1. Introduction
Cannabis sativa L. (cannabis) is a multipurpose plant species known for its psychoactive and medicinal properties that has gained significant attention worldwide [
1]. The increasing global acceptance of cannabis drives the necessity of scientific research aimed at understanding its biology for optimizing cultivation practices [
2]. Cannabis contains a diverse array of chemical compounds, including cannabinoids, which are responsible for its therapeutic effects [
3]. The concentration and composition of cannabinoids varies between cannabis cultivars, leading to distinct chemical profiles and differential biological effects [
4]. In addition to cannabinoid content, morphological traits play a crucial roles in the overall physiology, yield potential, and quality of cannabis plants [
5].
The morphological traits of cannabis encompass various aspects, including leaf morphology, plant architecture, and trichome characteristics [
6]. Leaf morphology plays a crucial role in the overall physiology and performance of plants [
7]. Cannabis leaves represent primary photosynthetic organs, where light is captured and converted into chemical energy [
8]. The morphology of leaves, including their size, shape, and structure, directly influence the plant’s ability to efficiently capture light, exchange gases, and regulate transpiration [
9]. Leaf morphology is tightly linked to key physiological processes such as water and nutrient uptake, carbon assimilation, and stress responses [
10]. Moreover, leaf traits are often used as indicators of plant health, growth vigor, and productivity in agricultural and horticultural practices [
7]. Understanding the emergent variations in cannabis leaf morphology and their implications can provide valuable insights for cultivation practices [
6]. Therefore, comprehensive cannabis leaf morphology studies are essential in better understanding cannabis physiology for improved cultivation and medicinal applications.
The architecture of cannabis plants, including plant height, branching pattern, and overall structure, plays a significant role in determining their growth, productivity, and adaptability [
6]. Cannabis exhibits considerable phenotypic plasticity, with variations in plant architecture observed among different cultivars and environmental conditions [
6]. Plant height influences light interception, resource utilization, and biomass production, thereby impacting overall yield potential [
11,
12]. The branching pattern, including the number and arrangement of lateral branches, affects the distribution of resources, flower development, and cannabinoid synthesis [
11]. The architecture of cannabis plants also influences pest and disease resistance, as well as the ease of cultivation, harvesting, and management practices [
6]. Understanding the factors that influence cannabis architecture can inform producers to develop cultivars with desired traits, guide in optimizing cultivation techniques, and ultimately maximize the quality and quantity of cannabinoid production [
6].
Glandular trichomes are structures found on the surfaces of cannabis plants that play a pivotal role in the biosynthesis and accumulation of the bioactive compounds responsible for the plant’s medicinal and recreational properties [
13]. The intricate morphology and characteristics of cannabis trichomes have significant implications for the production and quality of cannabinoids, terpenes, and other secondary metabolites [
14]. Trichome density, size, and structure directly relate to the potency, flavor, aroma, and therapeutic effects of cannabis products [
15]. Furthermore, trichomes are thought to serve as a defense mechanism against environmental stressors and herbivory, acting as both physical barriers and through the production of chemical compounds that deter pests [
16]. Understanding the variations in trichome characteristics throughout the cannabis life cycle can provide valuable insights into optimizing cultivation practices, strain selection, and the production of cannabis-based products with consistent and desirable attributes [
17]. Therefore, a deeper understanding of cannabis trichome characteristics is crucial for interpreting the plant’s chemical ecology, improving cultivation techniques, and meeting the demands of the growing cannabis industry.
While there is a wealth of research exploring the morphological traits of cannabis, most studies focus on specific growth stages. Thus, a comprehensive investigation of the complete life cycle of specific drug-type cultivars is warranted. Such studies will shed light on the morphological variations that occur from seed germination to flower harvesting.
This study aimed to characterize various morphological traits, including leaf morphology, plant height, and trichome features, throughout the life cycle of a popular drug-type cannabis cultivar, White Widow. By focusing on a specific cultivar, detailed insights into the morphological dynamics and their potential implications for cannabinoid production and plant quality can be achieved. White Widow, a widely popular cannabis cultivar with breeders and connoisseurs, is known to relay potent psychoactive effects. As such, it provides an excellent model to investigate the morphological changes occurring throughout development that are associated with drug-type cannabis [
18]. The findings from this study will contribute to the depth of knowledge surrounding the morphological plasticity of cannabis, providing practical and fundamental insight for cultivators and researchers in the cannabis industry. The information presented can be used to inform cultivation practices aimed at improving desirable morphological traits and cannabinoid profiles.
3. Discussion
Cannabis is known to exhibit considerable variation in growth habits, leaf morphology, flowering time, cannabinoid content, and secondary metabolite synthesis throughout the plant life cycle [
8,
19]. Understanding the mechanisms driving such morphological changes in cannabis is essential for optimizing cultivation practices and harnessing the plant’s bioactive compounds for medicinal and industrial purposes [
6,
11,
20,
21]. While several studies [
6,
11,
17,
21] have compared cannabis morphology in various contexts, this study is fundamentally different, delving into the ontological development of cannabis to provide a comprehensive examination of the morphological characteristics emerging throughout its life cycle. This holistic approach not only enhances our understanding of this plant but also holds several implications for horticultural practices. This type of study unravels the intricate biological processes governing growth and development to shed light on the emergent transitions and adaptations undergone throughout the cannabis life cycle [
22]. Insight into the plant’s morphological changes can serve as a valuable roadmap for horticulturists seeking to optimize cultivation practices. For instance, understanding the stages of juvenility in cannabis is crucial for improving rooting success, facilitating micropropagation establishment, and enhancing overall plant vigor. Additionally, understanding the process of transition to maturity and floral development can guide commercial practices for the production of floral biomass.
Moreover, the findings of the current study hold immense potential to advance the broader field of cannabis research. The morphological clues presented provide the foundation for further investigations related to gene regulation and transcriptome profiles at different developmental stages of the cannabis life cycle. This fundamental knowledge is indispensable for conducting more sophisticated studies on the genetic underpinnings of cannabis biology, enabling researchers to explore the plant’s relevance to various applications, from medicinal to industrial, with greater precision and efficacy.
Cannabis undeniably showcases some of the most recognizable leaves in the botanical realm [
7]. Characterized by palmately compound structures encompassing a diverse range of leaflet numbers, these leaves have ingrained themselves as symbols within popular culture [
7]. The extensive range of leaf morphology has been previously documented by Quimby et al. [
23], and subsequent work by Anderson [
24] provided the pioneering quantification of central leaflet dimensions, including length, width, and proportional ratios.
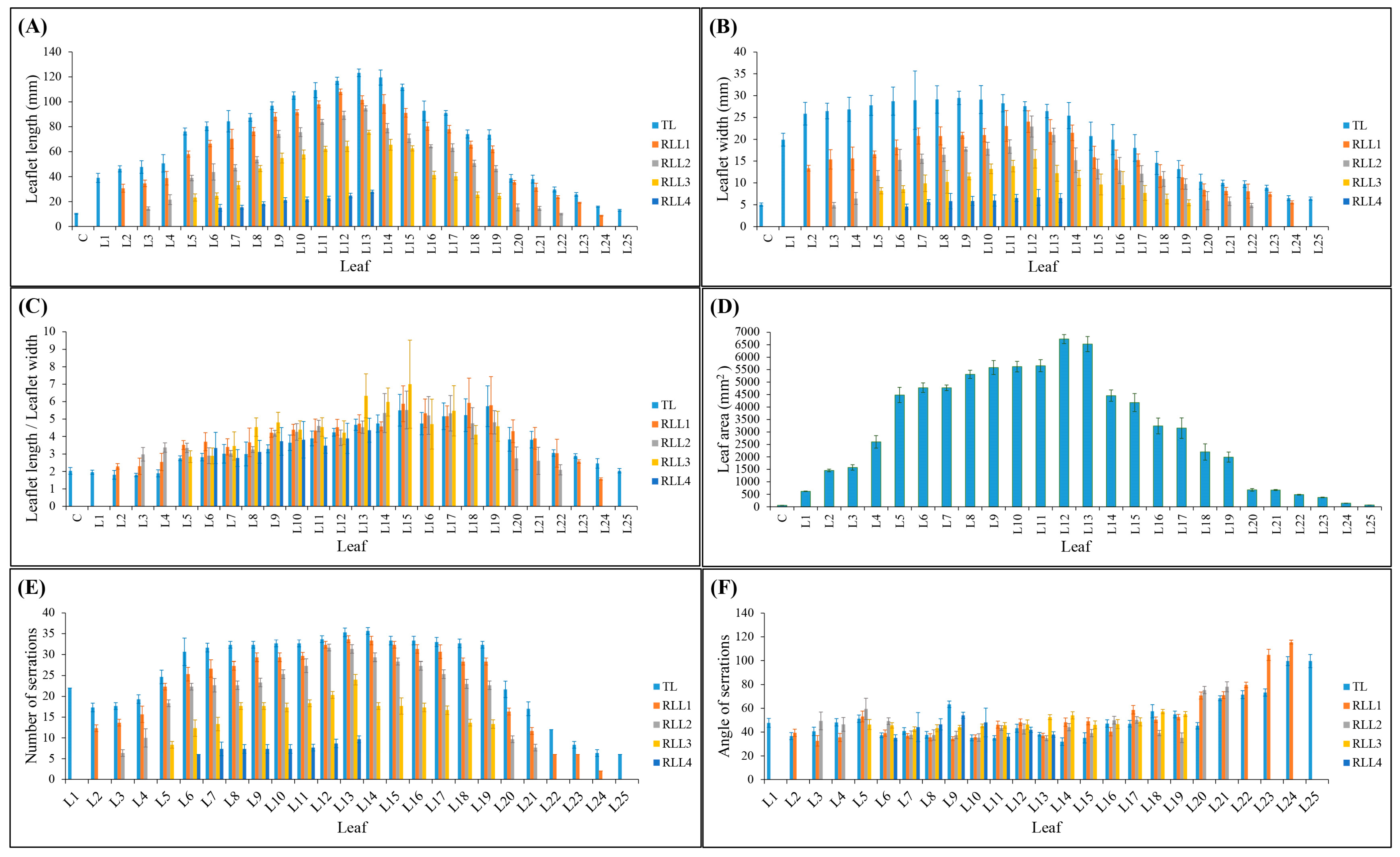
The intriguing developmental pattern observed in the number of leaflets within cannabis leaves represents one of the more interesting morphological properties presented. The transition from single leaflet leaves to leaves with nine leaflets during the mature stage, then reversion back to leaves of one leaflet during the flowering stage, prompts a comprehensive investigation into the underlying factors governing this phenomenon. The shift in leaflet count reflects a dynamic response to the plant’s developmental requirements and environmental cues [
25,
26].
During the early stages of growth, the gradual increase in the leaflet number, length, and width of terminal and lateral leaflets, as well as leaf area, can be attributed to the plant’s strategy to maximize photosynthetic efficiency and resource capture [
9]. The progression to leaves with nine serrated leaflets (L6 to L13) indicates a stable phase of optimized photosynthesis, suggesting that the plant has reached an equilibrium between energy production and consumption [
9]. This developmental phase could also be influenced by hormonal changes that dictate leaf patterning and growth [
25,
26].
The subsequent reduction in leaflet number, length, width of terminal and lateral leaflets, and leaf area as the cannabis plant transitions to its flowering stage may be attributed to the reallocation of resources towards reproductive structures. The energy-intensive process of flowering necessitates a redirection of nutrients and resources towards flower and seed production, potentially leading to a reduction in leaf complexity [
9]. The shift from leaves with nine serrated leaflets (L6 to L13) to leaves with decreasing leaflet numbers (L14 to L25) and overall leaf area suggests a trade-off between vegetative growth and reproductive efforts [
17]. The reversion to leaves with a single serrated leaflet (L25) at the flowering stage could be explained by the plant’s prioritization of reproductive success over vegetative expansion [
9]. Alterations in morphological traits of cannabis leaves throughout various developmental stages exhibit congruence with observations in diverse plant species [
8]. This consistency underscores fundamental patterns in leaf development that extend beyond taxonomic boundaries. For instance, the increase in the number of leaflets observed in cannabis during vegetative growth is analogous to species such as hairy bittercress and tomato, revealing a shared mechanism governing leaflet proliferation during this phase [
27,
28,
29]. Furthermore, the changes in leaf size and shape observed during cannabis’ transition from the vegetative to reproductive stages mirror the phenomena documented in other plant species, emphasizing the broader applicability of these findings and suggesting a conserved regulatory framework governing leaf morphogenesis in plants [
17,
25,
26]. This cross-species consistency in leaf morphological traits enhances the general understanding of cannabis development while contributing to the broader knowledge of plant biology.
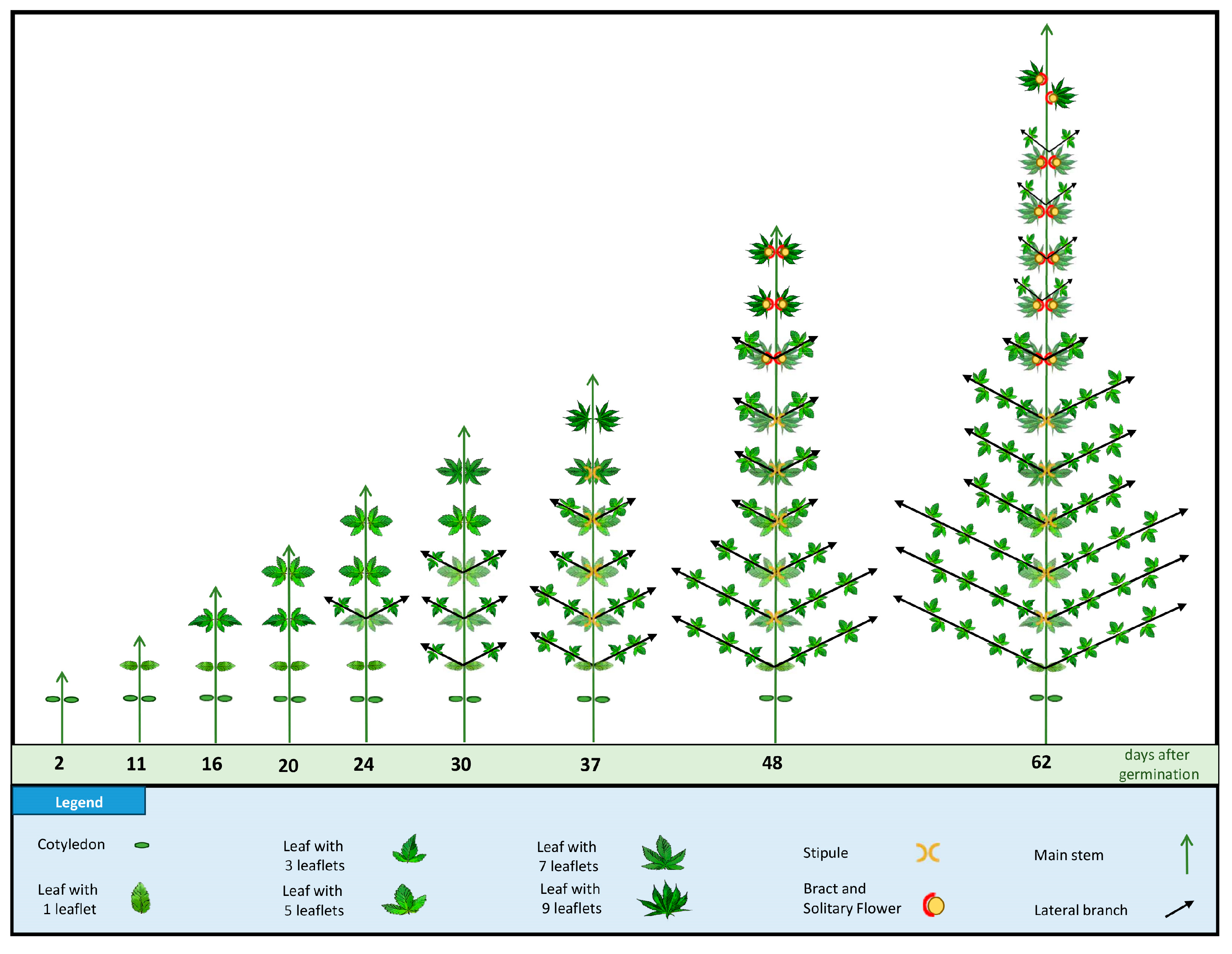
Under long-day photoperiod conditions, the developmental progression of cannabis plants is distinctly outlined. Nodes 2–4 displayed the presence of axillary buds and leaves, indicating the juvenile phase. Nodes 5 and 6 exhibited the emergence of stipules flanking the base of the leaf petiole, signifying the transition from juvenility to maturity. The shift to the reproductive phase was marked by nodes 7–12, and characterized by distinct elements such as foliar leaves, bracts, and solitary flowers (colloquially referred to as pre-flowers). In line with the results of the current study, previous studies showed that, under long-day photoperiod conditions, cannabis plants may undergo differentiation to show initial solitary flowers within the fourth to seventh internodal regions [
17,
22,
30]. The shift in phyllotaxy from an opposite to an alternate arrangement further signifies a developmental shift in the plant [
30]. The change in leaflet count and phyllotaxy in the axillary shoots emphasizes the dynamic nature of leaf development during different growth stages [
10]. Furthermore, the current investigation presents information that can be used for cannabis rejuvenation through tissue culture. It is well-documented that tissue culture techniques can effectively rejuvenate aging plants, prompting them to revert to a juvenile state, often accompanied by the re-establishment of characteristics such as opposite leaf arrangement [
2,
20]. This capability has profound implications for plant propagation, the maintenance of genetic integrity, and the potential enhancement of overall crop yields [
20,
31].
The emergence of solitary flowers during long-day conditions has complicated the characterization of the phase transition from the vegetative to reproductive stages in cannabis. Despite the designation of a long-day photoperiod as ‘non-inductive’ for cannabis, the presence of solitary flowers under a long-day photoperiod shows a phase transition in cannabis plants [
17]. The initiation of these solitary flowers is believed to be influenced by the plant’s age and regulated by intrinsic cues rather than being governed by the photoperiod [
22]. Since solitary flowers can manifest under both extended and shortened photoperiods, it has been suggested that cannabis displays a day-neutral response concerning this facet of flower induction [
22]. However, the question of whether the initiation of solitary flowers marks the culmination of the vegetative phase remains unresolved. Vegetative development may persist at the shoot apical meristem (SAM) during the interval between the appearance of solitary flowers and the ultimate induction of inflorescence development [
30].
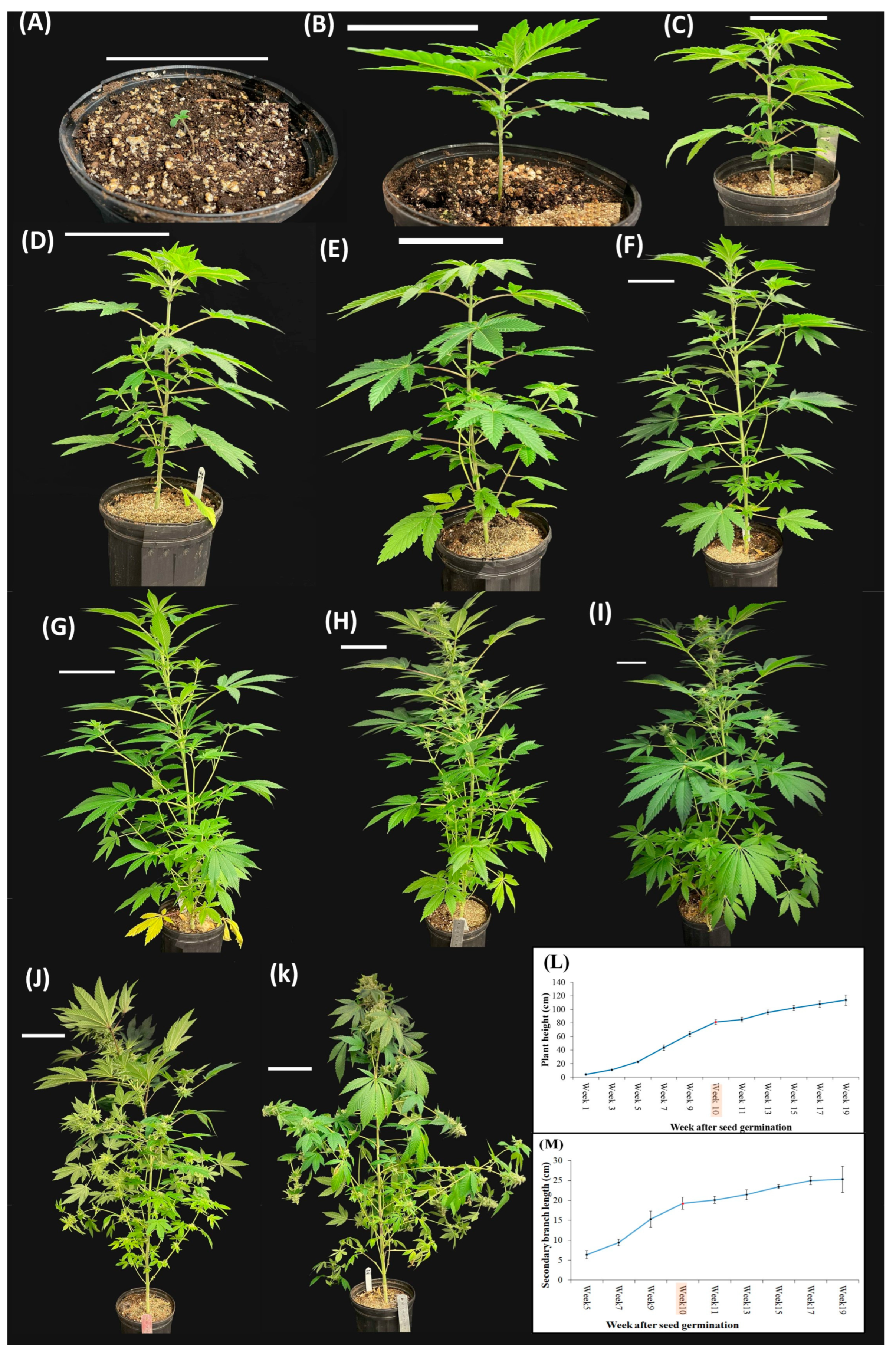
In the current study, the transition to a short-day photoperiod initiated a distinct set of morphological changes. The precise recognition of inflorescence onset, marked by the appearance of multiple pairs of stigmas, allowed the accurate delineation of the inflorescence initiation phase. After 10 days of short-day photoperiod exposure, inflorescences became evident at node 15. At full flowering (15 days of short-day photoperiod), the main inflorescences were observed at the apical extremities of the primary and secondary branches. The onset of inflorescence development is characterized by alterations in the structural configuration of the shoot apex [
17]. This transformation results in the development of a complex raceme featuring densely branched appendages and recurrent phytomer structures. These phytomer formations encompass an internode, a foliar leaf (sustained by a petiole), bracts, and solitary flowers (comprising the stigma, style, and perigonal bract) [
17].
The intricacy of morphophysiological attributes linked to flowering patterns in cannabis has resulted in the use of variable terms when documenting these characteristics [
5,
17,
32]. Previous investigations have postulated the occurrence of four primary events during florogenesis: (1) the initiation of solitary flowers, (2) the emergence of axillary branches and the shift towards more complex branching structures, (3) the initiation of inflorescences, characterized by the formation of flower clusters at both the shoot apex and axillary branches, and (4) terminal flowering, signifying the transformation of the apical meristem into a terminal flower [
22,
30]. Alterations in the shoot apex architecture and the onset of inflorescence flowering have demonstrated inducibility by short-day photoperiods, with evidence suggesting that these traits are regulated independently from the formation of solitary flowers [
17].
Upon exposure to the short-day photoperiods, cannabis plants transitioned to the reproductive phase, characterized by the emergence of compacted inflorescences in the apical regions of both the main stem, secondary-, and tertiary-order branches. The complex branched raceme structure of the cannabis inflorescence is a hallmark of monopodial growth, featuring a persistent apical meristem and indeterminate axillary inflorescences of higher orders [
17]. This branching pattern reflects the plant’s strategy to maximize floral development potential through repeated branching and reiteration, enabling the production of numerous reproductive structures [
17]. The emergence of glandular trichomes on the perigonal bracts encasing the ovary during flower development signifies the initiation of resin production, a characteristic feature of cannabis flowers [
15]. The transformation of the stigma color from whitish-yellow to reddish-brown as the inflorescences mature/senesce underscores the physiological changes that occur during flower development. This transition may correspond to the maturation of the flower [
14,
15]. The presence of paired female inflorescences within the leaf axils, associated with smaller branchlets of secondary axillary branches, demonstrates the branching pattern of cannabis inflorescences and the consistent phytomer structure of the higher-order branchlets [
17].
Sexual dimorphism (i.e., the distinct physical and physiological differences between males and females of a species) is a fascinating and intricate phenomenon in cannabis [
33]. Cannabis exhibits a karyotype composed of a single pair of heteromorphic sex chromosomes and nine pairs of homomorphic autosomal chromosomes [
34]. Cannabis plants, especially genotypes used for medicinal/recreational purposes, generally display dioecious attributes, characterized by well-defined male and female individuals [
34]. Nonetheless, the inherent flexibility in sexual expression can give rise to the emergence of hermaphroditic plants, recognized as monoecious phenotypes [
33]. Female cannabis plants have pistillate flowers densely clustered and interspersed with leafy bracts [
33]. Some dioecious plants are known to become hermaphroditic and produce male flowers [
33]. An interesting observation in the current study was the appearance of male flowers in various plants after a prolonged period. These flowers only appeared long after the plants would have normally been harvested in a commercial setting. Specifically, male flowers were observed at day 76 of the short-day photoperiod when this cultivar is general considered mature at day 56 of the short-day photoperiod. The emergence of male flowers in the late stages of senescence in an unpollinated female plant is a logical adaptation by the plant as a last effort to produce seeds. The late-stage transition into hermaphroditism underscores the genetic diversity and potential for both male and female reproductive structures to emerge from within the same plant, implications of which can be used to guide cultivation practices [
33,
35].
The investigation of trichome density on both the abaxial and adaxial sides of the bract revealed a notable increase from week 2 to week 8. This temporal trend suggests that, as the plants progressed in age, there was consistent generation of glandular trichomes, which are pivotal sites for resin synthesis and cannabinoid production. Remarkably, higher trichome densities emerged on the abaxial surfaces of the bracts. This asymmetry in trichome distribution might be attributed to the varied environmental conditions affecting the two surfaces differently or could be a result of genetic predisposition influencing the trichome placement [
15]. Consistent with the results of the current study, Punja et al. [
15] documented increased trichome density on the abaxial surface of bracts. Since trichomes represent major sites of secondary metabolite synthesis and sequestration, this temporal increase in trichome density aligns with the general understanding that secondary metabolite levels peak around the time that plants are considered mature.
The observed evolution of trichome development from week 2 to week 8 unveiled a synchronized enhancement in the trichome stalk length and glandular head diameter. This co-occurring increase suggests that, as trichomes matured, they underwent structural changes that led to the elongation of stalks and the expansion of glandular heads. Importantly, this developmental progression is correlated with color changes within the glandular heads. The shift from a transparent (translucent) initial state to an opaque (milky white) appearance, and, eventually, to an amber (brown) shade, indicates the synthesis and accumulation of cannabinoids and other resinous compounds within the glandular trichomes. This alteration in resin color could signify the maturation and chemical transformation of secondary metabolites, including cannabinoids and terpenes, within the trichomes [
13,
15,
16]. These compounds play crucial roles in the biological and therapeutic properties of cannabis [
13].
5. Conclusions
The current study offers a detailed characterization of the entire life cycle of Cannabis sativa L. cv. White Widow, encompassing key aspects such as leaf morphology, plant structure, flower development, and trichome features. By studying the whole life cycle, a dynamic progression of the developmental cues and responses of the cannabis plant are revealed. The intricate journey of cannabis leaf development is highlighted by a gradual increase in morphological complexity, from the initiation of L1, one single serrated leaflet, to the peak of nine leaflets characteristic of L6. The leaflet number remained stable from nodes 6–13, which produced leaves with nine leaflets. However, a reduction in leaflet number was observed at node 14, which produced leaves with eight leaflets, and leaves with seven leaflets emerging from node 15. Towards nodes 20–24, the leaf complexity diminished, ultimately resulting in leaves of one single leaflet at node 25. The significant observation of leaf area peaking at L12, and the sharp decline in leaf area thereafter, which also coincides with a change in phyllotaxy, underscores the phase transition of cannabis. Under the long-day photoperiod, the plant’s transition from maturity traits to reproductive traits is marked by the emergence of bracts and solitary flowers. An intriguing shift in leaf arrangement from opposite to alternate was noted at node 12 while still under the long-day photoperiod. The transition to the short-day photoperiod triggered complex raceme-like inflorescence formation at the apical regions, indicative of monopodial growth.
The investigation into flower development revealed abundant glandular trichomes on perigonal bracts, with a dynamic shift in stigma color during maturation. The trichome development exhibited a concurrent growth in stalk length and glandular head diameter, which mirrored the resin’s color shift from transparent to opaque and, ultimately, amber. It is critical that future experiments focus on investigating the genetic and hormonal mechanisms involved in the intricate developmental processes described. In addition to guiding future experiments, the presented findings have significant implications for horticulturists and cultivators seeking to maximize plant health and optimize propagation and cultivation techniques.
The comprehensive developmental roadmap presented in the current study serves as a crucial tool for evaluating and harnessing morphological traits and should be extended to additional cannabis cultivars. By providing a detailed account of the plant’s ontological development, this study enables researchers and cultivators to make informed decisions relating to applicable methods and the timing of implementation. The information provided can inform strategies for plant rejuvenation, propagation, and cultivation, thereby contributing to optimal production outcomes. Moreover, this developmental roadmap is of paramount importance for developing an ontological transcriptomic reference for Cannabis sativa. A deeper understanding of the plant’s developmental stages and phase transitions is instrumental in deciphering the underlying genetic mechanisms governing juvenility and other critical traits, such as flowering. The occurrence of the unique solitary flower arrangement, which is separate from inflorescence in cannabis, suggests that the regulatory processes involved in flower induction may differ from other species. Consequently, this study contributes to the foundational knowledge required to unravel the intricacies of cannabis biology, particularly in the context of flowering, which will ultimately facilitate advancements in cultivation techniques.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}