Q41 | 8.48 | 30.70 | 827.78 |
| Q92 | 7.30 | 26.00 | 736.11 |
| Year | Pollen Diameter/μm | |||||||
|---|---|---|---|---|---|---|---|---|
| Parents | Pollen Type Grading Scale (P) | |||||||
| Mean | 1.3 Times | 1.5 Times | Shrunk Pollen | n Pollen | 2n Pollen | 3n and Above Pollen | ||
| 2022 | DZ | 21.45 | 27.89 | 32.18 | ≤15.00 | 15.00 < P ≤ 25.99 | 25.99 < P ≤ 33.78 | >33.78 |
| CG | 25.99 | 33.78 | 38.98 | |||||
| 2023 | DZ | 20.14 | 26.19 | 30.21 | ≤15.00 | 15.00 < P ≤ 28.62 | 28.62 < P ≤ 37.21 | >37.21 |
| CG | 28.62 | 37.21 | 42.93 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, Y.; Ge, L.; Song, Y.; Luo, Z.; Wang, J.; Liu, M.; Yan, F. Analysis of Meiotic Behavior and 2n Pollen Formation Frequency in Triploid Hybrids of Chinese Jujube. Plants 2025, 14, 1643. https://doi.org/10.3390/plants14111643
Zhong Y, Ge L, Song Y, Luo Z, Wang J, Liu M, Yan F. Analysis of Meiotic Behavior and 2n Pollen Formation Frequency in Triploid Hybrids of Chinese Jujube. Plants. 2025; 14(11):1643. https://doi.org/10.3390/plants14111643
Chicago/Turabian StyleZhong, Yunxi, Lixin Ge, Yinfang Song, Zhi Luo, Jiurui Wang, Mengjun Liu, and Fenfen Yan. 2025. "Analysis of Meiotic Behavior and 2n Pollen Formation Frequency in Triploid Hybrids of Chinese Jujube" Plants 14, no. 11: 1643. https://doi.org/10.3390/plants14111643
APA StyleZhong, Y., Ge, L., Song, Y., Luo, Z., Wang, J., Liu, M., & Yan, F. (2025). Analysis of Meiotic Behavior and 2n Pollen Formation Frequency in Triploid Hybrids of Chinese Jujube. Plants, 14(11), 1643. https://doi.org/10.3390/plants14111643