

Comparative Morpho-Physiological, Biochemical, and Gene Expressional Analyses Uncover Mechanisms of Waterlogging Tolerance in Two Soybean Introgression Lines
 ,
,  ,
,
Abstract
1. Introduction
2. Results
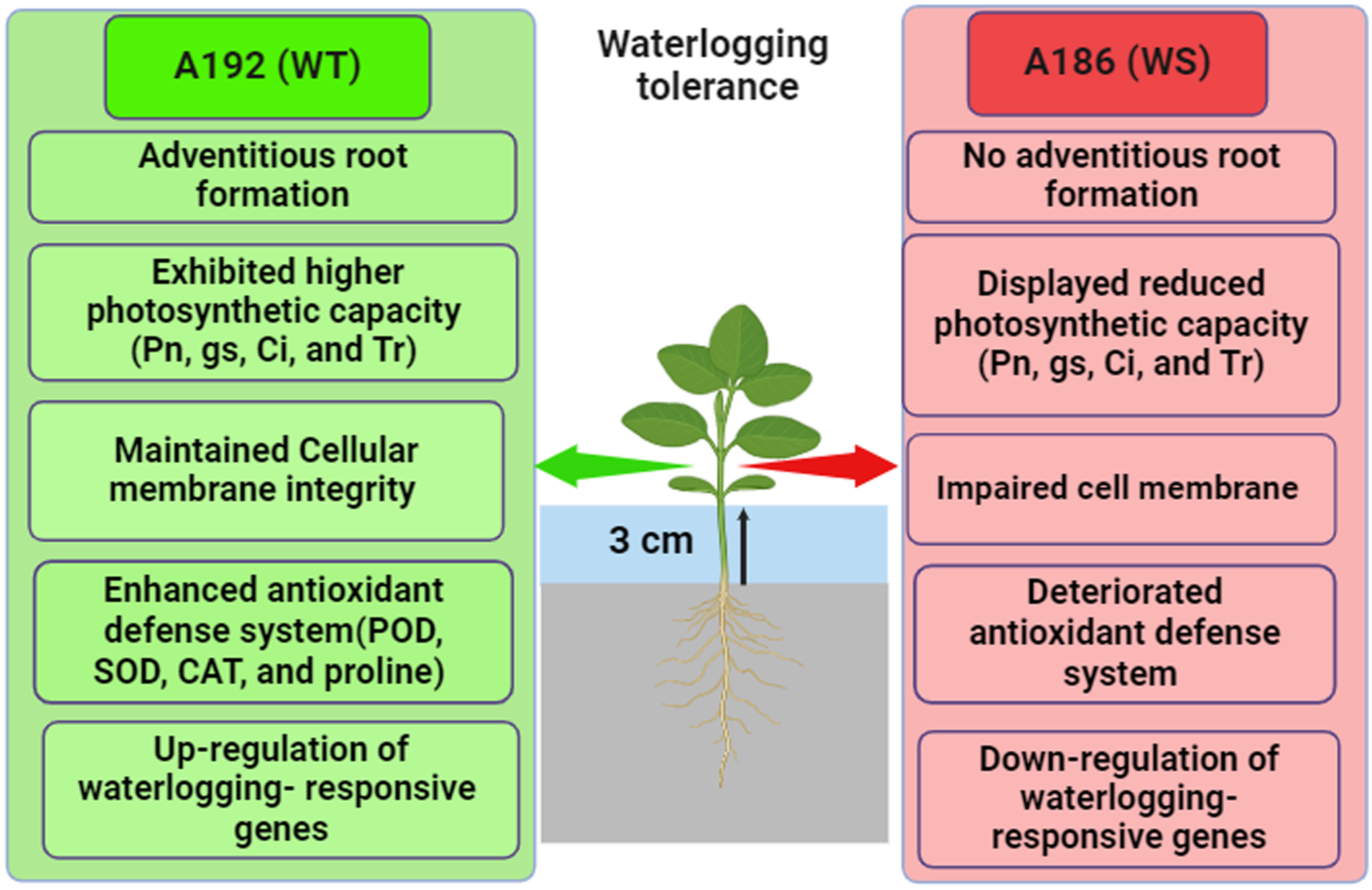
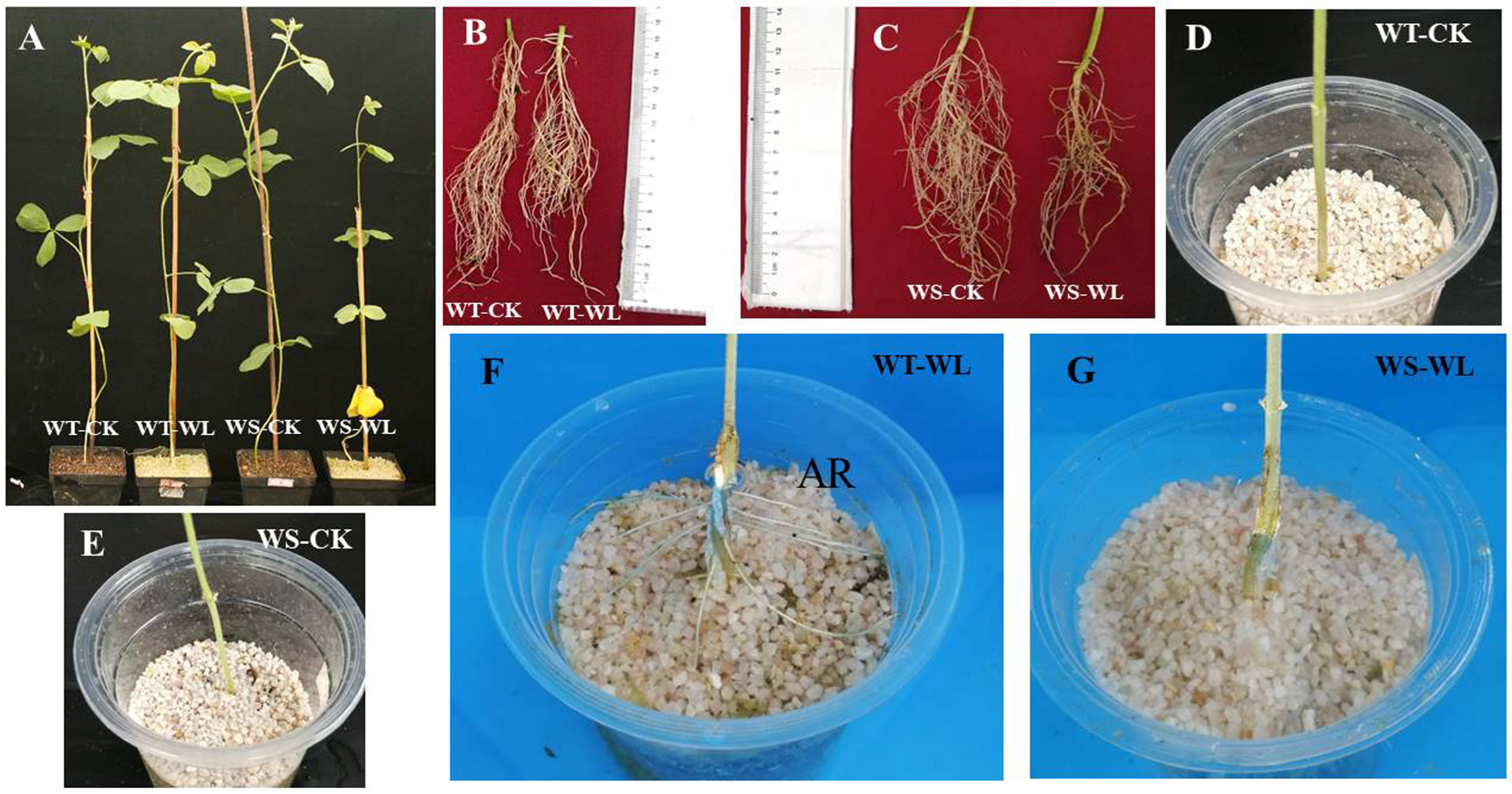
2.1. Morphological Differences between WT and WS Soybean Lines with and without Waterlogging
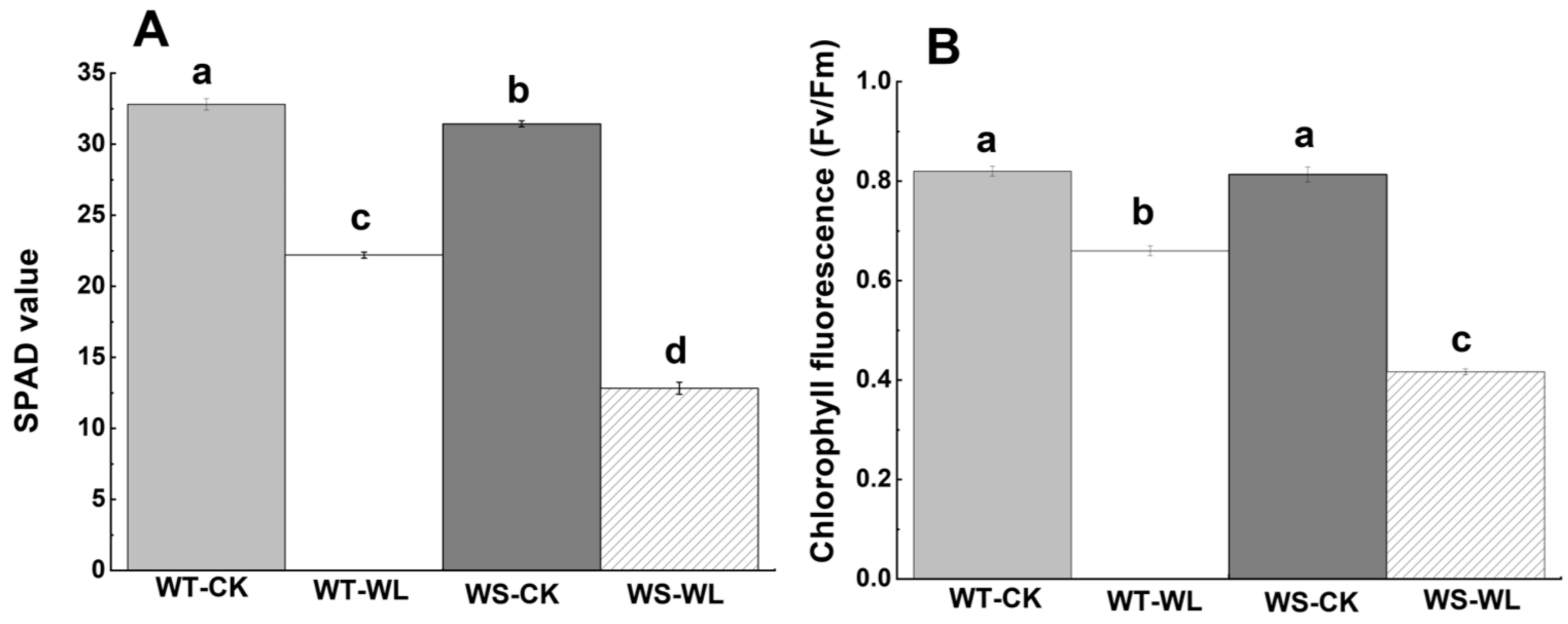
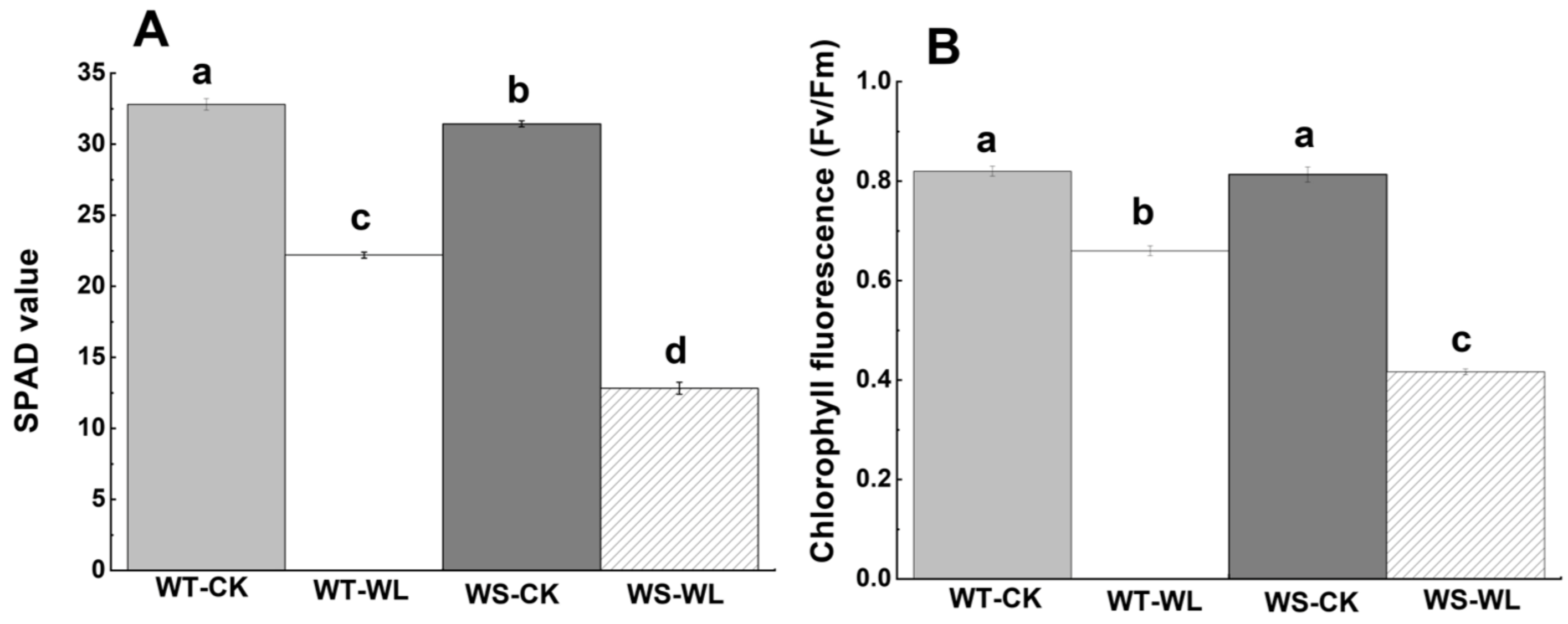
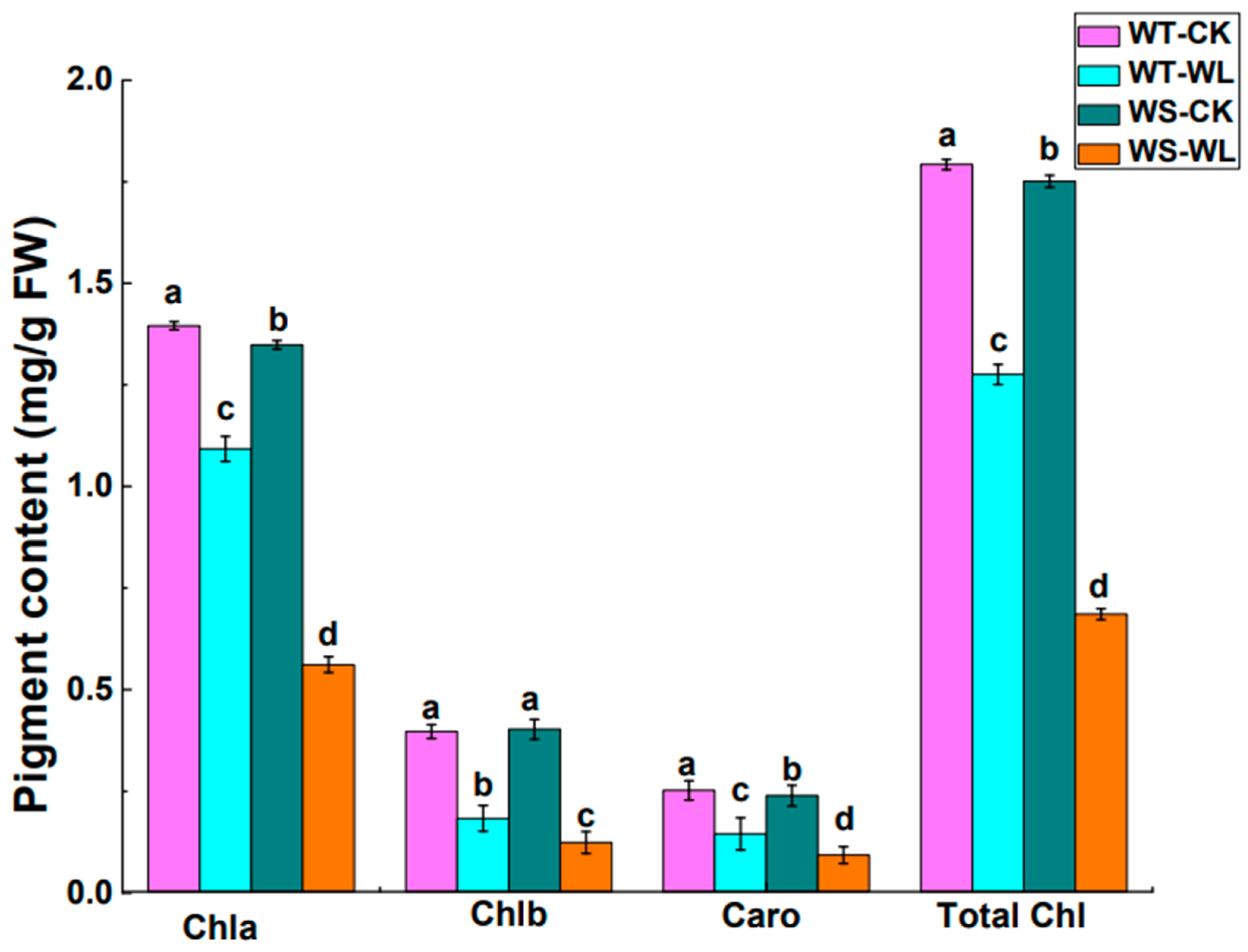
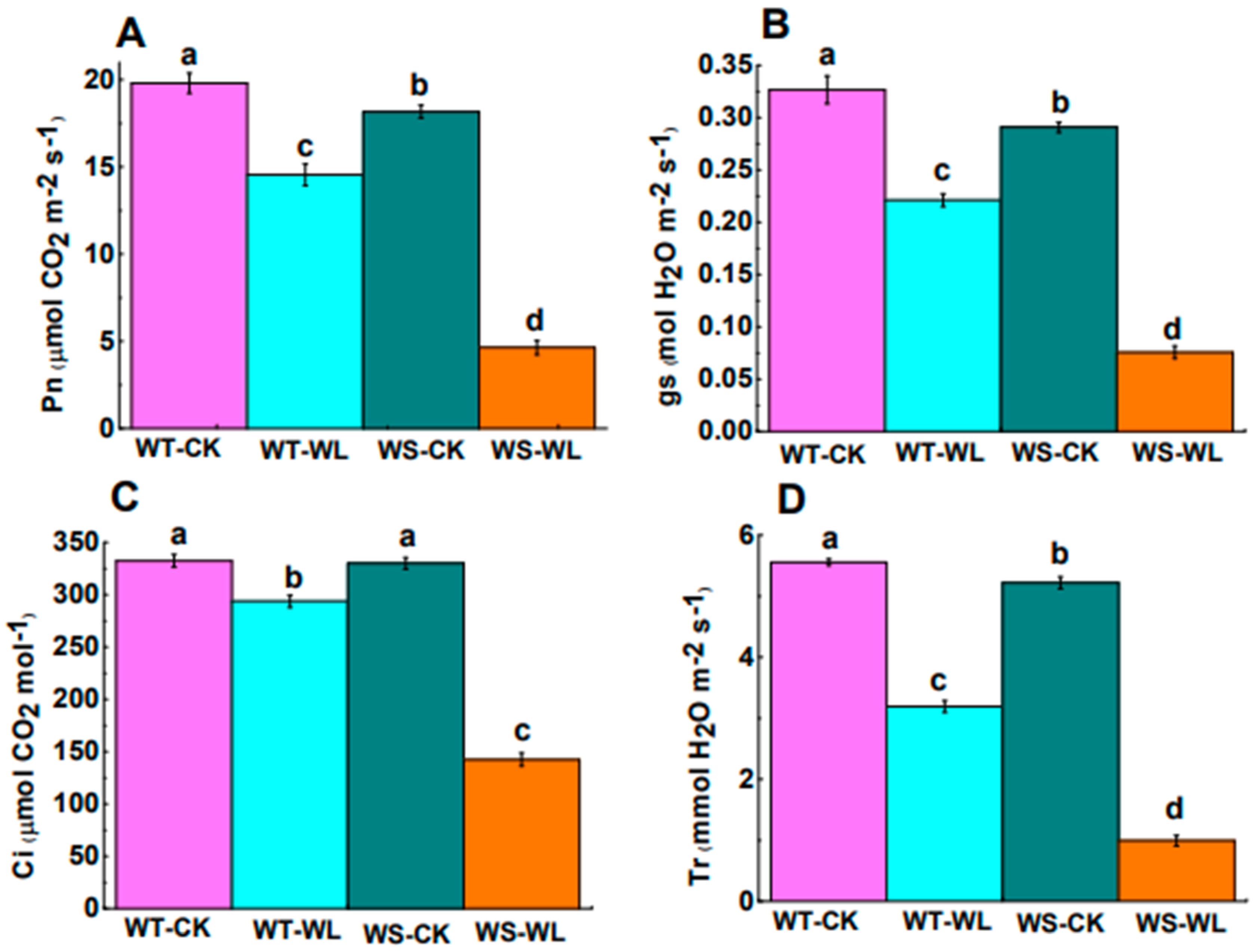
2.2. Physiological Differences between WT and WS Lines in Response to Waterlogged Conditions
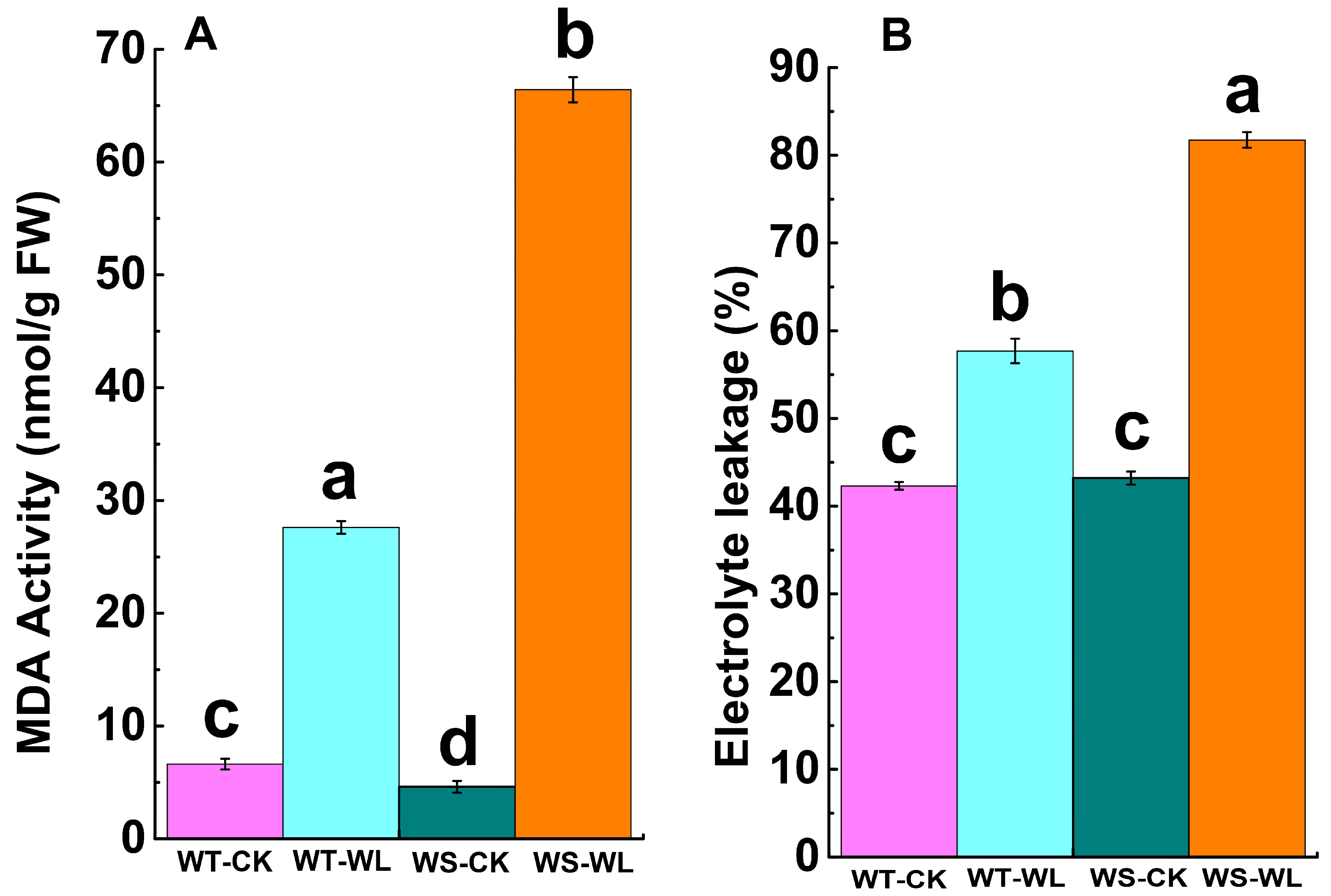
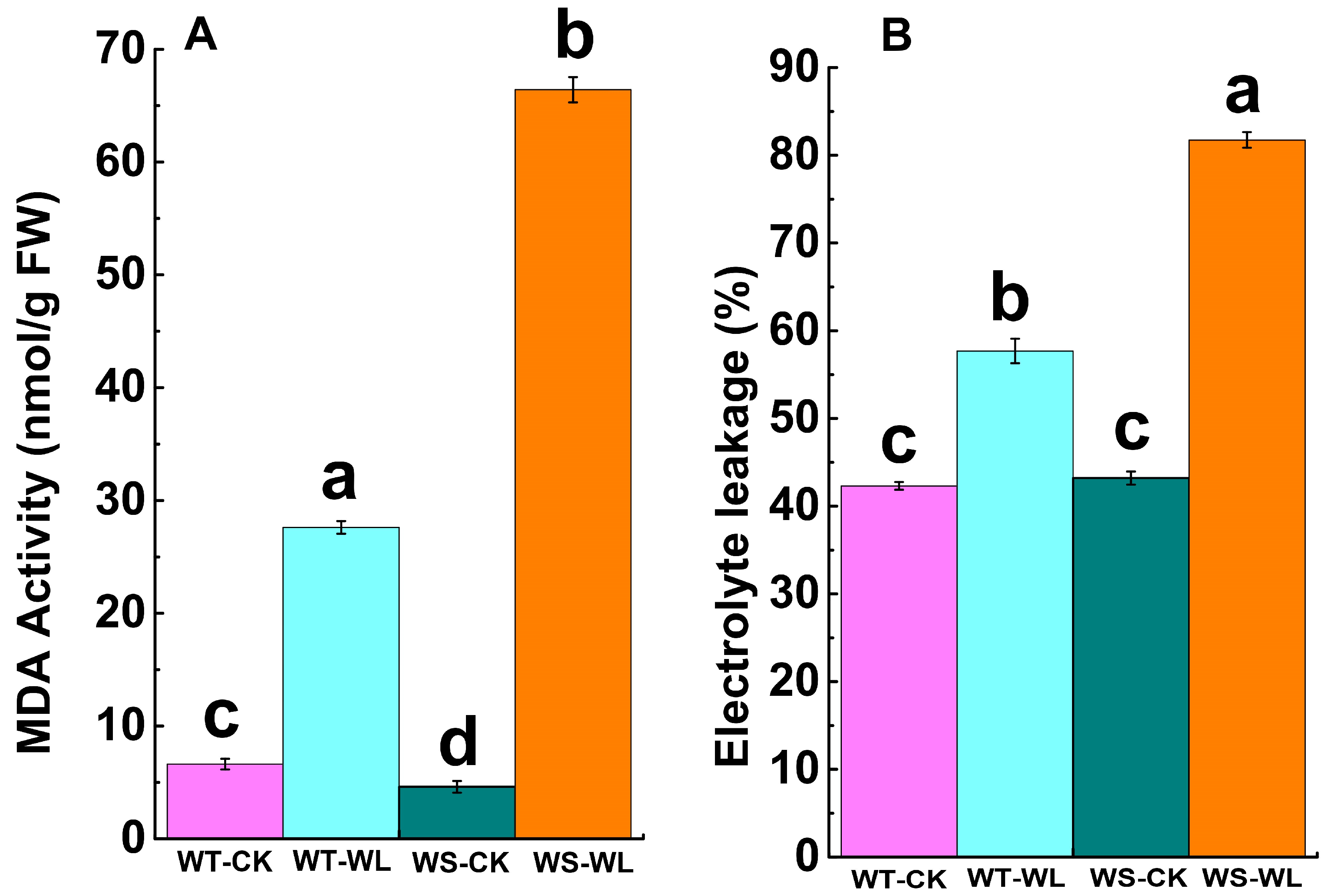
2.3. Cellular Membrane Integrity under Waterlogging Conditions
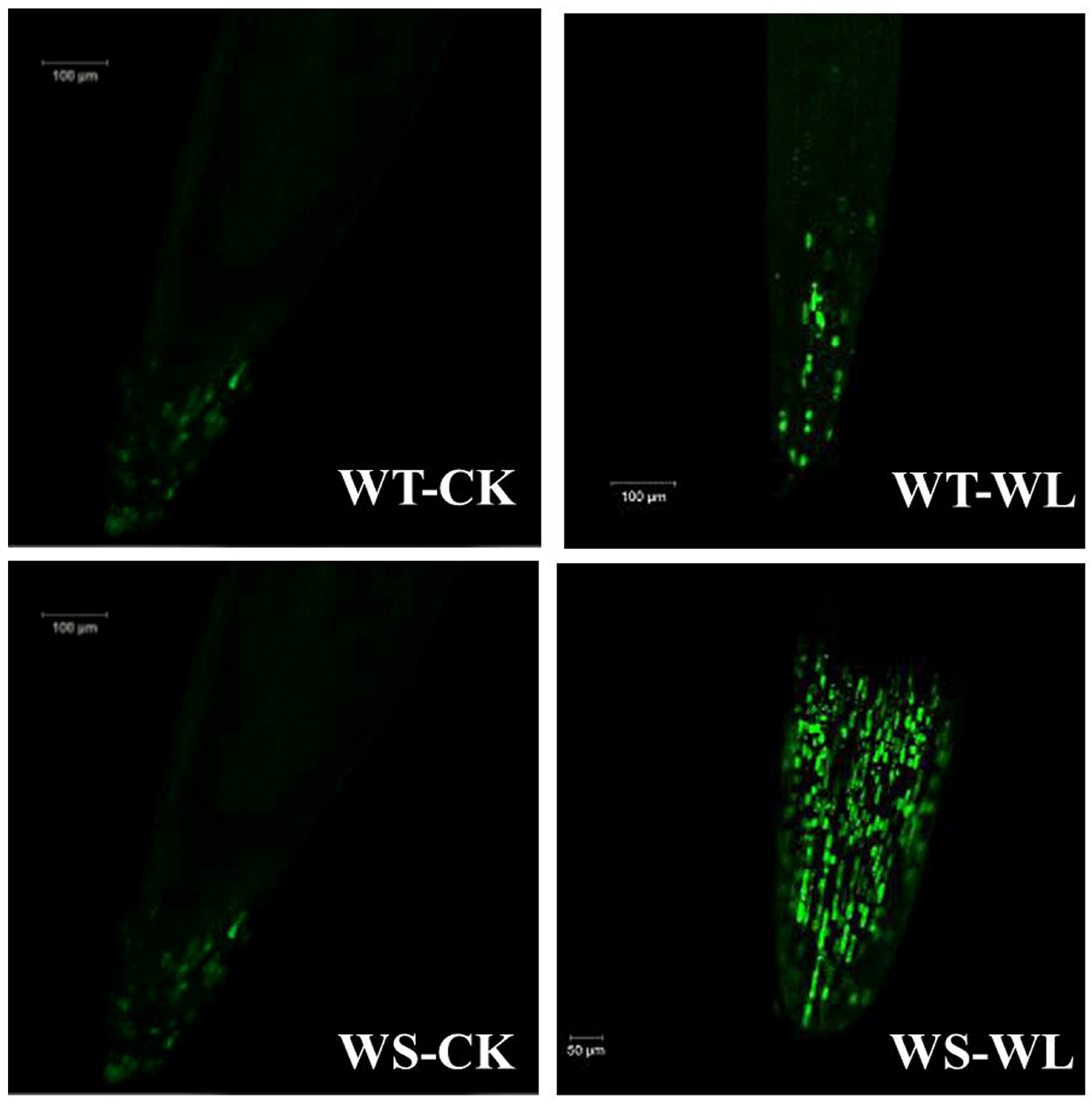
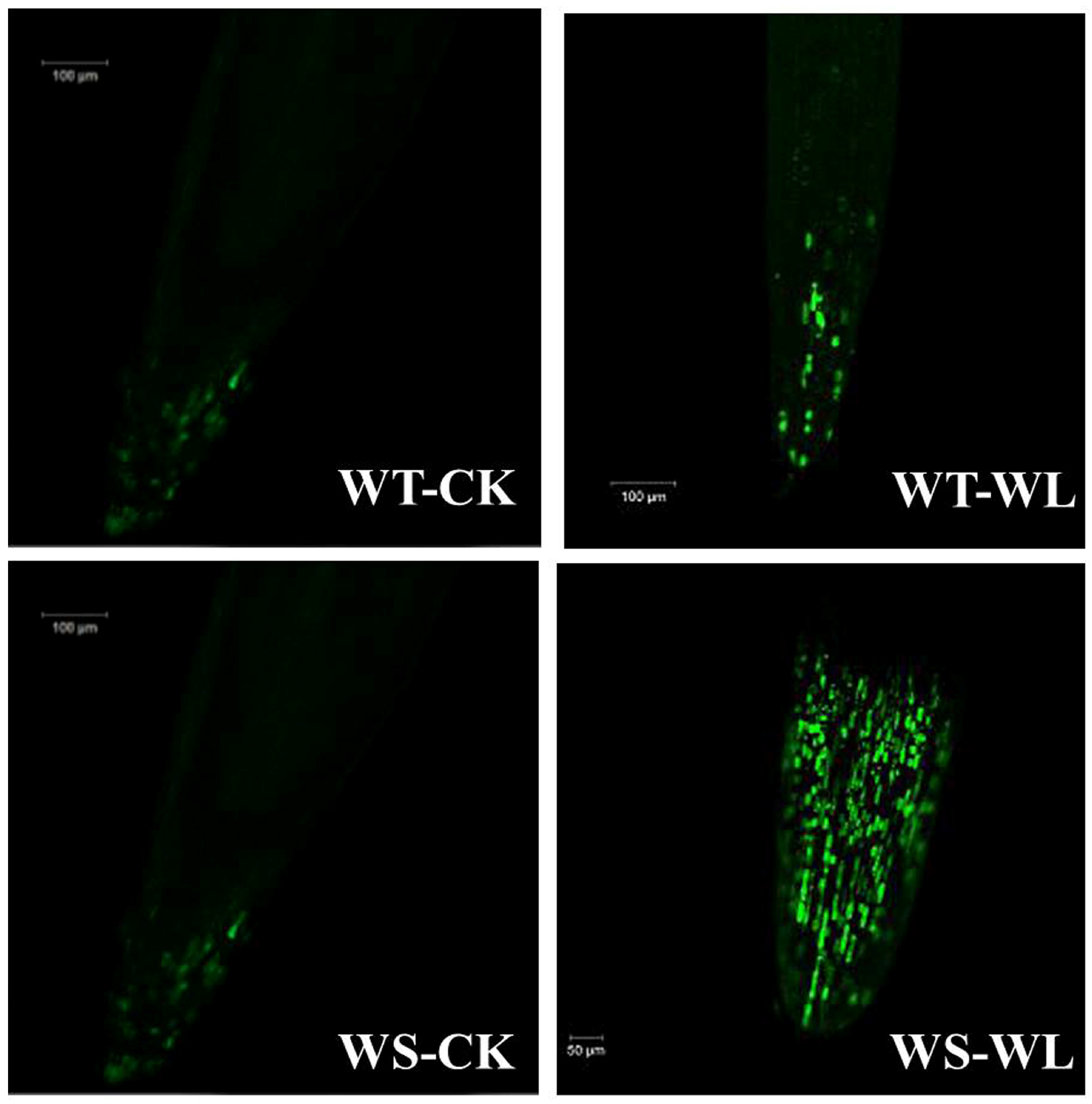
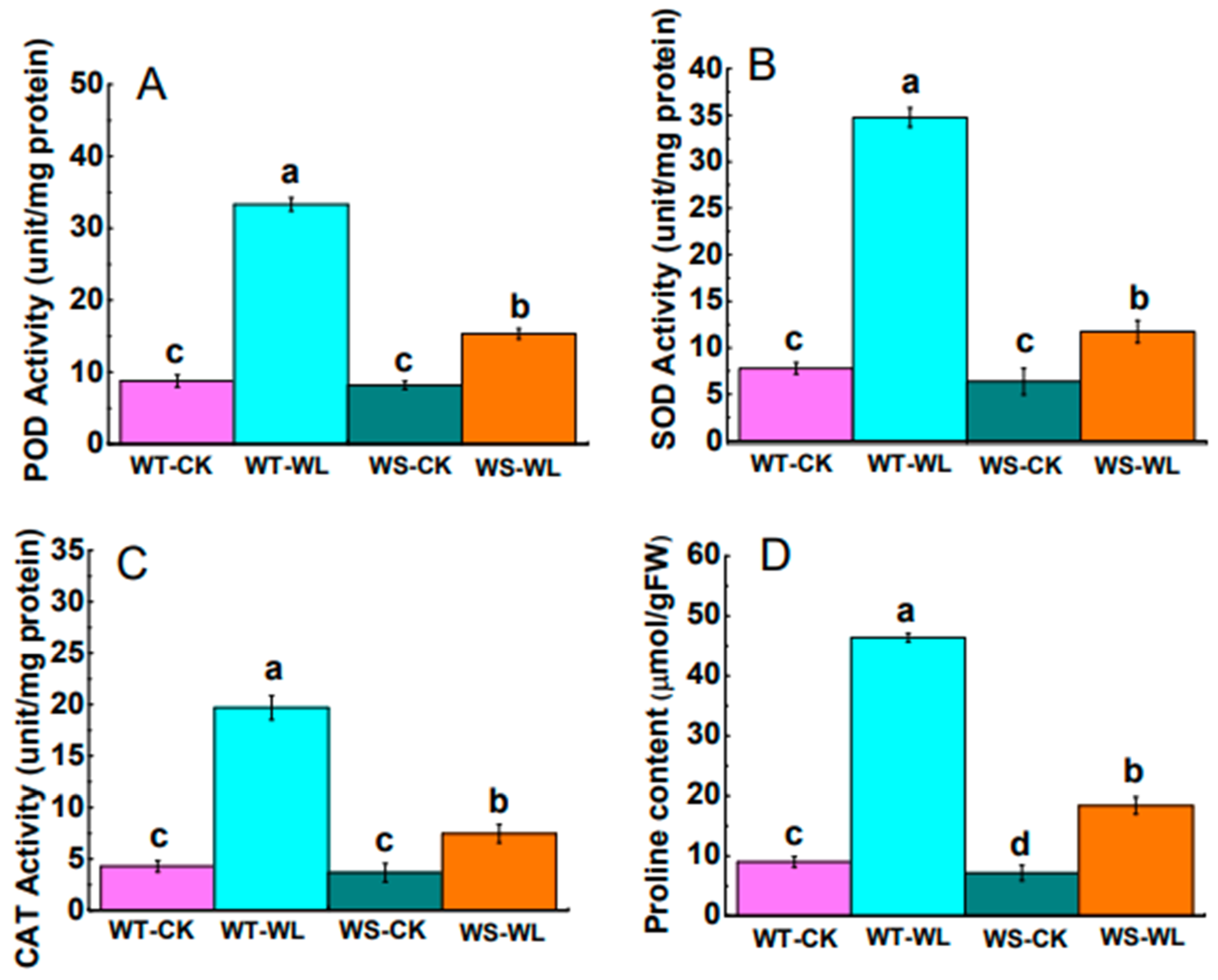
2.4. Changes in Antioxidant Defense System under Waterlogging Condition
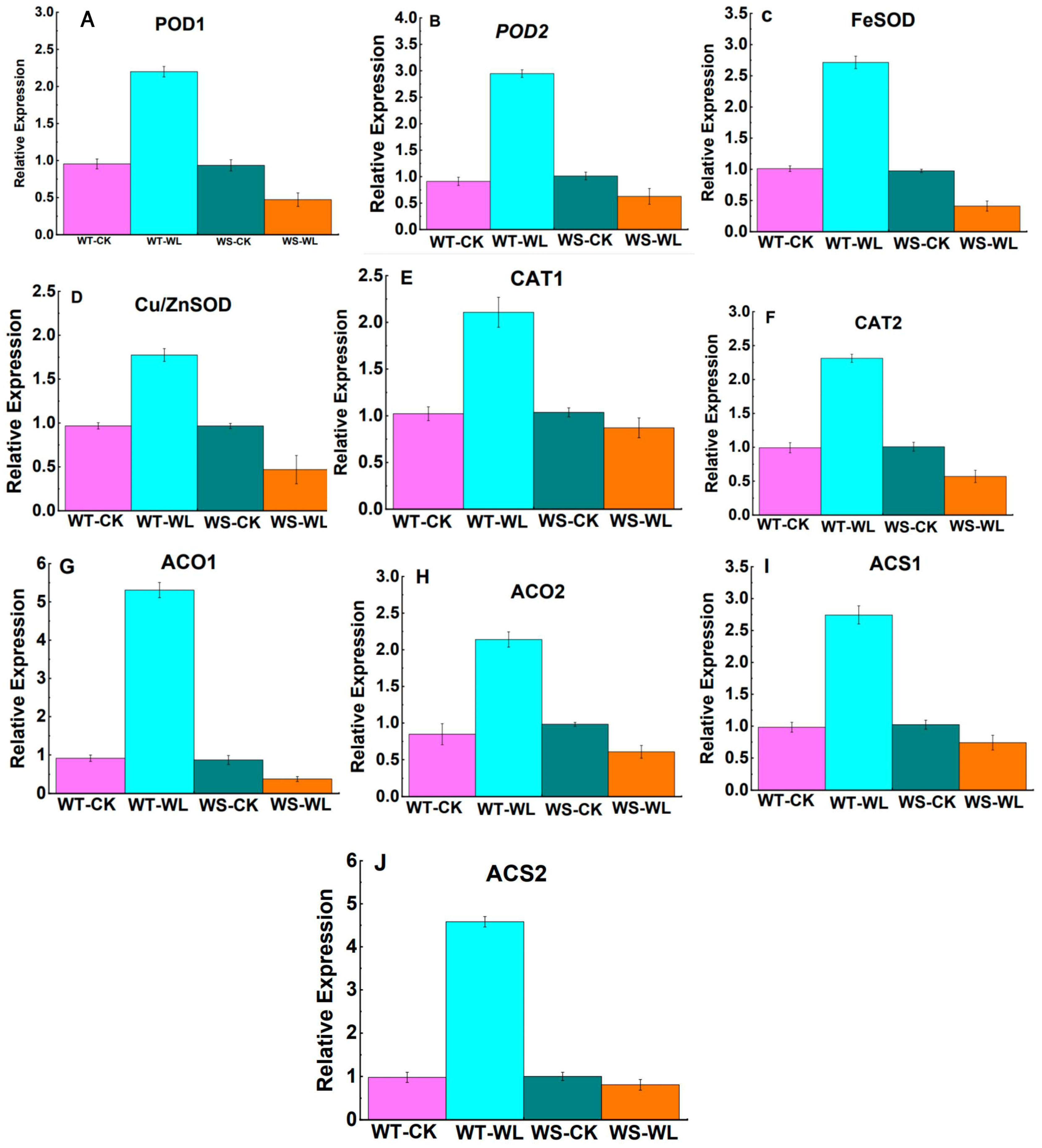
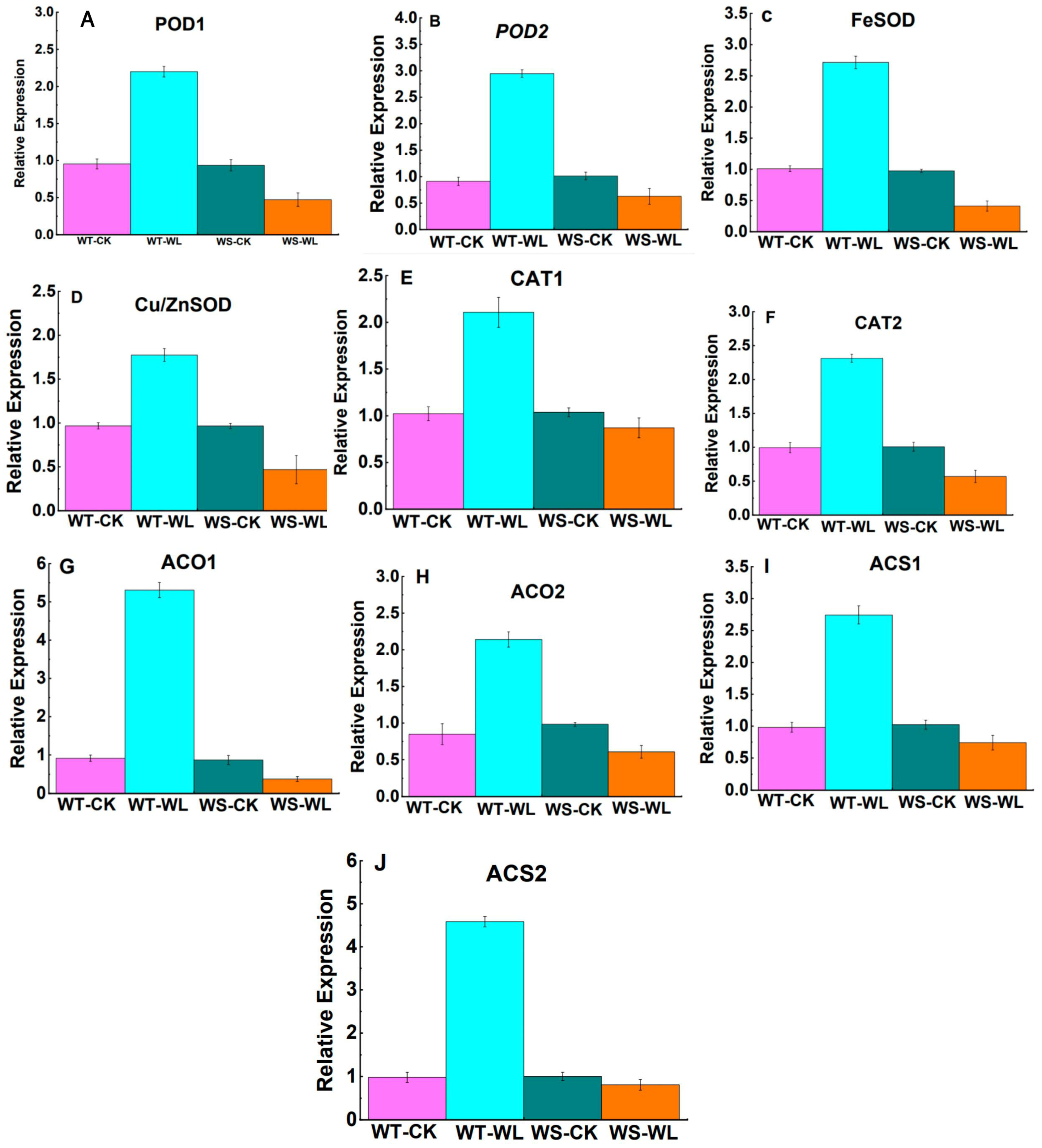
2.5. Gene Expression Analysis of Waterlogging-Responsive Genes
3. Discussion
3.1. WT Line Grows Better under Waterlogging Than WS Line
3.2. WT Line Had Better Photosynthetic Performance than WS Line under Waterlogging Stress
3.3. WT Line Maintained Cellular Membrane Integrity and Exhibited a Robust Antioxidant System under Waterlogging Stress
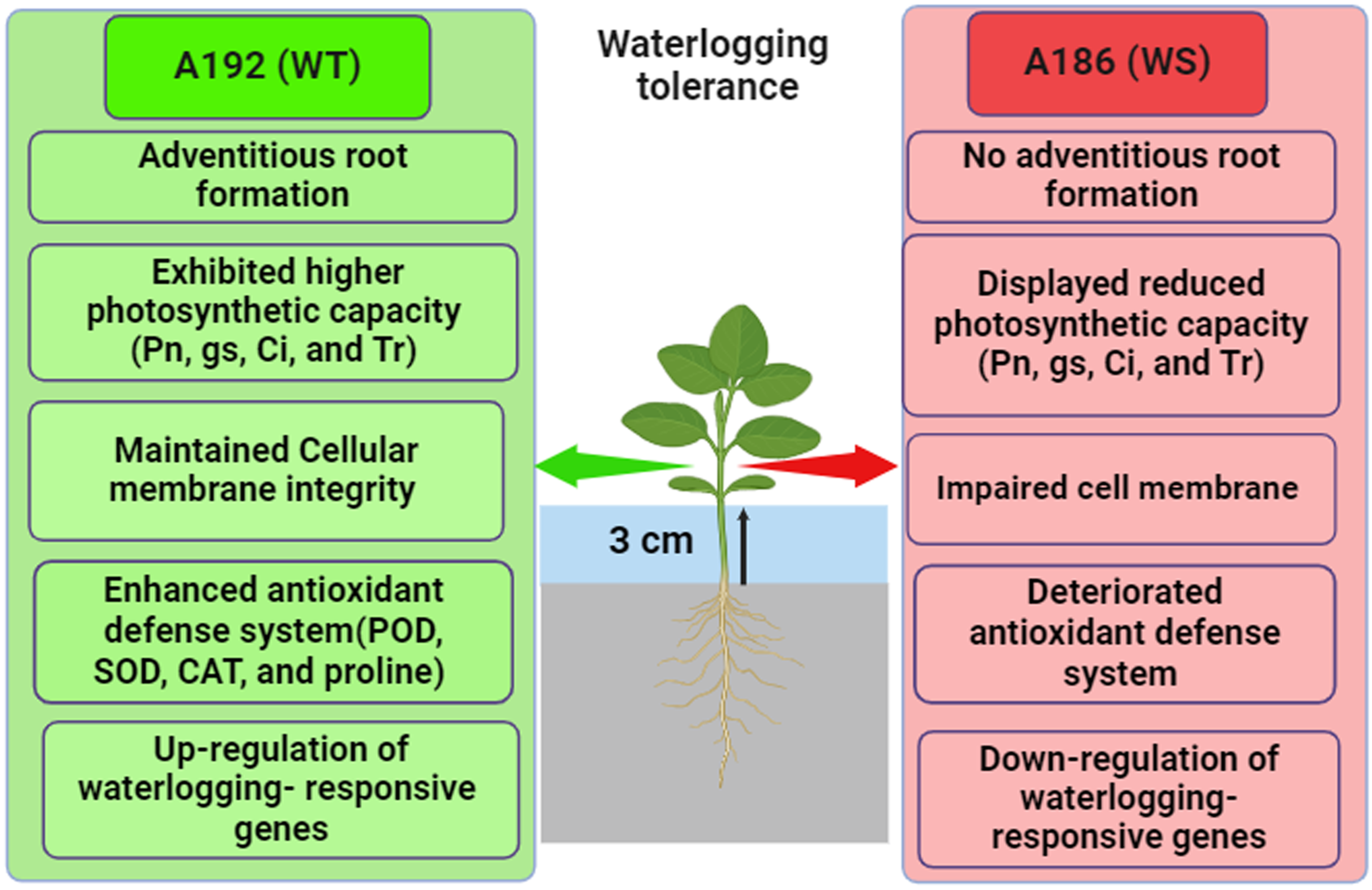
4. Conclusions
5. Materials and Methods
5.1. Plant Materials and Waterlogging Conditions
5.2. Measurement of Morpho-Physiological Parameters
5.3. Estimation of Antioxidant Enzyme Activity
5.4. H2O2 Detection
5.5. RNA Extraction and Gene Expression Analysis Using Quantitative Real-Time PCR
5.6. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Garcia, N.; Da-Silva, C.J.; Cocco, K.L.T.; Pomagualli, D.; de Oliveira, F.K.; da Silva, J.V.L.; de Oliveira, A.C.B.; Amarante, L.D. Waterlogging tolerance of five soybean genotypes through different physiological and biochemical mechanisms. Environ. Exp. Bot. 2020, 172, 103975. [Google Scholar] [CrossRef]
- Zhao, T.; Aleem, M.; Sharmin, R.A. Adaptation to water stress in soybean: Morphology to genetics. In Plant, Abiotic Stress Responses Climate Change; Andjelkovic, V., Ed.; Intech Open: Shanghai, China, 2018; pp. 34–68. [Google Scholar]
- Linkemer, G.; Board, J.E.; Musgrave, M.E. Water logging effects on growth and yield components in late planted soybean. Crop Sci. 1998, 38, 1576–1584. [Google Scholar] [CrossRef]
- Ploschuk, R.A.; Miralles, D.J.; Striker, G.G. A quantitative review of soybean responses to waterlogging: Agronomical, morphophysiological and anatomical traits of tolerance. Plant Soil 2022, 475, 237–252. [Google Scholar] [CrossRef]
- Adegoye, G.A.; Olorunwa, O.J.; Alsajri, F.A.; Walne, C.H.; Wijewandana, C.; Kethireddy, S.R.; Reddy, K.N.; Reddy, K.R. Waterlogging Effects on Soybean Physiology and Hyperspectral Reflectance during the Reproductive Stage. Agriculture 2023, 13, 844. [Google Scholar] [CrossRef]
- Nguyen, V.; Vuong, T.; VanToai, T.; Lee, J.; Wu, X.; Mian, M.; Dorrance, A.; Shannon, J.; Nguyen, H. Mapping of quantitative trait loci associated with resistance to and flooding tolerance in soybean. Crop Sci. 2012, 52, 2481–2493. [Google Scholar] [CrossRef]
- Yang, L.; Li, N.; Liu, Y.; Miao, P.; Liu, J.; Wang, Z. Updates and Prospects: Morphological, physiological, and molecular Regulation in crop Response to waterlogging stress. Agronomy 2023, 13, 2599. [Google Scholar] [CrossRef]
- Sathi, K.S.; Masud, A.A.; Anee, T.I.; Rahman, K.; Ahmed, N.; Hasanuzzaman, M. Soybean plants under waterlogging stress: Responses and adaptation mechanisms. In Managing Plant Production under Changing Environment; Hasanuzzaman, M., Ahammed, G.J., Nahar, K., Eds.; Springer Nature: Singapore, 2022; pp. 103–134. [Google Scholar]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Da-Silva, C.J.; do Amarante, L. Time-course biochemical analyses of soybean plants during waterlogging and reoxygenation. Environ. Exp. Bot. 2020, 1, 104242. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Gong, X.; Xu, Y.; Li, H.; Chen, X.; Song, Z. Antioxidant activation, cell wall reinforcement, and reactive oxygen species regulation promote resistance to waterlogging stress in hot pepper (Capsicum annuum L.). BMC Plant Biol. 2022, 22, 425. [Google Scholar] [CrossRef]
- Jia, W.; Ma, M.; Chen, J.; Wu, S. Plant Morphological, Physiological and Anatomical Adaption to Flooding Stress and the Underlying Molecular Mechanisms. Int. J. Mol. Sci. 2021, 22, 1088. [Google Scholar] [CrossRef] [PubMed]
- Sathi, K.S.; Masud, A.A.; Falguni, M.R.; Ahmed, N.; Rahman, K.; Hasanuzzaman, M. Screening of soybean genotypes for waterlogging stress tolerance and understanding the physiological mechanisms. Adv. Agric. 2022, 2022, 5544665. [Google Scholar] [CrossRef]
- Sairam, R.K.; Kumutha, D.; Ezhilmathi, K.; Chinnusamy, V.; Meena, R.C. Waterlogging induced oxidative stress and antioxidant enzyme activities in pigeon pea. Biol. Plant 2009, 53, 493–504. [Google Scholar] [CrossRef]
- Teoh, E.Y.; Teo, C.H.; Baharum, N.A.; Pua, T.-L.; Tan, B.C. Waterlogging Stress Induces Antioxidant Defense Responses, Aerenchyma Formation and Alters Metabolisms of Banana Plants. Plants 2022, 11, 2052. [Google Scholar] [CrossRef]
- Lee, Y.H.; Kim, K.S.; Jang, Y.S.; Hwang, J.H.; Lee, D.H.; Choi, I.H. Global gene expression responses to waterlogging in leaves of rape seedlings. Plant Cell Rep. 2014, 33, 289–299. [Google Scholar] [CrossRef]
- Phule, A.S.; Barbadikar, K.M.; Madhav, M.S.; Subrahmanyam, D.; Senguttuvel, P.; Babu, M.B.B.P.; Kumar, P.A. Studies on root anatomy, morphology and physiology of rice grown under aerobic and anaerobic conditions. Physiol. Mol. Biol. Plants 2019, 25, 197–205. [Google Scholar] [CrossRef]
- Malik, A.I.; Colmer, T.D.; Lambers, H.; Setter, T.L.; Schortemeyer, M. Short-term waterlogging has long-term effects on the growth and physiology of wheat. New Phytol. 2002, 153, 225–236. [Google Scholar] [CrossRef]
- Liang, K.; Tang, K.; Fang, T.; Qiu, F. Waterlogging tolerance in maize: Genetic and molecular basis. Mol. Breed. 2020, 40, 111. [Google Scholar] [CrossRef]
- Lin, H.H.; Lin, K.H.; Syu, J.Y.; Tang, S.Y.; Lo, H.F. Physiological and proteomic analysis in two wild tomato lines under waterlogging and high temperature stress. J. Plant Biochem. Biotechnol. 2016, 25, 87–96. [Google Scholar] [CrossRef]
- Chandra, S.; Das, R.T.; Nagar, S.; Satpute, G.K.; Kumawat, G.; Ratnaparkhe, M.B.; Gupta, S.; Rajesh, V.; Nataraj, V.; Shivakumar, M.; et al. Soybean improvement for waterlogging tolerance. In Soybean Improvement: Physiological, Molecular and Genetic Perspectives; Wani, S.H., Sofi, N.U.R., Bhat, M.A., Lin, F., Eds.; Springer: Cham, Switzerland, 2022; pp. 73–97. [Google Scholar]
- Bacanamwo, M.; Purcell, L.C. Soybean dry matter and N accumulation responses to flooding stress, N sources and hypoxia. J. Exp. Bot. 1999, 50, 689–696. [Google Scholar] [CrossRef]
- Colmer, T.D.; Voesenek, L.A. Flooding tolerance: Suites of plant traits in variable environments. Funct. Plant Biol. 2009, 36, 665–681. [Google Scholar] [CrossRef] [PubMed]
- De Klerk, G.-J.; van der Krieken, W.; de Jong, J.C. Review the formation of adventitious roots: New concepts, new possibilities. In Vitro Cell. Dev. Biol. Plant 1999, 35, 189–199. [Google Scholar] [CrossRef]
- Nguyen, T.N.; Tuan, P.A.; Mukherjee, S.; Son, S.; Ayele, B.T. Hormonal regulation in adventitious roots and during their emergence under waterlogged conditions in wheat. J. Exp. Bot. 2018, 69, 4065–4082. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Li, Q.; Ma, X.; Qian, C.; Wang, H.; Ren, N.; Shen, C.; Huang, S.; Xu, X.; Xu, Q.; et al. Waterlogging-induced adventitious root formation in cucumber is regulated by ethylene and auxin through reactive oxygen species signalling. Plant Cell Environ. 2019, 42, 1458–1470. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- Ashraf, M.H.; Harris, P.J. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.S. Response of plants to water stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Li, D.; Wang, L.; Ding, X.; Zhang, Y.; Gao, Y.; Zhang, X. Morpho-anatomical and physiological responses to waterlogging of sesame (Sesamum indicum L.). Plant Sci. 2013, 208, 102–111. [Google Scholar] [CrossRef]
- Anee, T.I.; Nahar, K.; Rahman, A.; Mahmud, J.A.; Bhuiyan, T.F.; Alam, M.U.; Fujita, M.; Hasanuzzaman, M. Oxidative damage and antioxidant defense in Sesamum indicum after different waterlogging durations. Plants 2019, 8, 196. [Google Scholar] [CrossRef]
- Alkahtani, M.D.; Attia, K.A.; Hafez, Y.M.; Khan, N.; Eid, A.M.; Ali, M.A.; Abdelaal, K.A. Chlorophyll fluorescence parameters and antioxidant defense system can display salt tolerance of salt acclimated sweet pepper plants treated with chitosan and plant growth promoting rhizobacteria. Agronomy 2020, 10, 1180. [Google Scholar] [CrossRef]
- Shah, A.; Gadol, N.; Priya, G.; Mishra, P.; Rao, M.; Singh, N.K.; Kumar, R.; Kalia, S.; Rai, V. Morpho-physiological and metabolites alteration in the susceptible and tolerant genotypes of sesame under waterlogging stress and post-waterlogging recovery. Plant Stress 2024, 11, 100361. [Google Scholar] [CrossRef]
- Ahmed, S.; Nawata, E.; Hosokawa, M.; Domae, Y.; Sakuratani, T. Alterations in photosynthesis and some antioxidant enzymatic activities of mungbean subjected to waterlogging. Plant Sci. 2002, 163, 117–123. [Google Scholar] [CrossRef]
- Sarkar, R.K.; Avijit, D. Changes in anti-oxidative enzymes and antioxidants in relation to flooding tolerance in rice. J. Plant Biol. 2000, 27, 307–311. [Google Scholar]
- Yin, X.; Zhang, C.; Song, X.; Jiang, Y. Interactive short-term effects of waterlogging and salinity stress on growth and carbohydrate, lipid peroxidation, and nutrients in two perennial ryegrass cultivars. J. Am. Soc. Hortic. 2017, 142, 110–118. [Google Scholar] [CrossRef]
- Kumutha, D.; Ezhilmathi, K.; Sairam, R.K.; Srivastava, G.C.; Deshmukh, P.S.; Meena, R.C. Waterlogging induced oxidative stress and antioxidant activity in pigeonpea genotypes. Biol. Plant 2009, 53, 75–84. [Google Scholar] [CrossRef]
- Zhang, G.; Tanakamaru, K.; Abe, J.; Morita, S. Influence of waterlogging on some anti-oxidative enzymatic activities of two barley genotypes differing in anoxia tolerance. Acta Physiol. Plant 2007, 29, 171–176. [Google Scholar] [CrossRef]
- Bin, T.; Xu, S.Z.; Zou, X.L.; Zheng, Y.L.; Qiu, F.Z. Changes of antioxidative enzymes and lipid peroxidation in leaves and roots of waterlogging-tolerant and waterlogging-sensitive maize genotypes at seedling stage. Agric. Sci. China 2010, 9, 651–661. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Bansal, R.; Srivastava, J.P. Antioxidative defense system in pigeon pea roots under waterlogging stress. Acta Physiol. Plant 2012, 34, 515–522. [Google Scholar] [CrossRef]
- Sairam, R.K.; Dharmar, K.; Lekshmy, S.; Chinnusamy, V. Expression of antioxidant defense genes in mung bean (Vigna radiata L.) roots under water-logging is associated with hypoxia tolerance. Acta Physiol. Plant 2011, 3, 735–744. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed]
- Kavi Kishor, P.B.; Sreenivasulu, N.E.S.E. Is proline accumulation per se correlated with stress tolerance or is proline homeostasis a more critical issue? Plant Cell Environ. 2014, 37, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Yordanova, R.Y.; Christov, K.N.; Popova, L.P. Antioxidative enzymes in barley plants subjected to soil flooding. Environ. Exp. Bot. 2004, 51, 93–101. [Google Scholar] [CrossRef]
- Horchani, F.; Stammitti-Bert, L.; Baldet, P.; Brouquisse, R.; Rolin, D.; Aschi-Smiti, S.; Raymond, P.; Gallusci, P. Effect of prolonged root hypoxia on the antioxidant content of tomato fruit. Plant Sci. 2010, 179, 209–218. [Google Scholar] [CrossRef]
- Barickman, T.C.; Simpson, C.R.; Sams, C.E. Waterlogging causes early modification in the physiological performance, carotenoids, chlorophylls, proline, and soluble sugars of cucumber plants. Plants 2019, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Tamang, B.G.; Li, S.; Rajasundaram, D.; Lamichhane, S.; Fukao, T. Overlapping and stress-specific transcriptomic and hormonal responses to flooding and drought in soybean. Plant J. 2021, 107, 100–117. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Duan, Y.; Yang, J.; Gan, L.; Chen, W.; Yang, J.; Xiao, G.; Guan, L.; Chen, J. Transcriptome Analysis Reveals Genes Associated with Flooding Tolerance in Mulberry Plants. Life 2023, 13, 1087. [Google Scholar] [CrossRef] [PubMed]
- Sharmin, R.A.; Bhuiyan, M.R.; Lv, W.; Yu, Z.; Chang, F.; Kong, J.; Bhat, J.A.; Zhao, T. RNA-Seq based transcriptomic analysis revealed genes associated with seed-flooding tolerance in wild soybean (Glycine soja Sieb. & Zucc.). Environ. Exp. Bot. 2020, 171, 103906. [Google Scholar]
- Kim, K.H.; Cho, M.J.; Kim, J.-M.; Lee, T.; Heo, J.H.; Jeong, J.Y.; Lee, J.; Moon, J.K.; Kang, S. Growth response and developing simple test method for waterlogging stress tolerance in soybean. J. Crop Sci. Biotechnol. 2019, 22, 371–378. [Google Scholar] [CrossRef]
- Fehr, W.R.; Caviness, C.E.; Burmood, D.T.; Pennington, J.S. Stage of development descriptions for soybeans, Glycine max (L.) Merrill. Crop Sci. 1971, 11, 929–931. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids-pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Kaur, N.; Sharma, I.; Kirat, K.; Pati, P.K. Detection of reactive oxygen species in Oryza sativa L. (rice). Bio-Protoc. 2016, 6, e2061. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Point | Line | Treatment | Root Length (cm) | Shoot Length (cm) | Root Fresh Weight (g) | Shoot Fresh Weight (g) | Root Dry Weight (g) | Shoot Dry Weight (g) |
|---|---|---|---|---|---|---|---|---|
| 21 DAT | WT | Control | 30.40 ± 0.19 a | 75.20 ± 0.06 a | 6.51 ± 0.16 a | 13.43 ±.05 a | 1.16 ± 0.08 a | 3.68 ± 0.07 a |
| Waterlogged | 27.18 ± 0.02 b | 70.17 ± 0.67 b | 4.88 ± 0.04 b | 11.23 ± 0.06 b | 0.88 ± 0.07 b | 2.55 ± 0.06 b | ||
| WS | Control | 30.49 ± 0.08 a | 73.48 ± 0.38 a | 6.37 ± 0.36 a | 13.40 ± 0.03 a | 1.14 ± 0.05 a | 3.56 ± 0.05 a | |
| Waterlogged | 20.13 ± 0.10 c | 53.77 ± 0.82 c | 2.38 ± 0.14 c | 8.47 ± 0.03 c | 0.41 ± 0.02 c | 1.65 ± 0.02 c |
| Gene ID | Gene Name | Gene Function | Primer Sequence (5′->3′) | |
|---|---|---|---|---|
| Glyma.02G008900 | POD1 | Peroxidase 5-like | F: | TGACACTGTCTGGAGCACAT |
| R: | AAAGAGTACAAGCGGTCGGA | |||
| Glyma.17G053000 | POD2 | Peroxidase N-like | F: | TATTGGCCGAGCAAGGTGTA |
| R: | ACTTTGCAGGTCAGAGAGCA | |||
| Glyma.02G087700 | FeSOD | Superoxide dismutase (Fe) | F: | TGGTGAAGACTCCCAATGCT |
| R: | TGACTGCATCCCAAGACACA | |||
| Glyma.11G192700 | Cu/ZnSOD | Superoxide dismutase (Cu-Zn) | F: | CCCTTTCTCCGGTCATCCTT |
| R: | CTTGGAGCGTGAAAGCGTTA | |||
| Glyma.04G017500 | CAT1 | Catalase | F: | TGGTCGCTTGGTCCTGAATA |
| R: | GGTCCAAGTCTGTGCCTTTG | |||
| Glyma.17G261700 | CAT2 | Catalase | F: | ACAAGAATCGGCCATCAAGC |
| R: | GCAAGCTTCTCCACCAGATG | |||
| Glyma.04G245900 | ACO1 | ACC oxidase 5 | F: | GCGTCATCCTACTCCTCCAA |
| R: | ACCATTGCTCAGGACCTCAA | |||
| Glyma.09G255000 | ACO2 | ACC oxidase homolog 1 | F: | CCTAAGGCCTCCATTTGCAC |
| R: | TCTCACTCTCCCACCTCTCA | |||
| Glyma.05G223000 | ACS1 | ACC synthase 7 | F: | AACAGCAATGGCAAGCTTCA |
| R: | AGAGCATCTCCTGGGTTAGC | |||
| Glyma.08G030100 | ACS2 | ACC synthase 7 | F: | AACAGCAATGGCAAGCTTCA |
| R: | GGCGTTGGAACAAGTAGAGC | |||
| Glyma.18G290800 | Actin11 | Reference gene | F: | CGGTGGTTCTATCTTGGCATC |
| R: | GTCTTTCGCTTCAATAACCCTA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharmin, R.A.; Karikari, B.; Bhuiyan, M.R.; Kong, K.; Yu, Z.; Zhang, C.; Zhao, T. Comparative Morpho-Physiological, Biochemical, and Gene Expressional Analyses Uncover Mechanisms of Waterlogging Tolerance in Two Soybean Introgression Lines. Plants 2024, 13, 1011. https://doi.org/10.3390/plants13071011
Sharmin RA, Karikari B, Bhuiyan MR, Kong K, Yu Z, Zhang C, Zhao T. Comparative Morpho-Physiological, Biochemical, and Gene Expressional Analyses Uncover Mechanisms of Waterlogging Tolerance in Two Soybean Introgression Lines. Plants. 2024; 13(7):1011. https://doi.org/10.3390/plants13071011
Chicago/Turabian StyleSharmin, Ripa Akter, Benjamin Karikari, Mashiur Rahman Bhuiyan, Keke Kong, Zheping Yu, Chunting Zhang, and Tuanjie Zhao. 2024. "Comparative Morpho-Physiological, Biochemical, and Gene Expressional Analyses Uncover Mechanisms of Waterlogging Tolerance in Two Soybean Introgression Lines" Plants 13, no. 7: 1011. https://doi.org/10.3390/plants13071011
APA StyleSharmin, R. A., Karikari, B., Bhuiyan, M. R., Kong, K., Yu, Z., Zhang, C., & Zhao, T. (2024). Comparative Morpho-Physiological, Biochemical, and Gene Expressional Analyses Uncover Mechanisms of Waterlogging Tolerance in Two Soybean Introgression Lines. Plants, 13(7), 1011. https://doi.org/10.3390/plants13071011