
Development and Characterization of Synthetic Allotetraploids Between Diploid Species Gossypium herbaceum and Gossypium nelsonii for Cotton Genetic Improvement

 ,
,  ,
,  , , , , , , , , , , , , , , and
, , , , , , , , , , , , , , and 
Abstract
1. Introduction
2. Results
2.1. Fertility of Parental Lines and Interspecific Hybrids: Boll- and Seed-Setting
2.2. Inheritance of Morpho-Biological Traits in Interspecific Polyploid Hybrids
2.2.1. Vegetative Growth Duration of Parental Lines and Polyploid Hybrids
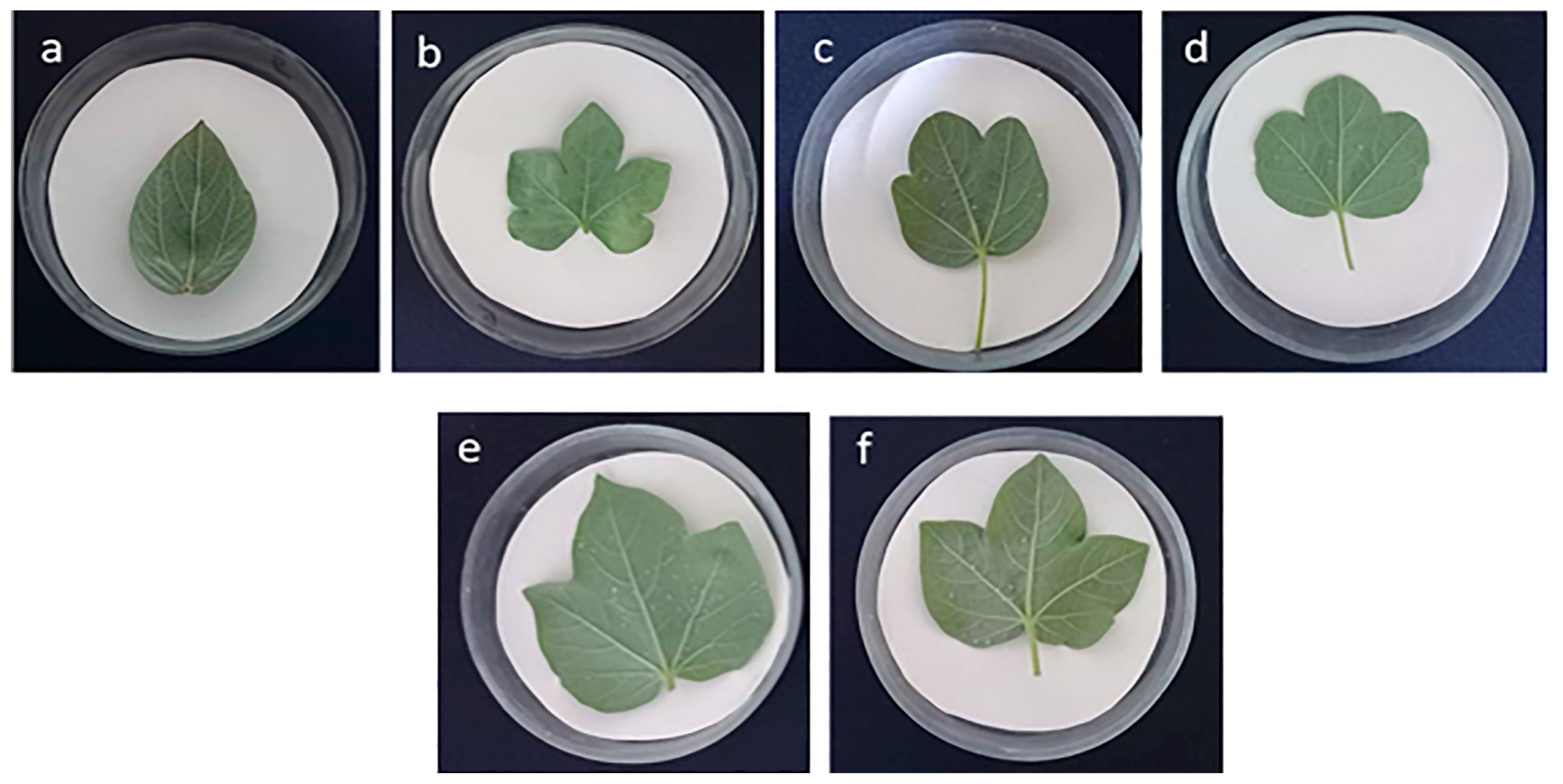
2.2.2. Morphological Description of the F1C Allotetraploid Hybrids
2.2.3. Inheritance and Improvement of Fiber Traits in Synthetic Hybrids
2.3. Wild Species-Derived Insect Pest Resistance in Synthetic Allotetraploids
2.3.1. Resistance to Cotton Aphids (Aphis gossypii Glover.)
2.3.2. Resistance to Whiteflies (Aleyrodidae)
2.4. Cytogenetic Analysis of Interspecific Hybrids
2.4.1. Cytogenetic Screening of Polyploid Hybrids
2.4.2. Pollen Fertility
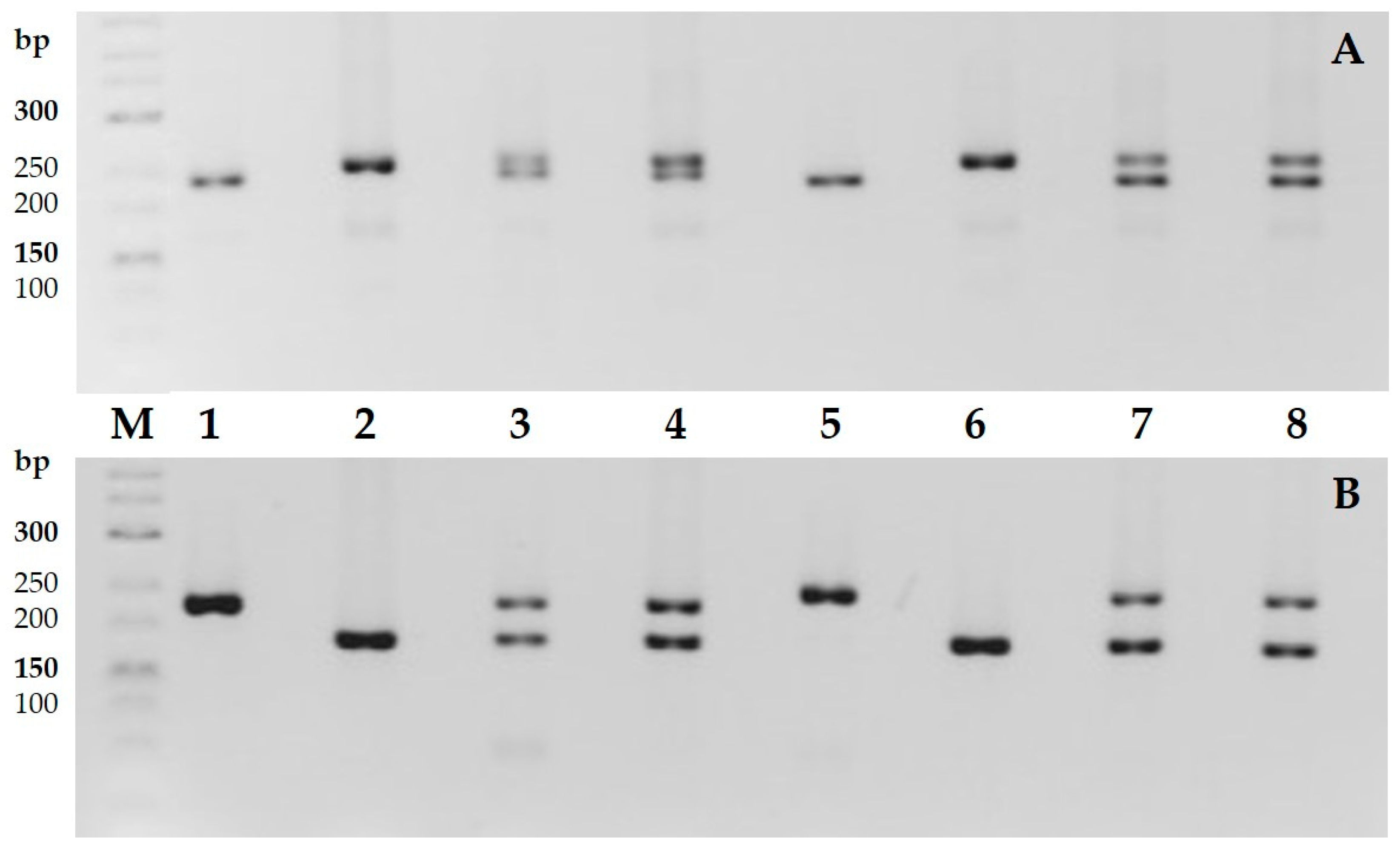
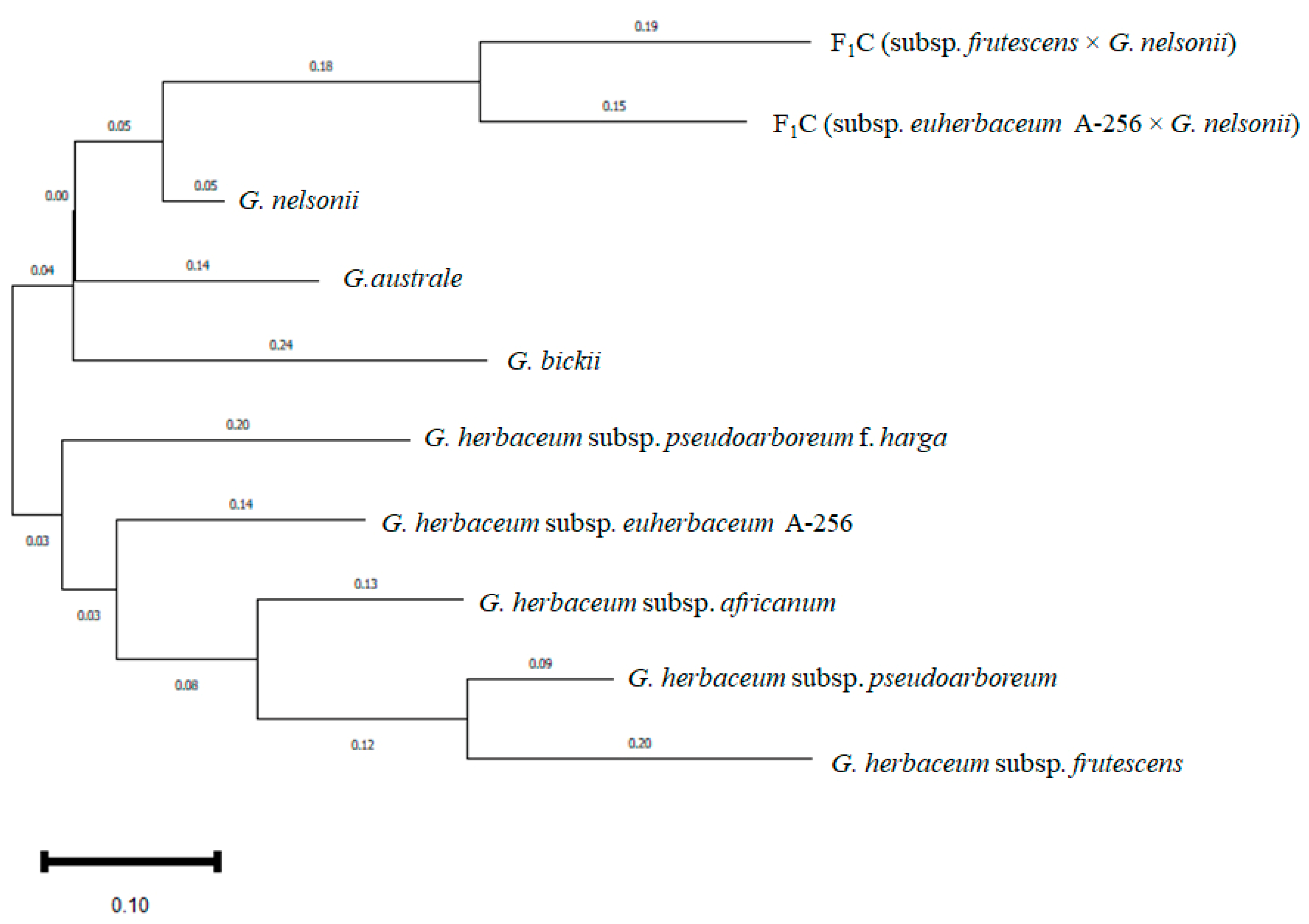
2.4.3. Molecular Genetic and Phylogenetic Analysis of the Intraspecific Hybrids
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Hybridization and Polyploidization
4.3. Phenological Observations
4.4. Insect Pest Resistance Evaluation
4.4.1. Experimental Design
4.4.2. Insect Infestation Assays
- Aphid colonies were maintained on susceptible cotton plants under greenhouse conditions.
- For experiments, 10 aphids were transferred to each test plant using a fine brush.
- Infestation severity was assessed every 48 h for 21 days using a standardized 0–5 scale:
- 4.
- Data from 10 replicates per genotype were used to calculate mean infestation percentages (Table 4).
- 1.
- Whitefly adults were collected from infected field plants and maintained in insectaries.
- 2.
- Test plants were infested with 15 adult whiteflies per plant, confined in mesh cages.
- 3.
- Damage assessment included:
- -
- Percentage of the chlorotic leaf area;
- -
- Nymph density per cm2 of leaf surface;
- -
- Overall plant vigor score (1–5 scale).
- 4.
- The results were statistically analyzed as described in Section 4.7 and presented in Table 4.
- -
- All the experiments included a randomized complete block design;
- -
- Insect populations were standardized by age and size before inoculation;
- -
- Environmental conditions were monitored continuously using data loggers;
- -
- Control genotype (cv. Ravnak-1) was included in each experimental run.
4.5. Cytological Analysis
4.6. Genomic DNA Isolation and SSR Marker Analysis
4.7. Statistical Analysis
4.8. Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kushanov, F.; Turaev, O.; Ernazarova, D.; Gapparov, B.; Oripova, B.; Kudratova, M.; Rafieva, F.; Khalikov, K.; Erjigitov, D.; Khidirov, M.; et al. Genetic diversity, QTL mapping, and marker-assisted selection technology in cotton (Gossypium spp.). Front. Plant Sci. 2021, 12, 779386. [Google Scholar] [CrossRef] [PubMed]
- Safiullina, A.K.; Ernazarova, D.K.; Turaev, O.S.; Rafieva, F.U.; Ernazarova, Z.A.; Arslanova, S.K.; Toshpulatov, A.K.; Oripova, B.B.; Kudratova, M.K.; Khalikov, K.K.; et al. Genetic diversity and subspecific races of upland cotton (Gossypium hirsutum L.). Genes 2024, 15, 1533. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Wendel, J.; Hua, J. Designations for individual genomes and chromosomes in Gossypium. J. Cotton Res. 2018, 1, 3. [Google Scholar] [CrossRef]
- Wendel, J.F.; Grover, C.E. Taxonomy and evolution of the cotton genus, Gossypium. Cotton 2015, 57, 25–44. [Google Scholar] [CrossRef]
- Kushanov, F.N.; Komilov, D.J.; Turaev, O.S.; Ernazarova, D.K.; Amanboyeva, R.S.; Gapparov, B.M.; Yu, J.Z. Genetic analysis of mutagenesis that induces the photoperiod insensitivity of wild cotton Gossypium hirsutum subsp. purpurascens. Plants 2022, 11, 3012. [Google Scholar] [CrossRef] [PubMed]
- Mithil, A.; Kumar, S.; Patel, R. Comparative analysis of upland and pima cotton fiber quality. J. Cotton Res. 2016, 5, 123–135. [Google Scholar]
- Zhou, T.; Wang, N.; Wang, Y.; Zhang, X.L.; Li, B.G.; Li, W.; Su, J.J.; Wang, C.X.; Zhang, A.; Ma, X.F.; et al. Nucleotide evolution, domestication selection, and genetic relationships of chloroplast genomes in the economically important crop genus Gossypium. Front. Plant Sci. 2022, 13, 873788. [Google Scholar] [CrossRef]
- Joshi, B.; Singh, S.; Tiwari, G.J.; Kumar, H.; Boopathi, N.M.; Jaiswal, S.; Adhikari, D.; Kumar, D.; Sawant, S.V.; Iquebal, M.A.; et al. Genome-wide association study of fiber yield-related traits uncovers the novel genomic regions and candidate genes in Indian upland cotton (Gossypium hirsutum L.). Front. Plant Sci. 2023, 14, 1252746. [Google Scholar] [CrossRef]
- Wendel, J.F.; Cronn, R.C. Polyploidy and the evolutionary history of cotton. Adv. Agron. 2003, 78, 139–186. [Google Scholar] [CrossRef]
- Wendel, J.F.; Brubaker, C.L.; Seelanan, T. The Origin and Evolution of Gossypium in Physiology of Cotton; Stewart, J.M., Oosterhuis, D.M., Heitholt, J.J., Mauney, J.R., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 1–18. [Google Scholar]
- Huang, G.; Huang, J.Q.; Chen, X.Y.; Zhu, Y.X. Recent advances and future perspectives in cotton research. Annu. Rev. Plant Biol. 2021, 72, 437–462. [Google Scholar] [CrossRef]
- Kulkarni, V.N.; Khadi, B.M.; Maralappanavar, M.S.; Deshapande, L.A.; Narayanan, S.S. The worldwide gene pools of Gossypium arboreum L. and G. herbaceum L.; and their improvement. In Genetics and Genomics of Cotton; Paterson, A.H., Ed.; Springer: New York, NY, USA, 2009; pp. 61–100. [Google Scholar] [CrossRef]
- Ranjan, A.; Nigam, D.; Asif, M.H.; Singh, R.; Ranjan, S.; Mantri, S.; Pandey, N.; Trivedi, I.; Rai, K.M.; Jena, S.N.; et al. Genome-wide expression profiling of two accessions of G. herbaceum L. in response to drought. BMC Genom. 2012, 13, 94. [Google Scholar] [CrossRef] [PubMed]
- Ramaraj, T.; Grover, C.E.; Mendoza, A.C.; Arick, M.A.; Jareczek, J.J.; Leach, A.G.; Peterson, D.G.; Wendel, J.F.; Udall, J.A. The Gossypium herbaceum L. Wagad genome as a resource for understanding cotton domestication. G3 2023, 13, jkac308. [Google Scholar] [CrossRef] [PubMed]
- Çınar, V.M.; Ünay, A. Combining suitable brown lint color, fiber quality, and yield in F2 cotton hybrids. Ind. Crops Prod. 2025, 224, 119674. [Google Scholar] [CrossRef]
- Naoumkina, M.; Hinchliffe, D.J.; Thyssen, G.N. Naturally, colored cotton for wearable applications. Front. Plant Sci. 2024, 15, 1350405. [Google Scholar] [CrossRef] [PubMed]
- Khidirov, M.T.; Ernazarova, D.K.; Rafieva, F.U.; Ernazarova, Z.A.; Toshpulatov, A.K.; Umarov, R.F.; Kholova, M.D.; Oripova, B.B.; Kudratova, M.K.; Gapparov, B.M.; et al. Genomic and cytogenetic analysis of synthetic polyploids between diploid and tetraploid cotton (Gossypium) species. Plants 2023, 12, 4184. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Zhan, R.; He, Y.; Song, S.; Wang, L.; Ge, Y.; Chen, D. Morphological description of a novel synthetic allotetraploid (A1A1G3G3) of Gossypium herbaceum L. and G. nelsonii Fryx. suitable for disease-resistant breeding applications. PLoS ONE 2020, 15, e0242620. [Google Scholar] [CrossRef]
- Anwar, M.; Iqbal, M.Z.; Abro, A.A.; Memon, S.; Bhutto, L.A.; Memon, S.A.; Peng, Y. Inter-specific hybridization in cotton (Gossypium hirsutum) for crop improvement. Agronomy 2022, 12, 3158. [Google Scholar] [CrossRef]
- Lavinscky, M.P.; Souza, M.M.; Silva, G.S.; Melo, C.A.F. Contributions of classical and molecular cytogenetic in meiotic analysis and pollen viability for plant breeding. Genet. Mol. Res. 2017, 16, gmr16039582. [Google Scholar] [CrossRef]
- Muramoto, H. Hexaploid cotton: Some plant and fiber properties. Crop Sci. 1969, 9, 27–29. [Google Scholar] [CrossRef]
- Endrizzi, J.E.; Phillips, L.L. A hybrid between Gossypium arboreum L. and G. raimondii Ulb. Can. J. Genet. Cytol. 1960, 2, 311–319. [Google Scholar] [CrossRef]
- Li, B.; Zhang, X.R. Studies on the hybrid between G. arboreum L. and G. bickii Prokh. Acta Genet. Sin. 1987, 14, 121–126. [Google Scholar]
- Sun, Y.Q.; Zhang, X.L.; Nie, Y.C.; Guo, X.P.; Jin, S.X.; Liang, S.G. Production and characterization of somatic hybrids between upland cotton (Gossypium hirsutum) and wild cotton (G. klotzschianum Anderss) via electrofusion. Theor. Appl. Genet. 2004, 109, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Masoomi-Aladizgeh, F.; Najeeb, U.; Hamzelou, S.; Pascovici, D.; Amirkhani, A.; Tan, D.K.; Atwell, B.J. Pollen development in cotton (Gossypium hirsutum) is highly sensitive to heat exposure during the tetrad stage. Plant Cell Environ. 2021, 44, 2150–2166. [Google Scholar] [CrossRef]
- Abdurakhmonov, I.Y.; Kohel, R.J.; Yu, J.Z.; Pepper, A.E.; Abdullaev, A.A.; Kushanov, F.N.; Salakhutdinov, I.B.; Buriev, Z.T.; Saha, S.; Scheffler, B.E.; et al. Molecular diversity and association mapping of fiber quality traits in exotic G. hirsutum L. germplasm. Genomics 2008, 92, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Arif, T.; Chaudhary, M.T.; Majeed, S.; Rana, I.A.; Ali, Z.; Elansary, H.O.; Moussa, I.M.; Sun, S.; Azhar, M.T. Exploitation of various physio-morphological and biochemical traits for the identification of drought-tolerant genotypes in cotton. BMC Plant Biol. 2023, 23, 508. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, Y.; Chen, Y.; Wang, Y.; Chen, J.; Zhang, T.; Zhou, B. A New Synthetic Allotetraploid (A1A1G2G2) between Gossypium herbaceum and G. australe: Bridging for Simultaneously Transferring Favorable Genes from These Two Diploid Species into Upland Cotton. PLoS ONE 2015, 10, e0123209. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Y.Y.; Wang, K.; Zhu, X.F.; Guo, W.Z.; Zhang, T.Z.; Zhou, B. Construction of a complete set of alien chromosome addition lines from Gossypium australe in Gossypium hirsutum: Morphological, cytological, and genotypic characterization. Theor. Appl. Genet. 2014, 127, 1105–1121. [Google Scholar] [CrossRef]
- Wang, Y.; Feng, S.; Li, S.; Tang, D.; Chen, Y.; Chen, Y.; Zhou, B. Inducement and identification of chromosome introgression and translocation of Gossypium australe on Gossypium hirsutum. BMC Genom. 2018, 19, 15. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.H.; Brubaker, C.L.; Wendel, J.F. A rapid method for extraction of cotton (Gossypium spp.) genomic DNA suitable for RFLP or PCR analysis. Plant Mol. Biol. Rep. 1993, 11, 122–127. [Google Scholar] [CrossRef]
- Maxwell, S.E.; Delaney, H.D. Designing Experiments and Analyzing Data: A Model Comparison Perspective, 3rd ed.; Routledge: New York, NY, USA, 2003. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Yang, Z. Phylogenetic analysis using parsimony and likelihood methods. J. Mol. Evol. 1996, 42, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-Q.; Xu, X.-J.; Dong, N.; Ai, N.-J.; Wang, Q.-L. Association mapping identifies markers related to major early-maturating traits in upland cotton (Gossypium hirsutum L.). Plant Breed. 2016, 135, 483–491. [Google Scholar] [CrossRef]
- Shao, Q.; Zhang, F.; Tang, S.; Liu, Y.; Fang, X.; Liu, D.; Zhang, Z. Identifying QTL for fiber quality traits with three upland cotton (Gossypium hirsutum L.) populations. Euphytica 2014, 198, 43–58. [Google Scholar] [CrossRef]
- Shang, L.; Liang, Q.; Wang, Y.; Wang, X.; Wang, K.; Abduweli, A.; Ma, L.; Cai, S.; Hua, J. Identification of stable QTLs controlling fiber traits properties in multi-environment using recombinant inbred lines in Upland cotton (Gossypium hirsutum L.). Euphytica 2015, 205, 877–888. [Google Scholar] [CrossRef]
- Pan, Z.-E.; Sun, J.; Jia, Y.; Zhou, Z.; Pang, B.; Du, X. Screening of SSR core primers with polymorphism on a cotton panel. J. Biodivers. Sci. 2008, 16, 555–561. [Google Scholar] [CrossRef]
- Yu, J.; Ulloa, M.; Hoffman, S.; Kohel, R.; Pepper, A.; Fang, D.; Percy, R.; Burke, J. Mapping genomic loci for cotton plant architecture, yield components, and fiber properties in an interspecific (Gossypium hirsutum L. × Gossypium barbadense L.) RIL population. Mol. Gen. Genom. 2014, 15, 1347–1367. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Chen, B.; Xu, X.; Li, D.; Dong, J. Simple sequence repeat markers associated/linked with agronomic traits, as core primers, are eminently suitable for DNA fingerprinting in Upland cotton. Breed. Sci. 2018, 68, 393–403. [Google Scholar] [CrossRef]
- Zhang, X.; Zhai, C.; He, L.; Guo, Q.; Zhang, X.; Xu, P.; Su, H.; Gong, Y.; Ni, W.; Shen, X. Morphological, cytological and molecular analyses of a synthetic hexaploid derived from an interspecific hybrid between Gossypium hirsutum and Gossypium anomalum. Crop J. 2014, 2, 272–277. [Google Scholar] [CrossRef]
- Fang, D.D.; Zeng, L.; Thyssen, G.N.; Delhom, C.D.; Bechere, E.; Jones, D.C.; Li, P. Stability and transferability assessment of the cotton fiber strength QTL qFS-c7-1 on chromosome A07. Crop J. 2021, 9, 380–386. [Google Scholar] [CrossRef]
- Kushanov, F.N.; Buriev, Z.T.; Shermatov, S.E.; Turaev, O.S.; Norov, T.M.; Pepper, A.E.; Saha, S.; Ulloa, M.; Yu, J.Z.; Jenkins, J.N.; et al. QTL mapping for flowering-time and photoperiod insensitivity of cotton Gossypium darwinii Watt. PLoS ONE 2017, 12, e0186240. [Google Scholar] [CrossRef]
- Jamshed, M.; Jia, F.; Gong, J.; Palanga, K.; Shi, Y.; Li, J.; Shang, H.; Liu, A.; Chen, T.; Zhang, Z.; et al. Identification of stable quantitative trait loci (QTLs) for fiber quality traits across multiple environments in G. hirsutum recombinant inbred line population. BMC Genom. 2016, 17, 197. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhao, Y.; Wang, Y.; Shang, L.; Hua, J. QTLs analysis and validation for fiber quality traits using maternal backcross population in Upland cotton. Front. Plant Sci. 2017, 8, 2168. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, J.; Ma, J.; Tang, Y.; Liu, D.; Teng, Z. Genetic mapping and quantitative trait locus analysis of fiber quality traits using a three-parent composite population in upland cotton (Gossypium hirsutum L.). Mol. Breed. 2012, 29, 335–348. [Google Scholar] [CrossRef]
- Osman, A.; Gutierrez, O.A.; Robinson, J.N.; Jenkins, J.C.; McCarty, J.M.; Wubben, F.E.; Callahan, R.L.; Nichols, R.L. Identification of QTL regions and SSR markers associated with resistance to reniform nematode in Gossypium barbadense L. accession GB713. Theor. Appl. Genet. 2011, 122, 673–683. [Google Scholar] [CrossRef]
- Wang, B.; Zhuang, Z.; Zhang, Z.; Draye, X.; Shuang, L.-S.; Shehzad, T.; Lubbers, E.L.; Jones, D.; May, O.L.; Paterson, A.H.; et al. Advanced Backcross QTL Analysis of Fiber Strength and Fineness in a Cross between Gossypium hirsutum and Gossypium mustelinum. Front. Plant Sci. 2017, 8, 1848. [Google Scholar] [CrossRef]
- Liang, Q.; Hu, C.; Hua, H.; Li, Z.; Hua, J. Construction of a linkage map and QTL mapping for fiber quality traits in upland cotton (Gossypium hirsutum L.). Chin. Sci. Bull. 2013, 58, 3233–3243. [Google Scholar] [CrossRef]
- Liang, Q.; Shang, L.; Wang, Y.; Hua, J. Partial dominance, overdominance and epistasis as the genetic basis of heterosis in Upland cotton (Gossypium hirsutum L.). PLoS ONE 2015, 10, e0143548. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yuan, Y.; Wei, Z.; Guo, X.; Guo, Y.; Zhang, S.; Zhao, J.; Zhang, G.; Song, X.; Sun, X. Molecular Mapping and Validation of a Major QTL Conferring Resistance to a Defoliating Isolate of Verticillium Wilt in Cotton (Gossypium hirsutum L.). PLoS ONE 2014, 9, e96226. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, B.; Zhao, N.; Kong, J.; Hua, J. Fiber quality improvement in upland cotton (Gossypium hirsutum L.): Quantitative trait loci mapping and marker assisted selection application. Front. Plant Sci. 2019, 10, 1585. [Google Scholar] [CrossRef]
- Li, H.; Pan, Z.; He, S.; Jia, Y.; Geng, X.; Chen, B.; Wang, L.; Pang, B.; Du, X. QTL mapping of agronomic and economic traits for four F2 populations of upland cotton. J. Cotton Res. 2021, 4, 3. [Google Scholar] [CrossRef]
- Yu, Y.; Yuan, D.; Liang, S.; Li, X.; Wang, X.; Lin, Z.; Zhang, X. Genome structure of cotton revealed by a genome-wide SSR genetic map constructed from a BC1 population between Gossypium hirsutum and Gossypium barbadense. BMC Genom. 2011, 12, 15. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ulloa, M.; Duong, T.; Roberts, P.A. Quantitative trait loci mapping of multiple independent loci for resistance to Fusarium oxysporum f. sp. vasinfectum races 1 and 4 in an interspecific cotton population. Phytopathology 2018, 108, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Mei, H.; Wang, S.; Li, X.; Zhu, X.; Zhang, T. Association mapping of seed oil and protein contents in upland cotton. Euphytica 2015, 205, 637–645. [Google Scholar] [CrossRef]
- Shang, L.G.; Liu, F.; Wang, Y.M.; Abduweli, A.; Cai, S.H.; Wang, K.B.; Hua, J.P. Dynamic QTL mapping for plant height in Upland cotton (Gossypium hirsutum). Plant Breed. 2015, 134, 703–712. [Google Scholar] [CrossRef]
- Shang, L.; Wang, Y.; Wang, X.; Liu, F.; Abduweli, A.; Cai, S.; Li, Y.; Ma, L.; Wang, K.; Hua, J. Genetic Analysis and QTL Detection on Fiber Traits Using Two Recombinant Inbred Lines and Their Backcross Populations in Upland Cotton. G3 2016, 6, 2717–2724. [Google Scholar] [CrossRef]
- Yu, J.Z.; Kohel, R.J.; Fang, D.D.; Cho, J.; Van, A.D.; Ulloa, M.; Hoffman, S.M.; Pepper, A.E.; Stelly, D.M.; Jenkins, J.N.; et al. A high-density simple sequence repeats and single nucleotide polymorphism genetic map of the tetraploid cotton genome. G3 2012, 2, 43–58. [Google Scholar] [CrossRef]
- Huang, G.; Wu, Z.; Percy, R.G.; Bai, M.; Li, Y.; Frelichowski, J.E.; Zhu, Y. Genome sequence of Gossypium herbaceum and genome updates of Gossypium arboreum and Gossypium hirsutum provide insights into cotton A-genome evolution. Nat. Genet. 2020, 52, 516–524. [Google Scholar] [CrossRef]
- Guo, W.Z.; Cai, C.P.; Wang, C.B.; Han, Z.G.; Song, X.; Wang, K.; Niu, X.; Wang, C.; Lu, K.; Shi, B.; et al. A microsatellite-based, gene-rich linkage map reveals genome structure, function, and evolution in Gossypium. Genetics 2007, 176, 527–541. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Cai, C.; Wu, S.; Zhang, F.; Hou, S.; Guo, W. Evaluation and exploration of favorable QTL alleles for salt stress related traits in cotton cultivars (G. hirsutum L.). PLoS ONE 2016, 11, e0151076. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, H.; Chen, W.; Li, Y. Genetic Structure, Linkage Disequilibrium and Association Mapping of Verticillium Wilt Resistance in Elite Cotton (Gossypium hirsutum L.) Germplasm Population. PLoS ONE 2014, 9, e86308. [Google Scholar] [CrossRef]
- Hoffman, S.M.; Yu, J.Z.; Grum, D.S.; Xiao, J.; Kohel, R.J.; Pepper, A.E. Identification of 700 new microsatellite loci from cotton (G. hirsutum L.). J. Cotton Sci. 2007, 11, 208–241. [Google Scholar]
- Wu, J.; Gutierrez, O.A.; Jenkins, J.N.; McCarty, J.C.; Zhu, J. Quantitative analysis and QTL mapping for agronomic and fiber traits in an RI population of upland cotton. Euphytica 2009, 165, 231–245. [Google Scholar] [CrossRef]
- Abdurakhmonov, I.Y.; Saha, S.; Jenkins, J.N.; Buriev, Z.T.; Shermatov, S.E.; Scheffler, B.E.; Pepper, A.E.; Yu, J.Z.; Kohel, R.Z.; Abdukarimov, A. Linkage disequilibrium based association mapping of fiber quality traits in G. hirsutum L. variety germplasm. Genetica 2009, 136, 401–417. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Fang, X.; Tang, S.; Zhang, J.; Liu, D.; Teng, Z.; Li, L.; Ni, H.; Zheng, F.; Liu, D.; et al. Genetic map and QTL controlling fibre quality traits in upland cotton (Gossypium hirsutum L.). Euphytica 2015, 203, 615–628. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, D.; Zhu, S.; Zhang, L.; Li, L. A New Synthetic Hybrid (A1D5) between Gossypium herbaceum and G. raimondii and Its Morphological, Cytogenetic, Molecular Characterization. PLoS ONE 2017, 12, e0169833. [Google Scholar] [CrossRef]
- Wang, C.; Ulloa, M.; Duong, T.T.; Roberts, P.A. QTL Analysis of Transgressive Nematode Resistance in Tetraploid Cotton Reveals Complex Interactions in Chromosome 11 Regions. Front. Plant Sci. 2017, 8, 1979. [Google Scholar] [CrossRef]
 —diploid G. herbaceum subsp. frutescens;
—diploid G. herbaceum subsp. frutescens;  —diploid G. nelsonii; F1—diploid first generation; F1C—allotetraploid hybrid progeny of the first generation. (B) —diploid G. herbaceum subsp. euherbaceum (cv. А-256); —diploid G. nelsonii; F1—diploid first generation; F1C—allotetraploid hybrid progeny of the first generation.
—diploid G. herbaceum subsp. frutescens; —diploid G. nelsonii; F1—diploid first generation; F1C—allotetraploid hybrid progeny of the first generation. (B) —diploid G. herbaceum subsp. euherbaceum (cv. А-256); —diploid G. nelsonii; F1—diploid first generation; F1C—allotetraploid hybrid progeny of the first generation.
—diploid G. nelsonii; F1—diploid first generation; F1C—allotetraploid hybrid progeny of the first generation. (B) —diploid G. herbaceum subsp. euherbaceum (cv. А-256); —diploid G. nelsonii; F1—diploid first generation; F1C—allotetraploid hybrid progeny of the first generation.
—diploid G. herbaceum subsp. frutescens; —diploid G. nelsonii; F1—diploid first generation; F1C—allotetraploid hybrid progeny of the first generation. (B) —diploid G. herbaceum subsp. euherbaceum (cv. А-256); —diploid G. nelsonii; F1—diploid first generation; F1C—allotetraploid hybrid progeny of the first generation. ) diploid maternal genotype; (B) diploid first generation—(F1) G. herbaceum subsp. frutescens × G. nelsonii; (C) allotetraploid hybrid progeny of the first generation—(F1C) G. herbaceum subsp. frutescens × G. nelsonii; (D) G. nelsonii—() diploid paternal genotype.
) diploid maternal genotype; (B) diploid first generation—(F1) G. herbaceum subsp. frutescens × G. nelsonii; (C) allotetraploid hybrid progeny of the first generation—(F1C) G. herbaceum subsp. frutescens × G. nelsonii; (D) G. nelsonii—() diploid paternal genotype.
) diploid maternal genotype; (B) diploid first generation—(F1) G. herbaceum subsp. frutescens × G. nelsonii; (C) allotetraploid hybrid progeny of the first generation—(F1C) G. herbaceum subsp. frutescens × G. nelsonii; (D) G. nelsonii—() diploid paternal genotype.
) diploid maternal genotype; (B) diploid first generation—(F1) G. herbaceum subsp. frutescens × G. nelsonii; (C) allotetraploid hybrid progeny of the first generation—(F1C) G. herbaceum subsp. frutescens × G. nelsonii; (D) G. nelsonii—() diploid paternal genotype. ) diploid maternal genotype; (B) diploid first generation—(F1) G. herbaceum subsp. euherbaceum cv. А-256 × G. nelsonii; (C) allotetraploid hybrid progeny of the first generation—(F1C) G. herbaceum subsp. euherbaceum cv. А-256 × G. nelsonii; (D) G. nelsonii—() diploid paternal genotype.
) diploid maternal genotype; (B) diploid first generation—(F1) G. herbaceum subsp. euherbaceum cv. А-256 × G. nelsonii; (C) allotetraploid hybrid progeny of the first generation—(F1C) G. herbaceum subsp. euherbaceum cv. А-256 × G. nelsonii; (D) G. nelsonii—() diploid paternal genotype.
) diploid maternal genotype; (B) diploid first generation—(F1) G. herbaceum subsp. euherbaceum cv. А-256 × G. nelsonii; (C) allotetraploid hybrid progeny of the first generation—(F1C) G. herbaceum subsp. euherbaceum cv. А-256 × G. nelsonii; (D) G. nelsonii—() diploid paternal genotype.
) diploid maternal genotype; (B) diploid first generation—(F1) G. herbaceum subsp. euherbaceum cv. А-256 × G. nelsonii; (C) allotetraploid hybrid progeny of the first generation—(F1C) G. herbaceum subsp. euherbaceum cv. А-256 × G. nelsonii; (D) G. nelsonii—() diploid paternal genotype. —G. herbaceum subsp. frutescens, —diploid G. nelsonii, F1C—synthetic hybrid of G. herbaceum subsp. frutescens × G. nelsonii combination; (B) —F1C synthetic hybrid of G. herbaceum subsp. frutescens × G. nelsonii combination, —G. hirsutum cv. Ravnak-1, F1—first generation hybrid of [(G. herbaceum subsp. frutescens × G. nelsonii) × G. hirsutum cv. Ravnak-1]; (C) —F1 hybrid of [(G. herbaceum subsp. frutescens × G. nelsonii) × G. hirsutum cv. Ravnak-1], —G. hirsutum cv. Ravnak-1, BC1F1—backcross hybrid (BC1F1) [(G. herbaceum subsp. frutescens × G. nelsonii) × G. hirsutum cv. Ravnak-1]. Note: The
—G. herbaceum subsp. frutescens, —diploid G. nelsonii, F1C—synthetic hybrid of G. herbaceum subsp. frutescens × G. nelsonii combination; (B) —F1C synthetic hybrid of G. herbaceum subsp. frutescens × G. nelsonii combination, —G. hirsutum cv. Ravnak-1, F1—first generation hybrid of [(G. herbaceum subsp. frutescens × G. nelsonii) × G. hirsutum cv. Ravnak-1]; (C) —F1 hybrid of [(G. herbaceum subsp. frutescens × G. nelsonii) × G. hirsutum cv. Ravnak-1], —G. hirsutum cv. Ravnak-1, BC1F1—backcross hybrid (BC1F1) [(G. herbaceum subsp. frutescens × G. nelsonii) × G. hirsutum cv. Ravnak-1]. Note: The  symbol denotes a hybridization (cross) between the respective parental genotypes.
—G. herbaceum subsp. frutescens, —diploid G. nelsonii, F1C—synthetic hybrid of G. herbaceum subsp. frutescens × G. nelsonii combination; (B) —F1C synthetic hybrid of G. herbaceum subsp. frutescens × G. nelsonii combination, —G. hirsutum cv. Ravnak-1, F1—first generation hybrid of [(G. herbaceum subsp. frutescens × G. nelsonii) × G. hirsutum cv. Ravnak-1]; (C) —F1 hybrid of [(G. herbaceum subsp. frutescens × G. nelsonii) × G. hirsutum cv. Ravnak-1], —G. hirsutum cv. Ravnak-1, BC1F1—backcross hybrid (BC1F1) [(G. herbaceum subsp. frutescens × G. nelsonii) × G. hirsutum cv. Ravnak-1]. Note: The symbol denotes a hybridization (cross) between the respective parental genotypes.
symbol denotes a hybridization (cross) between the respective parental genotypes.
—G. herbaceum subsp. frutescens, —diploid G. nelsonii, F1C—synthetic hybrid of G. herbaceum subsp. frutescens × G. nelsonii combination; (B) —F1C synthetic hybrid of G. herbaceum subsp. frutescens × G. nelsonii combination, —G. hirsutum cv. Ravnak-1, F1—first generation hybrid of [(G. herbaceum subsp. frutescens × G. nelsonii) × G. hirsutum cv. Ravnak-1]; (C) —F1 hybrid of [(G. herbaceum subsp. frutescens × G. nelsonii) × G. hirsutum cv. Ravnak-1], —G. hirsutum cv. Ravnak-1, BC1F1—backcross hybrid (BC1F1) [(G. herbaceum subsp. frutescens × G. nelsonii) × G. hirsutum cv. Ravnak-1]. Note: The symbol denotes a hybridization (cross) between the respective parental genotypes.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Plant Samples | Number of Bolls | Number of Seeds per Boll | Complete Seeds per Boll, % | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Per Plant | Analyzed | Total | Complete | Empty | Range | S | V % | |||
| G. herbaceum subspecies and forms | ||||||||||
| 1. | G. herbaceum subsp. africanum | 28 | 10 | 15.0 | 13.0 | 2.0 | 88.0 ± 1.3 | 86.0–100.0 | 4.31 | 4.9 |
| 2. | G. herbaceum subsp. pseudoarboreum | 22 | 10 | 20.0 | 18.0 | 2.0 | 90.0 ± 1.2 | 85.0–100.0 | 3.94 | 4.3 |
| 3. | G. herbaceum subsp. pseudoarboreum f. harga | 42 | 10 | 16.0 | 14.0 | 2.0 | 90.0 ± 2.4 | 80.0–100.0 | 7.63 | 8.4 |
| 4. | G. herbaceum subsp. euherbaceum (cv. А-256) | 49 | 10 | 16.0 | 15.0 | 1.0 | 91.0 ± 2.5 | 75.0–100.0 | 8.15 | 8.9 |
| Australian wild species | ||||||||||
| 5. | G. australe | 41 | 10 | 11.0 | 10.0 | 1.0 | 94.5 ± 1.5 | 90.0–100 | 4.8 | 5.1 |
| 6. | G. nelsonii | 45 | 10 | 13.0 | 12.0 | 1.0 | 95.0 ± 1.3 | 91.0–100 | 4.2 | 4.4 |
| 7. | G. bickii | 43 | 10 | 9.0 | 8.0 | 1.0 | 93.0 ± 1.9 | 86.0–100 | 6.12 | 6.6 |
| No. | Hybrid Combinations | Number of Crosses | Number of Obtained Hybrid Bolls | Boll-Setting Rate, % | Seed-Setting Rate, % | |||
|---|---|---|---|---|---|---|---|---|
| Range | S | V | ||||||
| 1. | G. herbaceum subsp. africanum × G. australe | 38 | - | - | - | - | - | - |
| 2. | G. herbaceum subsp. africanum × G. bickii | 24 | - | - | - | - | - | - |
| 3. | G. herbaceum subsp. africanum × G. nelsonii | 9 | 1 | 11.1 | - | - | - | - |
| 4. | G. herbaceum subsp. pseudoarboreum × G. nelsonii | 47 | 10 | 21.3 | 62.4 ± 4.6 | 42.1–86.4 | 23.4 | 7.4 |
| 5. | G. herbaceum subsp. pseudoarboreum × G. australe | 18 | 5 | 27.8 | 60.4 ± 5.3 | 42.1–77.8 | 12.7 | 21.1 |
| 6. | G. herbaceum subsp. pseudoarboreum × G. bickii | 51 | 10 | 19.6 | 66.7 ± 4.7 | 47.6–85.0 | 22.3 | 7.1 |
| 7. | G. herbaceum subsp. pseudoarboreumf. harga × G. nelsonii | 22 | 6 | 27.3 | 54.3 ± 0.8 | 52.0–53.9 | 2.1 | 3.8 |
| 8. | G. herbaceum subsp. pseudoarboreum f. harga × G. bickii | 11 | 2 | 18.2 | 51.2 ± 3.8 | 47.4–55.0 | 5.4 | 10.5 |
| 9. | G. herbaceum subsp. frutescens × G. australe | 40 | - | - | - | - | - | - |
| 10. | G. herbaceum subsp. frutescens × G. nelsonii | 109 | 2 | 1.8 | 62.6 ± 7.5 | 55.2–70.0 | 10.5 | 16.7 |
| 11. | G. herbaceum subsp. frutescens × G. bickii | 108 | - | - | - | - | - | - |
| 12. | G. herbaceum subsp. euherbaceum (cv. А-256) × G. bickii | 19 | 1 | 5.3 | 50.0 | 50.0 | - | - |
| 13. | G. herbaceum subsp. euherbaceum (cv. А-256) × G. nelsonii | 9 | 6 | 66.7 | 41.0 ± 3.8 | 30.0–61.5 | 29.4 | 9.3 |
| 14. | G. herbaceum subsp. euherbaceum (cv. А-256) × G. australe | 36 | 1 | 2.7 | 50.0 | - | - | - |
| No. | Plant Samples | Vegetative Growth Duration, Days | |||||
|---|---|---|---|---|---|---|---|
| Range | S | V% | hp | h2 | |||
| Parental lines (G. herbaceum subspecies) | |||||||
| 1. | G. herbaceum subsp. africanum | 142.9 ± 0.4 | 139–146 | 1.9 | 1.38 | - | - |
| 2. | G. herbaceum subsp. pseudoarboreum | 121.3 ± 0.4 | 119–123 | 1.5 | 1.2 | - | - |
| 3. | G. herbaceum subsp. pseudoarboreum f. harga | 126.2 ± 0.4 | 123–128 | 1.9 | 1.5 | - | - |
| 4. | G. herbaceum subsp. frutescens | 117.2 ± 0.4 | 116–119 | 1.3 | 1.1 | - | - |
| 5. | G. herbaceum subsp. euherbaceum (cv. А-256) | 137.1 ± 0.4 | 135–139 | 1.6 | 1.2 | - | - |
| Parental lines (Australian wild species) | |||||||
| 6 | G. australe | 133.6 ± 0.3 | 132–135 | 1.1 | 0.8 | - | - |
| 7 | G. nelsonii | 135.3 ± 0.3 | 134–138 | 1.5 | 1.1 | - | - |
| 8 | G. bickii | 135.5 ± 0.4 | 133–138 | 1.8 | 1.4 | - | - |
| F1С polyploid hybrids | |||||||
| 9 | G. herbaceum subsp. frutescens × G. nelsonii | 127.5 ± 0.4 | 126–130 | 1.4 | 1.1 | –0.13 | - |
| 10 | G. herbaceum subsp. euherbaceum (cv. А-256) × G. nelsonii | 128.1 ± 0.5 | 125–130 | 1.9 | 1.5 | –0.9 | - |
| F2С polyploid hybrids | |||||||
| 11 | G. herbaceum subsp. frutescens × G. nelsonii | 116 ± 1.05 | 113–123 | 3.3 | 2.9 | 0.13 | –0.1 |
| 12 | G. herbaceum subsp. euherbaceum (cv. А-256) × G. nelsonii | 127.9 ± 1.7 | 123–137 | 5.3 | 4.2 | 9.2 | –0.1 |
| No. | Genotypes | Х ± Sх | Range | S | V (%) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Aphids | Whiteflies | Aphids | Whiteflies | Aphids | Whiteflies | Aphids | Whiteflies | ||
| 1 | ssp. frutescens | 7.3 ± 0.38 | 8.4 ± 0.22 | 5.5–8.7 | 7.4–9.2 | 1.07 | 0.62 | 14.63 | 7.47 |
| 2 | ssp. euherbaceum (cv. A-256) | 7.9 ± 0.46 | 8.9 ± 0.32 | 6.0–9.3 | 7.7–10.1 | 1.29 | 0.91 | 16.25 | 10.2 |
| 3 | G. nelsonii | 6.0 ± 0.37 | 6.3 ± 0.22 | 4.7–7.7 | 5.2–7.2 | 1.04 | 0.62 | 17.36 | 9.83 |
| 4 | ssp. euhirsutum (cv. Ravnak-1) | 22.1 ± 0.32 | 23.9 ± 0.43 | 21.0–23.3 | 22.5–25.5 | 0.91 | 1.23 | 4.08 | 5.14 |
| 5 | F1 (ssp. frutescens × G nelsonii) | 6.5 ± 0.28 | 6.4 ± 0.22 | 5.3–7.5 | 5.9–7.7 | 0.79 | 0.62 | 14.36 | 9.63 |
| 6 | F1 (ssp. euherbaceum × G. nelsonii) | 6.55 ± 0.42 | 6.8 ± 0.35 | 4.9–8.0 | 5.7–8.0 | 1.19 | 0.98 | 18.16 | 14.42 |
| 7 | F1C (ssp. frutescens × G.nelsonii) | 5.56 ± 0.33 | 5.4 ± 0.27 | 4.3–6.7 | 4.4–6.5 | 0.92 | 0.75 | 16.59 | 13.95 |
| 8 | F1C (ssp. euherbaceum × G. nelsonii) | 5.16 ± 0.19 | 5.8 ± 0.22 | 4.3–6.0 | 4.9–6.6 | 0.54 | 0.62 | 10.45 | 10.76 |
| 9 | F1 (ssp. frutescens × G.nelsonii) × cv.Ravnak-1 | 6.56 ± 0.30 | 7.5 ± 0.26 | 5.7–7.7 | 6.5–8.6 | 0.84 | 0.73 | 12.77 | 9.8 |
| 10 | F1 (ssp. euherbaceum × G. nelsonii) × cv. Ravnak-1 | 8.00 ± 0.30 | 10.6 ± 0.44 | 6.4–8.7 | 8.8–12.4 | 0.84 | 1.24 | 10.52 | 11.67 |
| No. | Plant Samples | Total Number of Spores | Meiotic Index, % | Micronuclear Tetrads, % | Polyads, % |
|---|---|---|---|---|---|
| Parental forms | |||||
| 1 | G. herbaceum subsp. frutescens | 787 | 94.2 ± 0.8 | - | 0.7 ± 0.3 |
| 2 | G. herbaceum subsp. euherbaceum (cv. А-256) | 907 | 89.8 ± 1.01 | 2.65 ± 0.5 | 2.7 ± 0.5 |
| 3 | G. nelsonii | 1003 | 95.6 ± 0.9 | - | 0.1 ± 0.02 |
| F1 hybrids | |||||
| 4 | G. herbaceum subsp. frutescens × G. nelsonii | 547 | 57.8 ± 2.1 | 20.3 ± 1.7 | 5.5 ± 0.9 |
| 5 | G. herbaceum subsp. euherbaceum (cv. А-256) × G. nelsonii | 1011 | 78.6 ± 1.3 | 6.9 ± 0.8 | 2.5 ± 05 |
| F1C allotetraploid hybrids | |||||
| 6 | G. herbaceum subsp. frutescens × G. nelsonii | 380 | 46.8 ± 2.6 | 37.6 ± 2.5 | 1.8 ± 0.7 |
| 7 | G. herbaceum subsp. euherbaceum (cv. А-256) × G. nelsonii | 378 | 47.1 ± 1.9 | 36.7 ± 2.5 | 1.9 ± 0.7 |
| No. | Hybrid Combinations | Total Number of Pollens | Pollen Fertility, % | |||
|---|---|---|---|---|---|---|
| Range | S | V % | ||||
| F1 hybrids | ||||||
| 1 | G. herbaceum subsp. frutescens × G. nelsonii | 1227 | 4.56 ± 0.35 | 0.75–7.3 | 0.60 | 0.4 |
| 2 | G.nelsonii × G. herbaceum subsp. frutescens | 1356 | 4.72 ± 0.33 | 0.75–7.54 | 0.58 | 0.3 |
| 3 | G. herbaceum subsp. pseudoarboreum × G. nelsonii | 879 | 6.14 ± 0.66 | 5.51–8.71 | 0.81 | 0.7 |
| 4 | G. nelsonii × G. herbaceum subsp. pseudoarboreum | 1112 | 5.67 ± 0.48 | 5.28–7.53 | 0.69 | 0.5 |
| 5 | G. herbaceum subsp. pseudoarboreum × G. australe | 1334 | 2.25 ± 0.17 | 1.96–2.87 | 0.41 | 0.2 |
| 6 | G. herbaceum subsp. pseudoarboreum f. harga × G. nelsonii | 512 | 8.79 ± 1.57 | 5.18–9.98 | 1.25 | 1.3 |
| 7 | G. nelsonii × G. herbaceum subsp. pseudoarboreum f. harga | 1660 | 7.29 ± 0.41 | 7.13–8.67 | 0.64 | 0.4 |
| 8 | G. herbaceum subsp. euherbaceum (cv. А-256) × G. australe | 685 | 4.82 ± 0.67 | 3.18–7.66 | 0.82 | 0.7 |
| 9 | G. herbaceum subsp. euherbaceum (cv. А-256) × G.nelsonii | 721 | 7.07 ± 0.91 | 6.98–11.1 | 0.95 | 0.9 |
| 10 | G. herbaceum subsp. euherbaceum (cv. А-256) × G. bickii | 825 | 4.73 ± 0.55 | 3.94–5.34 | 0.74 | 0.6 |
| F1C allotetraploid hybrids | ||||||
| 11 | G. herbaceum subsp. frutescens × G. nelsonii | 2307 | 47.2 ± 1.04 | 43.1–51.7 | 1.08 | 1.04 |
| 12 | G. herbaceum subsp. euherbaceum (cv. А-256) × G. nelsonii | 2306 | 54.1 ± 1.04 | 46.8–66.5 | 1.07 | 1.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arslanova, S.K.; Ernazarova, Z.A.; Ernazarova, D.K.; Turaev, O.S.; Safiullina, A.K.; Toshpulatov, A.K.; Kholova, M.D.; Azimova, L.A.; Rafiyeva, F.U.; Gapparov, B.M.; et al. Development and Characterization of Synthetic Allotetraploids Between Diploid Species Gossypium herbaceum and Gossypium nelsonii for Cotton Genetic Improvement. Plants 2025, 14, 1620. https://doi.org/10.3390/plants14111620
Arslanova SK, Ernazarova ZA, Ernazarova DK, Turaev OS, Safiullina AK, Toshpulatov AK, Kholova MD, Azimova LA, Rafiyeva FU, Gapparov BM, et al. Development and Characterization of Synthetic Allotetraploids Between Diploid Species Gossypium herbaceum and Gossypium nelsonii for Cotton Genetic Improvement. Plants. 2025; 14(11):1620. https://doi.org/10.3390/plants14111620
Chicago/Turabian StyleArslanova, Sevara K., Ziraatkhan A. Ernazarova, Dilrabo K. Ernazarova, Ozod S. Turaev, Asiya K. Safiullina, Abdulqahhor Kh. Toshpulatov, Madina D. Kholova, Laylo A. Azimova, Feruza U. Rafiyeva, Bunyod M. Gapparov, and et al. 2025. "Development and Characterization of Synthetic Allotetraploids Between Diploid Species Gossypium herbaceum and Gossypium nelsonii for Cotton Genetic Improvement" Plants 14, no. 11: 1620. https://doi.org/10.3390/plants14111620
APA StyleArslanova, S. K., Ernazarova, Z. A., Ernazarova, D. K., Turaev, O. S., Safiullina, A. K., Toshpulatov, A. K., Kholova, M. D., Azimova, L. A., Rafiyeva, F. U., Gapparov, B. M., Khalikov, K. K., Khidirov, M. T., Iskandarov, A. A., Kodirov, D. M., Turaev, O. Y., Maulyanov, S. A., Udall, J. A., Yu, J. Z., & Kushanov, F. N. (2025). Development and Characterization of Synthetic Allotetraploids Between Diploid Species Gossypium herbaceum and Gossypium nelsonii for Cotton Genetic Improvement. Plants, 14(11), 1620. https://doi.org/10.3390/plants14111620