
Moving Away from 12:12; the Effect of Different Photoperiods on Biomass Yield and Cannabinoids in Medicinal Cannabis

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Reproductive Development
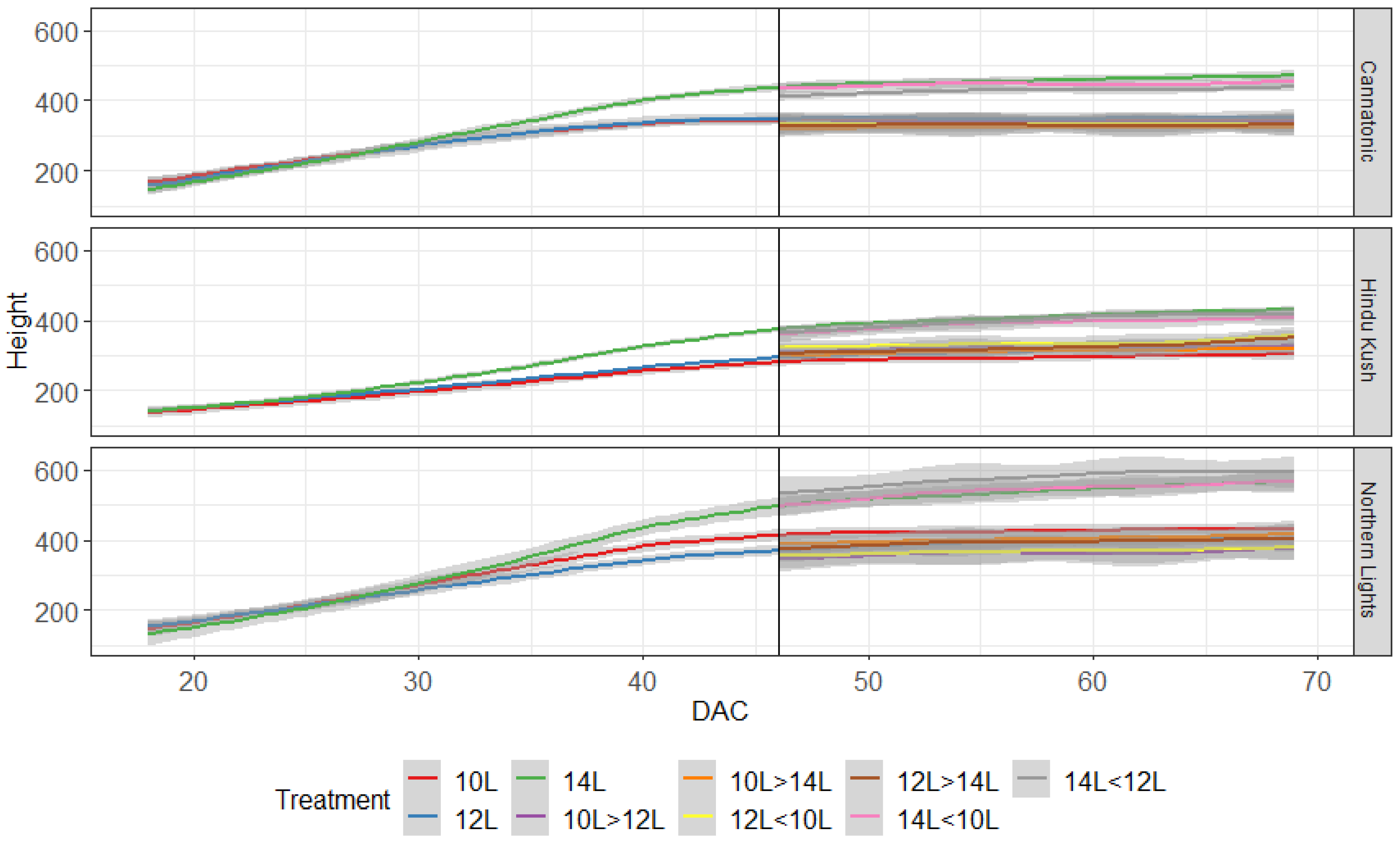
2.2. Plant Height
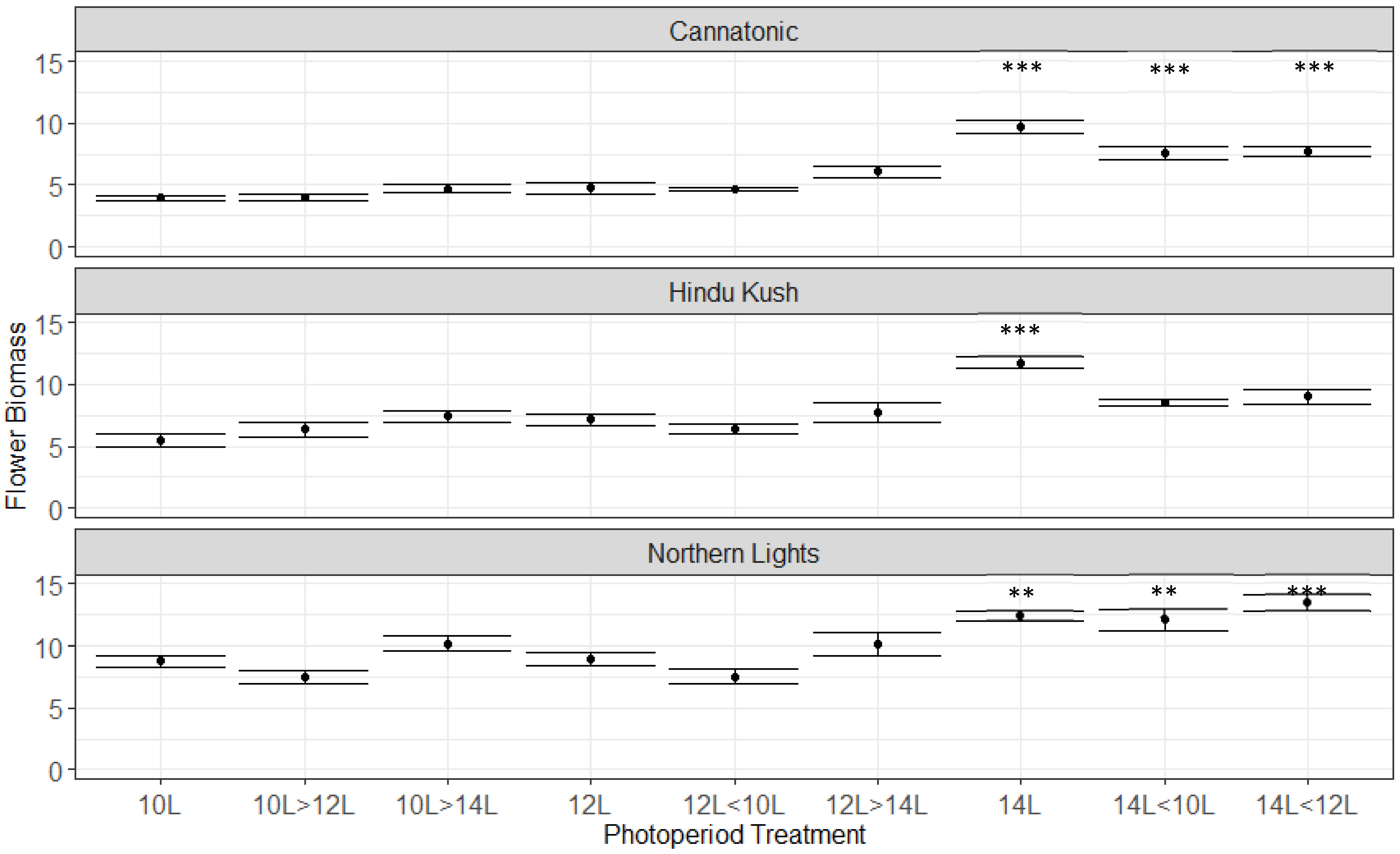
2.3. Flower Biomass Yield
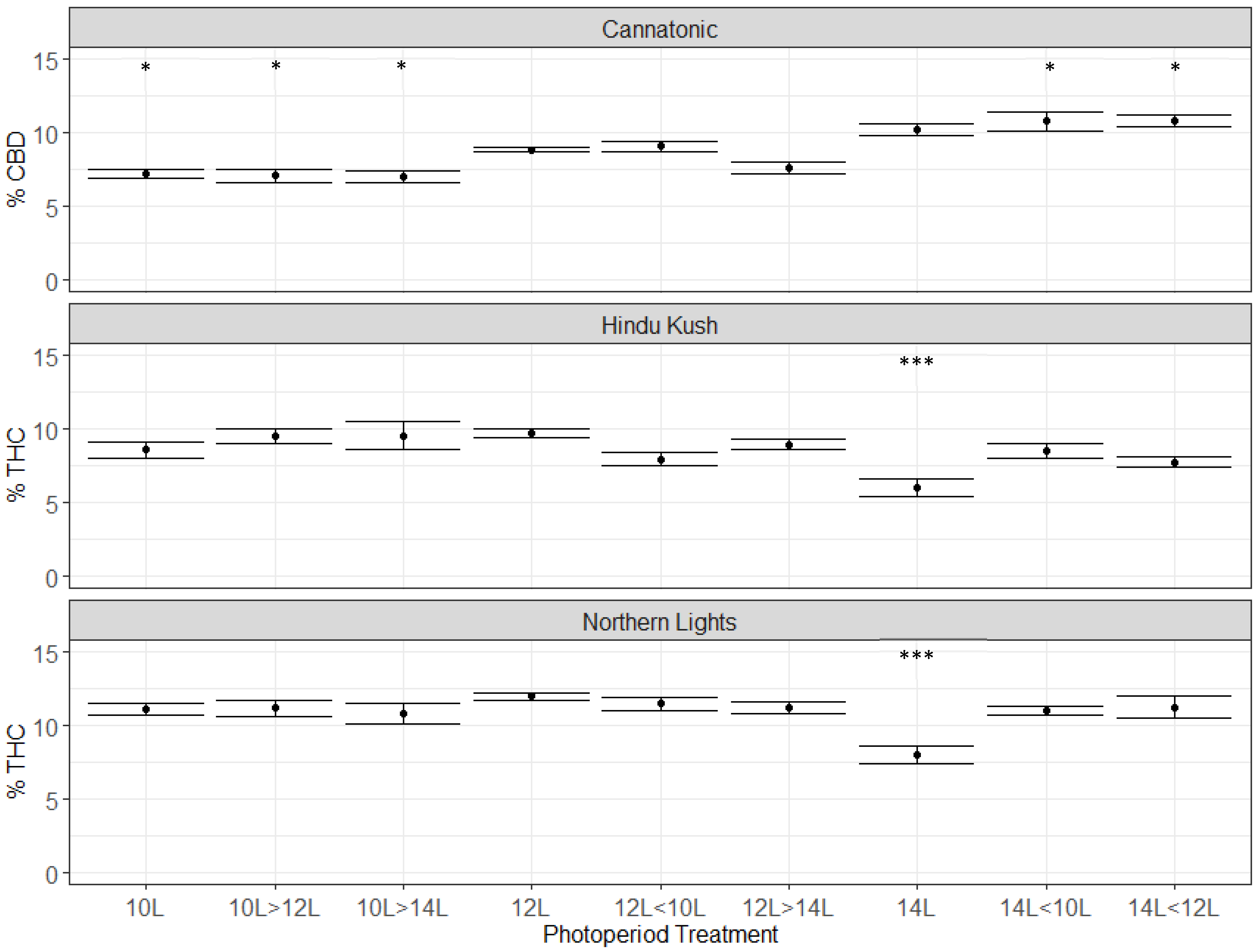
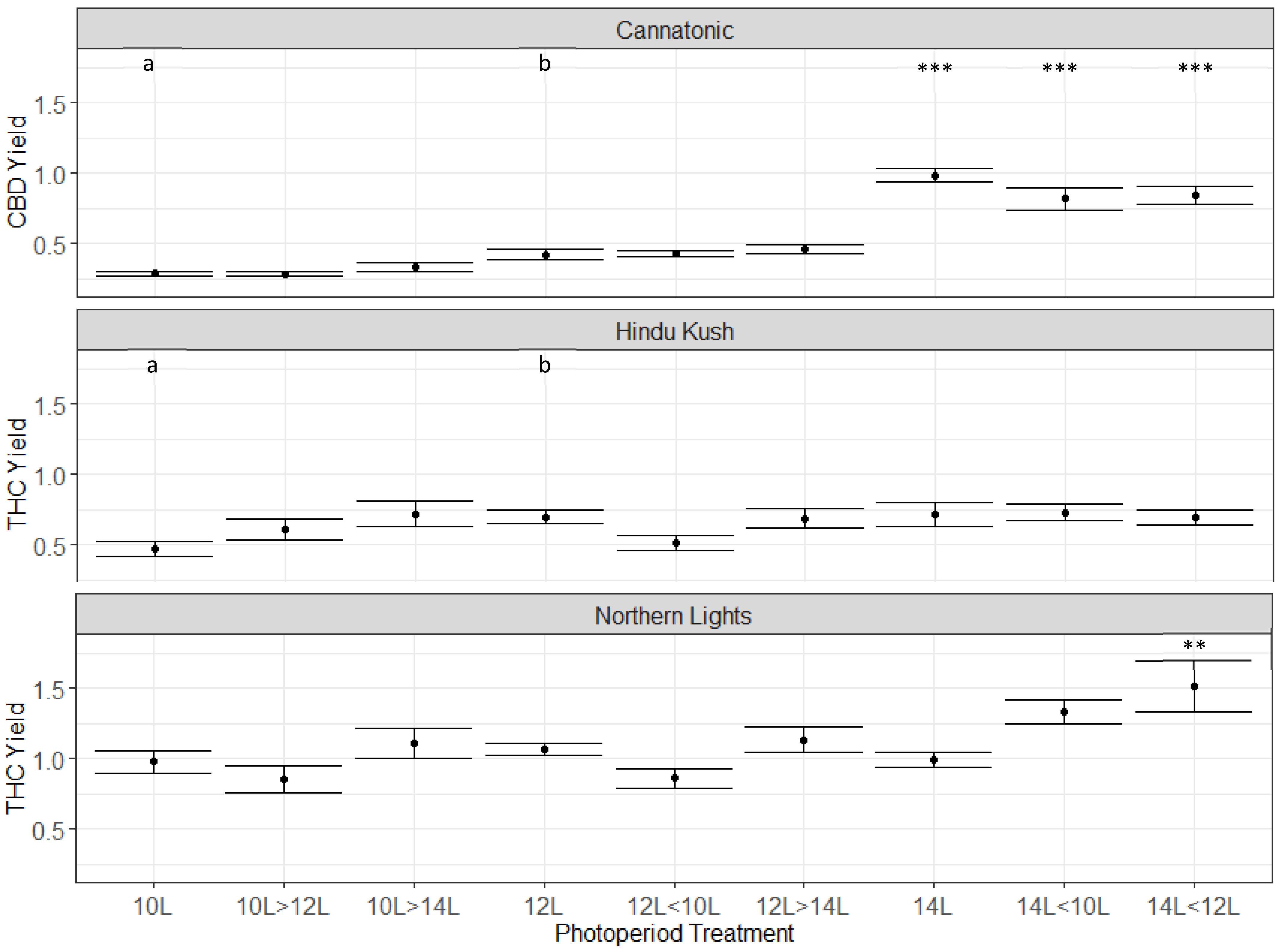
2.4. Cannabinoid Concentration
2.5. Physical Appearance of Flowers
2.6. Total Yield
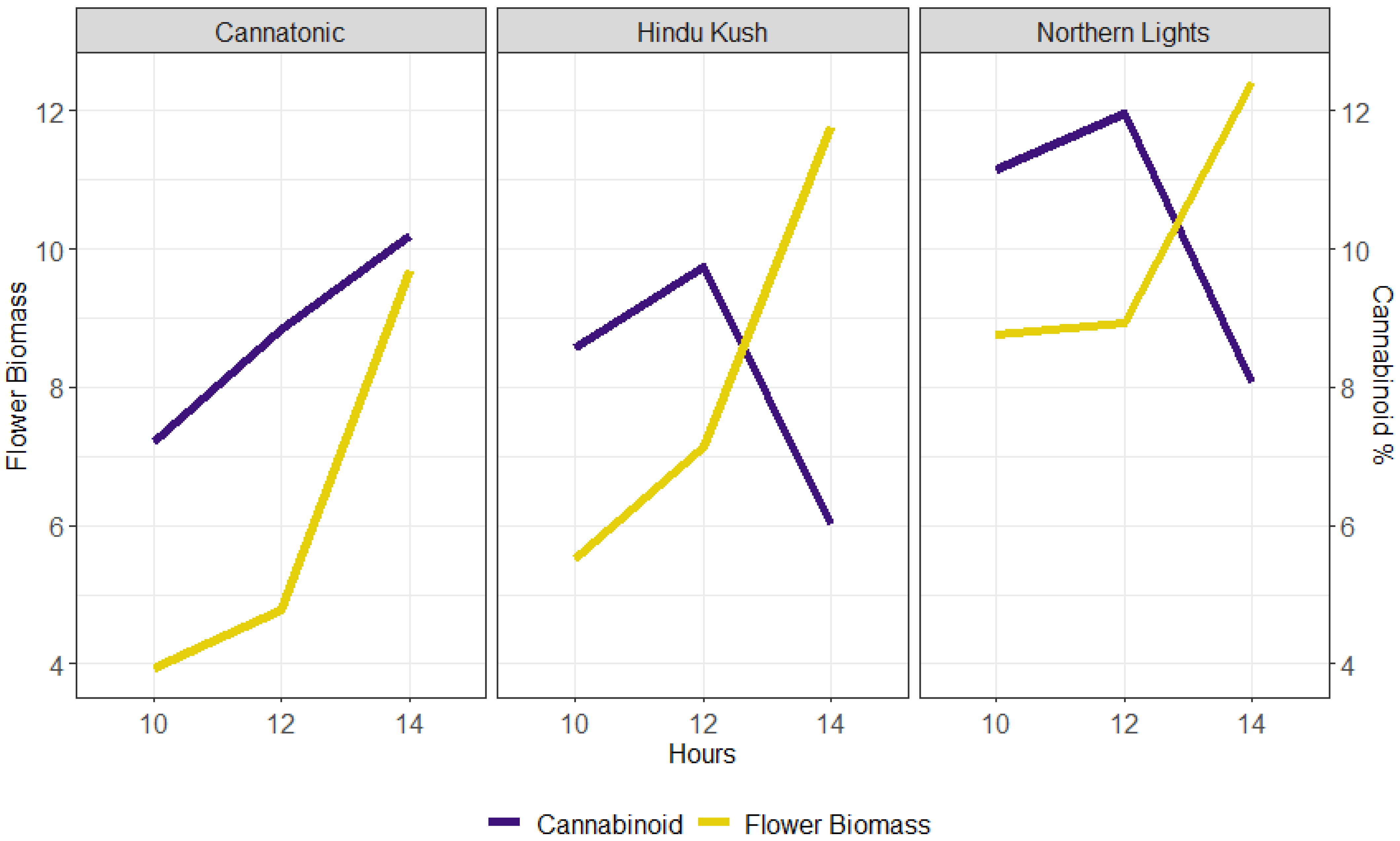
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growing Conditions
4.2. Treatments
4.3. Measurements
4.4. Analytics
4.5. Graphics and Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Livingston, S.J.; Rensing, K.H.; Page, J.E.; Samuels, A.L. A polarized supercell produces specialized metabolites in Cannabis trichomes. Curr. Biol. 2022, 32, 4040–4047. [Google Scholar] [CrossRef] [PubMed]
- Conneely, L.J.; Mauleon, R.; Mieog, J.; Barkla, B.J.; Kretzschmar, T. Characterization of the Cannabis sativa glandular trichome proteome. PLoS ONE 2021, 16, e0242633. [Google Scholar] [CrossRef] [PubMed]
- Brousseau, V.D.; Wu, B.S.; MacPherson, S.; Morello, V.; Lefsrud, M. Cannabinoids and Terpenes: How Production of Photo-Protectants Can Be Manipulated to Enhance Cannabis sativa L. Phytochemistry. Front. Plant. Sci. 2021, 12, 620021. [Google Scholar] [CrossRef]
- Chandra, S.; Lata, H.; ElSohly, M.A.; Walker, L.A.; Potter, D. Cannabis cultivation: Methodological issues for obtaining medical-grade product. Epilepsy Behav. 2017, 70, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Ben-Shabat, S. From gan-zi-gun-nu to anandamide and 2-arachidonoylglycerol: The ongoing story of cannabis. Nat. Prod. Rep. 1999, 16, 131–143. [Google Scholar] [CrossRef]
- Blake, A.; Wan, B.A.; Malek, L.; DeAngelis, C.; Diaz, P.; Lao, N.; Chow, E.; O’Hearn, S. A selective review of medical cannabis in cancer pain management. Ann. Palliat Med. 2017, 6, S215–S222. [Google Scholar] [CrossRef]
- Campbell, L.G.; Dufresne, J.; Sabatinos, S.A. Cannabinoid Inheritance Relies on Complex Genetic Architecture. Cannabis Cannabinoid Res. 2020, 5, 105–116. [Google Scholar] [CrossRef]
- Caplan, D.; Dixon, M.; Zheng, Y.B. Optimal Rate of Organic Fertilizer during the Flowering Stage for Cannabis Grown in Two Coir-based Substrates. Hortscience 2017, 52, 1796–1803. [Google Scholar] [CrossRef]
- Caplan, D.; Dixon, M.; Zheng, Y.B. Increasing Inflorescence Dry Weight and Cannabinoid Content in Medical Cannabis Using Controlled Drought Stress. Hortscience 2019, 54, 964–969. [Google Scholar] [CrossRef]
- Danziger, N.; Bernstein, N. Light matters: Effect of light spectra on cannabinoid profile and plant development of medical cannabis (Cannabis sativa L.). Ind. Crop. Prod. 2021, 164, 113351. [Google Scholar] [CrossRef]
- Reichel, P.; Munz, S.; Hartung, J.; Kotiranta, S.; Graeff-Honninger, S. Impacts of Different Light Spectra on CBD, CBDA and Terpene Concentrations in Relation to the Flower Positions of Different Cannabis Sativa L. Strains. Plants 2022, 11, 2695. [Google Scholar] [CrossRef] [PubMed]
- Dang, M.C.L.; Arachchige, N.M.; Campbell, L.G. Optimizing Photoperiod Switch to Maximize Floral Biomass and Cannabinoid Yield in Cannabis sativa L.: A Meta-Analytic Quantile Regression Approach. Front. Plant. Sci. 2022, 12, 797425. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, J. The Cannabis Encyclopedia: The Definitive Guide to Cultivation & Consumption of Medical Marijuana, Illustrated ed.; Van Patten Publishing: Vancouver, WA, USA, 2015. [Google Scholar]
- Moher, M.; Jones, M.; Zheng, Y.B. Photoperiodic Response of In Vitro Cannabis sativa Plants. Hortscience 2021, 56, 108–113. [Google Scholar] [CrossRef]
- Yang, S.; Cai, Y.; Liu, X.; Dong, M.; Zhang, Y.; Chen, S.; Zhang, W.; Li, Y.; Tang, M.; Zhai, X.; et al. A CsMYB6-CsTRY module regulates fruit trichome initiation in cucumber. J. Exp. Bot. 2018, 69, 1887–1902. [Google Scholar] [CrossRef] [PubMed]
- Hillig, K.W. Genetic evidence for speciation in Cannabis (Cannabaceae). Genet. Resour. Crop. Ev. 2005, 52, 161–180. [Google Scholar] [CrossRef]
- Zhang, M.; Anderson, S.L.; Brym, Z.T.; Pearson, B.J. Photoperiodic Flowering Response of Essential Oil, Grain, and Fiber Hemp (Cannabis sativa L.) Cultivars. Front. Plant. Sci. 2021, 12, 694153. [Google Scholar] [CrossRef]
- Zhang, Q.Y.; Chen, X.; Guo, H.Y.; Trindade, L.M.; Salentijn, E.M.J.; Guo, R.; Guo, M.B.; Xu, Y.P.; Yang, M. Latitudinal Adaptation and Genetic Insights Into the Origins of Cannabis sativa L. Front. Plant. Sci. 2018, 9, 1876. [Google Scholar] [CrossRef]
- The Hemp Mine. Hemp Planting Map. Available online: https://thehempmine.com/pages/hemp-planting-map (accessed on 28 October 2022).
- Sikora, V.; Berenji, J.; Latković, D. Influence of agroclimatic conditions on content of main cannabinoids in industrial hemp (Cannabis sativa L.). Genetics 2011, 43, 449–456. [Google Scholar] [CrossRef]
- Purdy, S.J.; Hewavitharana, A.K.; Halimi, R.A.; Magner, N.J.; Peterswald, T.J.; Trebilco, A.; Kretzschmar, T.; Hailstones, D. A One-Step Grafting Methodology Can Adjust Stem Morphology and Increase THCA Yield in Medicinal Cannabis. Agronomy 2022, 12, 852. [Google Scholar] [CrossRef]
- Kojima, S.; Takahashi, Y.; Kobayashi, Y.; Monna, L.; Sasaki, T.; Araki, T.; Yano, M. Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowering downstream of Hd1 under short-day conditions. Plant Cell Physiol. 2002, 43, 1096–1105. [Google Scholar] [CrossRef]
- Suarez-Lopez, P.; Wheatley, K.; Robson, F.; Onouchi, H.; Valverde, F.; Coupland, G. CONSTANS mediates between the circadian clock and the control of flowering in Arabidopsis. Nature 2001, 410, 1116–1120. [Google Scholar] [CrossRef] [PubMed]
- Yano, M.; Kojima, S.; Takahashi, Y.; Lin, H.X.; Sasaki, T. Genetic control of flowering time in rice, a short-day plant. Plant Physiol. 2001, 127, 1425–1429. [Google Scholar] [CrossRef] [PubMed]
- Hayama, R.; Coupland, G. The molecular basis of diversity in the photoperiodic flowering responses of Arabidopsis and rice. Plant Physiol. 2004, 135, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Frank, M. The Marijuana Growers Guide Delux; And/Or Press: Berkley, CA, USA, 1978. [Google Scholar]
- Park, S.-H.; Pauli, C.S.; Gostin, E.L.; Staples, S.K.; Seifried, D.; Kinney, C.; Vanden Heuvel, B.D. Effects of short-term environmental stresses on the onset of cannabinoid production in young immature flowers of industrial hemp (Cannabis sativa L.). J. Cannabis Res. 2022, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Naim-Feil, E.; Pembleton, L.W.; Spooner, L.E.; Malthouse, A.L.; Miner, A.; Quinn, M.; Polotnianka, R.M.; Baillie, R.C.; Spangenberg, G.C.; Cogan, N.O.I. The characterization of key physiological traits of medicinal cannabis (Cannabis sativa L.) as a tool for precision breeding. BMC Plant Biol. 2021, 21, 294. [Google Scholar] [CrossRef]
- Bustan, A.; Goldschmidt, E.E. Estimating the cost of flowering in a grapefruit tree. Plant Cell Environ. 1998, 21, 217–224. [Google Scholar] [CrossRef]
- Dovis, V.L.; Machado, E.C.; Ribeiro, R.V.; Magalhaes, J.R.; Marchiori, P.E.R.; Sales, C.R.G. Roots are important sources of carbohydrates during flowering and fruiting in ‘Valencia’ sweet orange trees with varying fruit load. Sci. Hortic. Amsterdam 2014, 174, 87–95. [Google Scholar] [CrossRef]
- Tang, F.Y.; Gao, X.; Peng, J.J. The dynamics of carbohydrate and associated gene expression in the stems and roots of upland cotton (Gossypium hirsutum L.) during carbon remobilization. Plant Physiol. Bioch. 2022, 178, 125–136. [Google Scholar] [CrossRef]
- Telfer, A.; Bollman, K.M.; Poethig, R.S. Phase change and the regulation of trichome distribution in Arabidopsis thaliana. Development 1997, 124, 645–654. [Google Scholar] [CrossRef]
- Chien, J.C.; Sussex, I.M. Differential regulation of trichome formation on the adaxial and abaxial leaf surfaces by Gibberellins and photoperiod in Arabidopsis thaliana (L) Heynh. Plant Physiol. 1996, 111, 1321–1328. [Google Scholar] [CrossRef]
- Stack, G.M.; Toth, J.A.; Carlson, C.H.; Cala, A.R.; Marrero-Gonzalez, M.I.; Wilk, R.L.; Gentner, D.R.; Crawford, J.L.; Philippe, G.; Rose, J.K.C.; et al. Season-long characterization of high-cannabinoid hemp (Cannabis sativa L.) reveals variation in cannabinoid accumulation, flowering time, and disease resistance. GCB Bioenergy 2021, 13, 546–561. [Google Scholar] [CrossRef]
- Toth, J.A.; Stack, G.M.; Carlson, C.H.; Smart, L.B. Identification and mapping of major-effect flowering time loci Autoflower1 and Early1 in Cannabis sativa L. Front. Plant. Sci. 2022, 13, 991680. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Ogiso-Tanaka, E.; Matsubara, K.; Yamanouchi, U.; Ebana, K.; Yano, M. Hd16, a gene for casein kinase I, is involved in the control of rice flowering time by modulating the day-length response. Plant. J. 2013, 76, 36–46. [Google Scholar] [CrossRef] [PubMed]
- ElSohly, M.A.; Mehmedic, Z.; Foster, S.; Gon, C.; Chandra, S.; Church, J.C. Changes in Cannabis Potency Over the Last 2 Decades (1995–2014): Analysis of Current Data in the United States. Biol. Psychiatry 2016, 79, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Smart, R.; Caulkins, J.P.; Kilmer, B.; Davenport, S.; Midgette, G. Variation in cannabis potency and prices in a newly legal market: Evidence from 30 million cannabis sales in Washington state. Addiction 2017, 112, 2167–2177. [Google Scholar] [CrossRef] [PubMed]
- Wallace, M.S.; Marcotte, T.D.; Umlauf, A.; Gouaux, B.; Atkinson, J.H. Efficacy of Inhaled Cannabis on Painful Diabetic Neuropathy. J. Pain 2015, 16, 616–627. [Google Scholar] [CrossRef] [PubMed]
- Abrams, D.I.; Jay, C.A.; Shade, S.B.; Vizoso, H.; Reda, H.; Press, S.; Kelly, M.E.; Rowbotham, M.C.; Petersen, K.L. Cannabis in painful HIV-associated sensory neuropathy—A randomized placebo-controlled trial. Neurology 2007, 68, 515–521. [Google Scholar] [CrossRef]
- Cash, M.C.; Cunnane, K.; Fan, C.Y.; Romero-Sandoval, E.A. Mapping cannabis potency in medical and recreational programs in the United States. PLoS ONE 2020, 15, e0230167. [Google Scholar] [CrossRef]
- PharmOut Pty Ltd. Australian Medicinal Cannabis Cultivation Indoors or Outdoors Debate. Available online: https://www.pharmout.net/medicinal-cannabis-cultivation-indoors-or-outdoors/ (accessed on 10 January 2023).
- Summers, H.M.; Sproul, E.; Quinn, J.C. The greenhouse gas emissions of indoor cannabis production in the United States. Nat. Sustain. 2021, 4, 644–650. [Google Scholar] [CrossRef]
- Hewavitharana, A.K.; Gloerfelt-Tarp, F.; Nolan, M.; Barkla, B.J.; Purdy, S.; Kretzschmar, T. Simultaneous Quantification of 17 Cannabinoids in Cannabis Inflorescence by Liquid Chromatography-Mass Spectrometry. Separations 2022, 9, 85. [Google Scholar] [CrossRef]
- R Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2020. [Google Scholar]
- Dunnett, C.W. A multiple comparison procedure for comparing several treatments with a control. J. Am. Stat. Assoc. 1955, 50, 1096–1121. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % of Plants with Pistils | % Plants with Trichomes | ||||||
|---|---|---|---|---|---|---|---|
| Genotype | Photoperiod | DAC 25 | DAC_34 | DAC_25 | DAC_34 | DAC_41 | DAC_46 |
| Cannatonic | 10L | 0 | 100 | 0 | 0 | 100 | 100 |
| 12L | 0 | 100 | 0 | 100 | 100 | 100 | |
| 14L | 0 | 100 | 0 | 80 | 100 | 100 | |
| Northern Lights | 10L | 60 | 100 | 0 | 0 | 26.6 | 100 |
| 12L | 66.6 | 100 | 0 | 40 | 100 | 100 | |
| 14L | 73.3 | 100 | 0 | 33.3 | 100 | 100 | |
| Hindu Kush | 10L | 0 | 100 | 0 | 0 | 26.6 | 100 |
| 12L | 20 | 100 | 0 | 0 | 100 | 100 | |
| 14L | 20 | 100 | 0 | 6.6 | 100 | 100 | |
| Treatment 1 (Light/Dark) | Treatment 2 (Light/Dark) | Name | Description |
|---|---|---|---|
| 10L:14D | 10L:14D | 10L | 10L:14D for duration of experiment |
| 10L:14D | 12L:12D | 10L > 12L | Increased photoperiod from 10L to 12L (+2 h light) |
| 10L:14D | 14L:10D | 10L > 14L | Increased photoperiod from 10L to 14L (+4 h light) |
| 12L:12D | 12L:12D | 12L | 12L:12D for duration of experiment |
| 12L:12D | 10L:14D | 12L < 10L | Decreased photoperiod from 12L to 10L (−2 h light) |
| 12L:12D | 14L:10D | 12L > 14L | Increased photoperiod from 12L to 14L (+2 h light) |
| 14L:10D | 14L:10D | 14L | 14L:10D for duration of experiment |
| 14L:10D | 10L:14D | 14L < 10L | Decreased photoperiod from 14L to 10L (−4 h light) |
| 14L:10D | 12L:12D | 14L < 12L | Decreased photoperiod from 14L to 12L (−2 h light) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peterswald, T.J.; Mieog, J.C.; Azman Halimi, R.; Magner, N.J.; Trebilco, A.; Kretzschmar, T.; Purdy, S.J. Moving Away from 12:12; the Effect of Different Photoperiods on Biomass Yield and Cannabinoids in Medicinal Cannabis. Plants 2023, 12, 1061. https://doi.org/10.3390/plants12051061
Peterswald TJ, Mieog JC, Azman Halimi R, Magner NJ, Trebilco A, Kretzschmar T, Purdy SJ. Moving Away from 12:12; the Effect of Different Photoperiods on Biomass Yield and Cannabinoids in Medicinal Cannabis. Plants. 2023; 12(5):1061. https://doi.org/10.3390/plants12051061
Chicago/Turabian StylePeterswald, Tyson James, Jos Cornelis Mieog, Razlin Azman Halimi, Nelson Joel Magner, Amy Trebilco, Tobias Kretzschmar, and Sarah Jane Purdy. 2023. "Moving Away from 12:12; the Effect of Different Photoperiods on Biomass Yield and Cannabinoids in Medicinal Cannabis" Plants 12, no. 5: 1061. https://doi.org/10.3390/plants12051061
APA StylePeterswald, T. J., Mieog, J. C., Azman Halimi, R., Magner, N. J., Trebilco, A., Kretzschmar, T., & Purdy, S. J. (2023). Moving Away from 12:12; the Effect of Different Photoperiods on Biomass Yield and Cannabinoids in Medicinal Cannabis. Plants, 12(5), 1061. https://doi.org/10.3390/plants12051061