

Salicylic Acid as a Salt Stress Mitigator on Chlorophyll Fluorescence, Photosynthetic Pigments, and Growth of Precocious-Dwarf Cashew in the Post-Grafting Phase

 ,
,  , ,
, , 
 ,
,
Abstract
1. Introduction
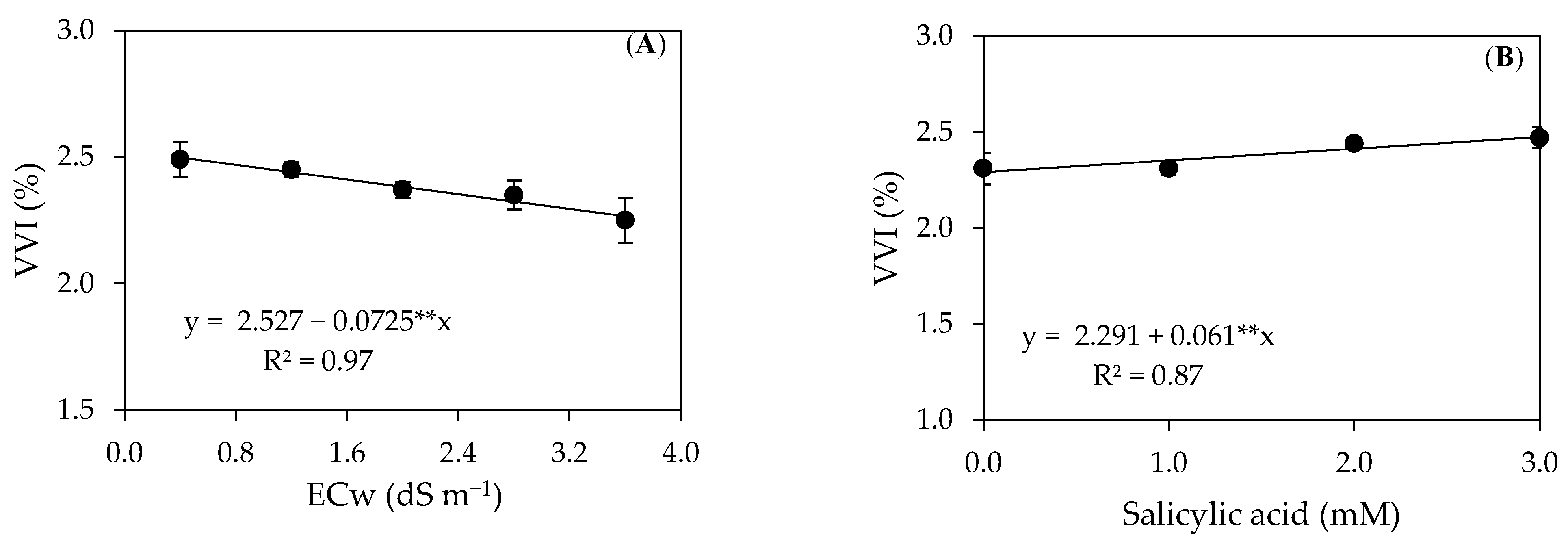
2. Results and Discussion
3. Materials and Methods
3.1. Location of the Experiment
3.2. Treatments and Experimental Design
3.3. Experiment Setup and Conduction
3.4. Traits Analyzed
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Souza, L.P.; Nobre, R.G.; Gheyi, H.R.; de Fatima, R.T.; de Silva, E.L.; Elias, J.J. Alocação de fitomassa e crescimento de clones de cajueiro irrigados com águas de diferentes salinidades. Rev. Interdiscip. UFT 2020, 7, 52–63. [Google Scholar] [CrossRef]
- Silva, A.F.; Sousa, F.d.O.; dos Santos, G.L.; Araújo Júnior, E.S.; da Silva, S.L.F.; de Macedo, C.E.C.; de Melo, A.S.; Maia, J.M. Antioxidant protection of photosynthesis in two cashew progenies under salt stress. J. Agric. Sci. 2018, 10, 388–404. [Google Scholar] [CrossRef]
- Farias, A.P.; Fonseca, N.C.P.; Oliveira, J.P.; Bastos, G.G. Caju: O sabor do Nordeste. Rev. Gastron. 2019, 1, 1–15. [Google Scholar]
- IBGE—Instituto Brasileiro de Geografia e Estatística. 2021; Disponível em. Available online: https://www.ibge.gov.br/explica/producao-agropecuaria/castanha-de-caju-cultivo/br (accessed on 12 November 2022).
- Pinheiro, F.W.A.; Lima, G.S.D.; Sousa, P.F.d.N.; de Souza, W.B.B.; Soares, L.A.d.A.; Gheyi, H.R.; Felix, C.M.; Paiva, F.J.d.S. Potassium fertilization in the cultivation of sour passion fruit under irrigation strategies with brackish water. Rev. Bras. Eng. Agrícola Ambient. 2023, 27, 42–50. [Google Scholar] [CrossRef]
- Lacerda, C.N.; Lima, G.S.D.; Soares, L.A.d.A.; de Fátima, R.T.; Gheyi, H.R.; de Azevedo, C.A.V. Morphophysiology and production of guava as a function of water salinity and salicylic acid. Rev. Bras. Eng. Agrícola Ambient. 2022, 26, 451–458. [Google Scholar] [CrossRef]
- Bonifácio, B.F.; Nobre, R.G.; Sousa, A.d.S.; Gomes, E.M.; da Silva, E.M.; Sousa, L.d.P. Efeitos da adubação potássica e irrigação com águas salinas no crescimento de porta-enxerto de goiabeira. Rev. Bras. Cienc. Agrar. 2018, 41, 101–110. [Google Scholar] [CrossRef]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef]
- Najar, R.; Aydi, S.; Sassi-Aydi, S.; Zarai, A.; Abdelly, C. Effect of salt stress on photosynthesis and chlorophyll fluorescence in Medicago truncatula. Plant. Biosyst. 2019, 153, 88–97. [Google Scholar] [CrossRef]
- Oliveira, F.I.F.; Souto, A.G.L.; Cavalcante, F.L.; Medeiros, W.J.F.; Medeiros, S.A.S.; Oliveira, F.F. Biomass and chloroplast pigments in jackfruit seedlings under saline stress and nitrogen fertilization. Rev. Caatinga 2018, 31, 662–663. [Google Scholar] [CrossRef]
- Sousa, V.F.O.; Santos, G.L.; Maia, J.M.; Maia Júnior, S.O.; Santos, J.P.O.; Costa, J.E.; Silva, A.F.; Dias, T.J.; Ferreira-Silva, S.L.; Taniguchi, C.A.K. Salinity-tolerant dwarf cashew tree rootstock has better ionic homeostasis and morphophysiological performance of seedlings. Rev. Bras. Eng. Agrícola Ambient. 2023, 27, 92–100. [Google Scholar] [CrossRef]
- Aldesuquy, H.S.; Ibraheem, F.L.; Ghanem, H.E. Exogenously supplied salicylic acid and trehalose protect growth vigor, chlorophylls and thylakoid membranes of wheat flag leaf from drought-induced damage. J. Agric. For. Meteorol. 2018, 1, 13–20. [Google Scholar]
- Wang, Y.Y.; Wang, Y.; Li, G.Z.; Hao, L. Arabidopsis plant response that alters salicylic acid to cadmium exposure: Underlying mechanisms affecting antioxidation and photosynthesis-related processes. Ecotoxicol. Environ. Saf. 2019, 69, 645–653. [Google Scholar] [CrossRef]
- da Silva, A.A.R.; de Lima, G.S.; de Azevedo, C.A.V.; Veloso, L.L.d.S.; Gheyi, H.R. Salicylic acid as anattenuator of salt stress in soursop. Rev. Caatinga 2020, 33, 1092–1101. [Google Scholar] [CrossRef]
- Poór, P.; Borbély, P.; Bódi, N.; Bagyánszki, M.; Tari, I. Effects of salicylic acid on photosynthetic activity and chloroplast morphology under light and prolonged darkness. Photosynthetica 2019, 57, 367–376. [Google Scholar] [CrossRef]
- Ekbic, H.B.; Ozcan, N.; Erdem, H. Impacts of salicylic acid treatments on salt resistance of some American grapevine rootstocks. Fresenius Environ. Bull. 2020, 29, 685–692. [Google Scholar]
- Samadi, S.; Habibi, G.; Vaziri, A. Effects of exogenous salicylic acid on antioxidative responses, phenolic metabolism and photochemical activity of strawberry under salt stress. J. Plant Physiol. 2019, 9, 2685–2694. [Google Scholar] [CrossRef]
- de Morais, M.B.; Camara, T.R.; Ulisses, C.; Carvalho Filho, J.L.S.; Willadino, L. Multiple stresses on the oxidative metabolism of sugarcane varieties. Ciênc. Rural. 2018, 48, 1–8. [Google Scholar] [CrossRef]
- da Silva, A.A.R.; de Lima, G.S.; de Azevedo, C.A.V.; Gheyi, H.R.; de Souza, A.R.; Fernandes, P.D. Salicylic acid relieves the effect of saline stress on soursop morphysiology. Rev. Ciênc. Agrotec. 2021, 33, e007021. [Google Scholar] [CrossRef]
- Batista, V.C.V.; Pereira, I.M.C.; Paulo-Marinho, S.d.O.; Canuto, K.M.; Pereira, R.d.C.A.; Rodrigues, T.H.S.; Daloso, D.d.M.; Gomes Filho, E.; de Carvalho, H.H. Salicylic acid modulates primary and volatile metabolites to alleviate salt stress-induced photosynthesis impairment on medicinal plant Egletes viscosa. Environ. Exp. Bot. 2019, 167, e103870. [Google Scholar] [CrossRef]
- Yudina, L.; Sukhova, E.; Gromova, E.; Nerush, V.; Vodeneev, V.; Sukhov, V. A light-induced decrease in the photochemical reflectance index (PRI) can be used to estimate the energy-dependent component of non-photochemical quenching under heat stress and soil drought in pea, wheat, and pumpkin. Photosynth. Res. 2020, 146, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, A.F.G.; Gomes, J.W.d.S.; Avilez, A.A.; Sarria, S.D.; Broetto, F.; Vieites, R.L.; Guimarães, M.L.C.S. Foliar salicylic acid application to mitigate the effect of water deficiency on potato (Solanum tuberosum L.). Plant Stress 2023, 7, e100135. [Google Scholar] [CrossRef]
- Sullivan, C.Y. Mechanisms of heat drought resistence in grain sorghum and methods of measurement. In Sorghum in Seventies; Rao, N.G.P., House, L.R., Eds.; Oxford and IBH Publication: New Delhi, India, 1971; 247p. [Google Scholar]
- Mostajeran, A.; Gholaminejad, A.; Asghari, G. Salinity alters curcumin, essential oil and chlorophyll of turmeric (Curcuma longa L.). Res. Pharm Sci. 2014, 9, 49–57. Available online: http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4292181/ (accessed on 15 January 2023).
- Taibi, K.; Taibi, F.; Abderrahim, L.A.; Ennajah, A.; Belkhodja, M.; Mulet, J.M. Effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidant defence systems in Phaseolus vulgaris L. S. Afr. J. Bot. 2016, 105, 306–312. [Google Scholar] [CrossRef]
- de Lima, G.S.; da Silva, J.B.; Soares, L.A.d.A.; Nobre, R.G.; Gheyi, H.R.; Diniz, G.L.; Fernandes, P.D.; Dantas, S.J.; de Lima, V.L.A. Effect of water salinity and potassium doses on physiological traits and growth of ‘Embrapa 51′ precocious dwarf cashew (Anacardium occidentale L.) rootstock. Aust. J. Crop Sci. 2020, 14, 1748–1755. [Google Scholar] [CrossRef]
- da Silva, A.R.A.; Bezerra, F.M.L.; de Lacerda, C.F.; de Sousa, C.H.C.; Chagas, K.L. Pigmentos fotossintéticos e potencial hídrico foliar em plantas jovens de coqueiro sob estresses hídrico e salino. Rev. Agro@Mbiente 2017, 10, 317–325. [Google Scholar] [CrossRef]
- Szepesi, Á. Influence of exogenous salicylic acid on antioxidant enzyme activities in the roots of salt stressed tomato plants. Acta Biol. Szeged. 2008, 52, 199–200. [Google Scholar]
- Aires, E.S.; Ferraz, A.K.L.; Carvalho, B.L.; Teixeira, F.P.; Rodrigues, J.D.; Ono, E.O. Foliar application of salicylic acid intensifies antioxidant system and photosynthetic efficiency in tomato plants. Bragantia 2022, 81, e1522. [Google Scholar] [CrossRef]
- da Silva, A.A.R.; de Lima, G.S.; de Azevedo, C.A.V.; Gheyi, H.R.; Soares, L.A.d.A.; Veloso, L.L.d.S.A. Salicylic acid improves physiological indicators of soursop irrigated with saline water. Rev. Bras. Eng. Agrícola Ambient. 2022, 26, 412–419. [Google Scholar] [CrossRef]
- da Silva, E.M.; Nobre, R.G.; Souza, L.d.P.; Pinheiro, F.W.A.; de Lima, G.S.; Gheyi, H.R.; Almeida, L.d.L.S. Physiology of Paluma guava under irrigation with saline water and nitrogen fertilization. Semin. Ciênc. Agrar. 2017, 38, 623–634. [Google Scholar] [CrossRef]
- Hu, Y.; Yue, J.; Nie, J.; Luo, D.; Cao, S.; Wang, C.; Chen, P. Salicylic acid alleviates the salt toxicity in kenaf by activating antioxidant system and regulating crucial pathways and genes. Ind. Crop. Prod. 2023, 199, e116691. [Google Scholar] [CrossRef]
- de Lima, G.S.; da Silva, J.B.; Souza, L.d.P.; Nobre, R.G.; Soares, L.A.d.A.; Gheyi, H.R. Tolerance of precocious dwarf cashew clones to salt stress during rootstock formation stage. Rev. Bras. Eng. Agrícola Ambient. 2020, 24, 474–481. [Google Scholar] [CrossRef]
- Dantas, M.V.; de Lima, G.S.; Gheyi, H.R.; da Silva, A.A.R.; de Melo, A.S.; de Medeiros, L.C. Gas exchange and photosynthetic pigments of West Indian cherry under salinity stress and salicylic acid. Comun. Sci. 2021, 12, e3664. [Google Scholar]
- Martins, J.B.; Santos Júnior, J.A.; Bartusch, V.P.; Gheyi, H.R.; Bezerra Neto, E.; Silva, M.M.D. Water relations in parsley plants cultivated in brackish nutrient solutions of different cationic natures. Rev. Bras. Eng. Agrícola Ambient. 2019, 23, 662–668. [Google Scholar] [CrossRef]
- Lotfi, R.; Ghassemi-Golezani, K.; Pessarakli, M. Salicylic acid regulates photosynthetic electron transfer and stomatal conductance of mung bean (Vigna radiata L.) under salinity stress. Biocatal. Agric. Biotechnol. 2020, 26, e101635. [Google Scholar] [CrossRef]
- de Lima, G.S.; de Neto, V.E.S.; Gheyi, H.R.; Nobre, R.G.; Diniz, G.L.; Soares, L.A.d.A.; Pinheiro, F.W.A. Photosynthetic pigments and photochemical efficiency of precocious dwarf cashew (Anacardium occidentale L.) under salt stress and potassium fertilization. AIMS Agric. Food. 2019, 4, 1007–1019. [Google Scholar] [CrossRef]
- Saddiq, M.S.; Iqbal, S.; Hafeez, M.B.; Ibrahim, A.M.; Raza, A.; Fatima, E.M.; Ciarmiello, L.F. Effect of salinity stress on physiological changes in winter and spring wheat. Agronomy 2021, 11, 1193. [Google Scholar] [CrossRef]
- Fernandes, E.A.; Soares, L.A.d.A.; de Lima, G.S.; Gheyi, H.R.; Nobre, R.G.; Fernandes, P.D. Photosynthetic pigments, photochemical efficiency and growth of custard-apple under salt stress and potassium fertilization. Rev. Bras. Eng. Agrícola Ambient. 2022, 26, 365–373. [Google Scholar] [CrossRef]
- Diniz, G.L.; Nobre, R.G.; de Lima, G.S.; Soares, L.A.d.A.; Gheyi, H.R. Irrigation with saline water and silicate fertilization in the cultivation of ‘Gigante Amarelo’ passion fruit. Rev. Caatinga 2021, 34, 199–207. [Google Scholar] [CrossRef]
- Xavier, A.V.; de Lima, G.S.; Gheyi, H.R.; da Silva, A.A.R.; de Lacerda, C.N.; Soares, L.A.d.A.; Fernandes, P.D. Salicylic acid alleviates salt stress on guava plant physiology during rootstock formation. Rev. Bras. Eng. Agrícola Ambient. 2022, 26, 855–862. [Google Scholar] [CrossRef]
- Boukraâ, D.; Benabdelli, K.; Belabid, L.; Bennabi, F. Effect of salinity on chickpea seed germination pre-treated with salicylic acid. Sci. J. Biol. Sci. 2013, 2, 86–93. [Google Scholar]
- Ferreira, J.F.; da Silva Filho, J.B.; Liu, X.; Sandhu, D. Spinach plants favor the absorption of K+ over Na+ regardless of salinity, and may benefit from Na+ when K+ is deficient in the soil. Plants 2020, 9, 507. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.; Iqbal, M.; Aslam, A.; Nawaz, A.; Khan, M.B. Foliar spray of salicylic acid enhances growth, yield and biochemical attributes of tomato (Solanum lycopersicum L.) seedlings. Plant Growth Regul. 2018, 84, 147–158. [Google Scholar]
- Zvanarou, S.; Vágnerová, R.; Mackievic, V.; Usnich, S.; Smolich, I.; Sokolik, A.; Demidchik, V. Salt stress triggers generation of oxygen free radicals and DNA breaks in Physcomitrella patens, protonema. Environ. Exp. Bot. 2020, 180, e104236. [Google Scholar] [CrossRef]
- Zahid, A.; Razzaq, A.A.; Munawar, M.; Ramzan, M.; Almutairi, B.O.; Almutairi, M.H. Foliar spray of salicylic acid and ascorbic acid ameliorates the biochemical compounds in hybrid chillies. J. King Saud Univ. Sci. 2023, 35, e102660. [Google Scholar] [CrossRef]
- Galvão Sobrinho, T.; da Silva, A.A.R.; de Lima, G.S.; de Lima, V.L.A.; Borges, V.E.; Nunes, K.G.; Azevedo, C.A.V. Foliar applications of salicylic acid on boosting salt stress tolerance in sour passion fruit in two cropping cycles. Plants 2023, 12, 2023. [Google Scholar] [CrossRef]
- Souza, A.R.; da Silva, A.A.R.; de Lima, G.S.; de Azevedo, C.A.V.; Soares, L.A.d.A.; Silva, F.d.A.d.; Costa, D.S. Morphophysiology and production of bell pepper grown under salt stress and salicylic acid foliar application. Braz. J. Biol. 2023, 83, e273017. [Google Scholar] [CrossRef]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonçalves, J.L.d.M.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- Cavalcante Júnior, L.F. Eficiências de Absorção, Transporte e Utilização de Nutrientes de Mudas de Cajueiro-anão-Precoce. Dissertação de Mestrado, (Mestrado em Agronomia/Solos e Nutrição de Plantas). Centro de Ciências Agrárias, Universidade Federal do Ceará, Fortaleza, Brazil, 2013; 65p. [Google Scholar]
- Teixeira, P.C.; Donagemma, G.K.; Fontana, A.; Teixeira, W.G. Manual de Métodos de Análise de Solo, 3rd ed.; Embrapa Solos: Brasília, Brazil, 2017; 574p. [Google Scholar]
- Medeiros, J.F.; Lisboa, R.A.; de Oliveira, M.; de Silva Júnior, M.J.; Alves, L.P. Caracterização das águas subterrâneas usadas para irrigação na área produtora de melão da Chapada do Apodi. Rev. Bras. Eng. Agricola Ambient. 2003, 7, 469–472. [Google Scholar] [CrossRef]
- Richards, L.A. Diagnosis and Improvement of Saline and Alkali Soils; Department of Agriculture: Washington, DC, USA, 1954; 160p. [Google Scholar]
- Oliveira, V.H. Cultivo do Cajueiro Anão Precoce; Embrapa Agroindústria Tropical: Fortaleza, Brazil, 2002; 44p. [Google Scholar]
- Weatherley, P.E. Studies in the water relations of the cotton plant. I. The field measurement of water deficits in leaves. New Phytol. 1950, 49, 81–97. [Google Scholar] [CrossRef]
- Scotti-Campos, P.; Pham-Thi, A.T.; Semedo, J.N.; Pais, I.P.; Ramalho, J.C.; Matos, M.C. Physiological responses and membrane integrity in three Vigna genotypes with contrasting drought tolerance. Emir. J. Food Agric. 2013, 25, 1002–1013. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts: Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Portella, C.R.; Marinho, C.S.; Amaral, B.D.; Carvalho, W.S.G.; Campos, G.S.; de Silva, M.P.S.; de Sousa, M.C. Desempenho de cultivares de citros enxertadas sobre o trifoliateiro ‘Flying Dragon’ e limoeiro ‘cravo’ em fase de formação do pomar. Bragantia 2016, 75, 70–75. [Google Scholar] [CrossRef][Green Version]
- Benincasa, M.M.P. Análise de Crescimento de Plantas, Noções Básicas, 2nd ed.; FUNEP: Jaboticabal, Brazil, 2003; 41p. [Google Scholar]
- Ferreira, D.F. SISVAR: A computer analysis system to fixed effects split plot type designs. Rev. Bras. Biom. 2019, 37, 529–535. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | DF | Mean Squares | |
|---|---|---|---|
| RWC | % EL | ||
| Salinity (S) | 4 | 560.97 ** | 495.86 ** |
| Linear regression | 1 | 1843.26 ** | 1641.91 ** |
| Quadratic regression | 1 | 364.88 ** | 329.44 ** |
| Salicylic acid (SA) | 3 | 109.17 ** | 110.16 ** |
| Linear regression | 1 | 131.59 ** | 191.02 ** |
| Quadratic regression | 1 | 159.12 ** | 44.29 ** |
| Interaction (S × SA) | 12 | 14.01 ** | 7.59 * |
| Blocks | 8 | 10.54 ns | 6.73 ns |
| Residual | 3 | 3.81 | 3.67 |
| CV (%) | 2.70 | 8.12 | |
| Source of Variation | DF | Mean Squares | |||
|---|---|---|---|---|---|
| Chl a | Chl b | Car | Chl Total | ||
| Salinity (S) | 4 | 4779.97 ** | 2468.87 * | 796.35 ** | 13,243.18 ** |
| Linear regression | 1 | 14,828.74 ** | 9562.95 ** | 3099.47 ** | 48,208.62 ** |
| Quadratic regression | 1 | 4134.65 * | 1.2411 ns | 29.48 * | 3993.30 ns |
| Salicylic acid (SA) | 3 | 6849.98 ** | 3055.15 ** | 115.81 ns | 1556.24 ns |
| Linear regression | 1 | 2552.66 ns | 3771.73 ** | 62.62 ns | 118.64 ns |
| Quadratic regression | 1 | 140,487.66 ** | 2715.82 ** | 272.85 ns | 4433.81 ns |
| Interaction (S × SA) | 12 | 1097.39 ns | 498.76 ns | 147.17 ns | 1977.88 ns |
| Blocks | 8 | 2029.53 ns | 865.03 ns | 198.14 ns | 5377.52 ns |
| Residual | 3 | 918.14 | 289.94 | 91.70 | 1523.33 |
| CV (%) | 17.38 | 22.73 | 13.42 | 15.70 | |
| Mean Squares | |||||
|---|---|---|---|---|---|
| Source of Variation | DF | F0 | Fm | Fv | Fv/Fm |
| Salinity (S) | 4 | 16,496.041 * | 136,642.79 ** | 262,471.097 ** | 0.04619 ** |
| Linear regression | 1 | 5526.20 ** | 521,027.81 ** | 1,043,077.75 ** | 0.177 ** |
| Quadratic regression | 1 | 7668.00 ns | 12,325.89 ns | 627.21 ns | 0.00526 ns |
| Salicylic acid (SA) | 3 | 2189.75 ns | 3753.62 ns | 5578.44 ns | 0.002156 ns |
| Linear regression | 1 | 2852.08 ns | 4004.71 ns | 15,513.28 ns | 0.00563 ns |
| Quadratic regression | 1 | 350.41 ns | 245.19 ns | 1075.69 ns | 0.001 ns |
| Interaction (S × SA) | 12 | 6749.22 ns | 1564.98 ns | 9407.10 ns | 0.00668 ns |
| Blocks | 8 | 5273.45 ns | 34,893.38 ns | 49,964.031 ns | 0.00862 ns |
| Residual | 3 | 6180.82 | 9491.58 | 1035.32 | 0.005102 |
| CV (%) | 20.86 | 8.31 | 12.92 | 9.94 | |
| Source of Variation | DF | Mean Squares | ||||||
|---|---|---|---|---|---|---|---|---|
| SDrootstock | SDscion | SDgrafting point | PH | VCrown | DCrown | VVI | ||
| Salinity (S) | 4 | 41.62 ** | 8.10 ** | 17.69 ** | 101.4 ns | 0.00252 ns | 0.00475 ns | 0.107 ** |
| Linear regression | 1 | 152.46 ** | 30.9 ** | 56.73 ** | 74.30 ns | 0.00917 ns | 0.00179 ns | 0.414 ** |
| Quadratic regression | 1 | 4.82 ** | 0.0572 ns | 13.57 ** | 0.43 ns | 0.000213 ns | 0.00047 ns | 0.00095 ns |
| Salicylic acid (SA) | 3 | 20.85 ** | 3.15 ns | 6.35 ns | 326.55** | 0.00193 ns | 0.00318 ns | 0.114 ** |
| Linear regression | 1 | 61.32 ** | 7.76 ns | 0.139 ns | 703.52 ** | 0.0011 ns | 0.00042 ns | 0.293 ** |
| Quadratic regression | 1 | 1.14 ns | 1.4 ns | 0.2018 ns | 44.63 ns | 0.00016 ns | 0.00876 ns | 0.0026 ** |
| Interaction (S × SA) | 12 | 8.43 ** | 7.7 ** | 19.01 ns | 128.07 ns | 0.00851 ns | 0.0127 ns | 0.102 ns |
| Blocks | 8 | 0.34 ns | 0.85 ns | 0.011 ns | 38.66 ns | 0.00993 ns | 0.00913 ns | 0.0201 ns |
| Residual | 3 | 1.39 | 1.54 | 1.87 | 12.88 | 0.00193 | 0.00913 | 0.0197 |
| CV (%) | 4.40 | 8.14 | 5.81 | 4.20 | 19.28 | 6.87 | 5.89 | |
| Source of Variation | DF | Mean Squares | |
|---|---|---|---|
| RGBSDrootstock | RGBSDscion | ||
| Salinity (S) | 4 | 9.0 × 10−6 ** | 2.0 × 10−6 ** |
| Linear regression | 1 | 3.0 × 10−5 ** | 3.0 × 10−6 ** |
| Quadratic regression | 1 | 4.0 × 10−6 ns | 2.0 × 10−6 ** |
| Salicylic acid (SA) | 3 | 5.0 × 10−6 ** | 4.0 × 10−6 ** |
| Linear regression | 1 | 1.1 × 10−5 * | 1.0 × 10−8 ns |
| Quadratic regression | 1 | 2.0 × 10−6 ** | 1.1 × 10−6 ** |
| Interaction (S × SA) | 12 | 3.0 × 10−6 ** | 2.0 × 10−6 ** |
| Blocks | 8 | 2.0 × 10−6 ns | 2.5 × 10−7 ns |
| Residual | 3 | 3.69 × 10−7 | 3.8 × 10−7 |
| CV (%) | 15.27 | 18.22 | |
| ECw (S) | Salicylic Acid (SA) Concentrations | |||
|---|---|---|---|---|
| 0 mM | 1 mM | 2 mM | 3 mM | |
| 0.4 dS m−1 | S1SA1 (control) | S1SA2 | S1SA3 | S1SA4 |
| 1.2 dS m−1 | S2SA1 | S2SA2 | S2SA3 | S2SA4 |
| 2.0 dS m−1 | S3SA1 | S3SA2 | S3SA3 | S3SA4 |
| 2.8 dS m−1 | S4SA1 | S4SA2 | S4SA3 | S4SA4 |
| 3.6 dS m−1 | S5SA1 | S5SA2 | S5SA3 | S5SA4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arruda, T.F.d.L.; Lima, G.S.d.; Silva, A.A.R.d.; Azevedo, C.A.V.d.; Souza, A.R.d.; Soares, L.A.d.A.; Gheyi, H.R.; Lima, V.L.A.d.; Fernandes, P.D.; Silva, F.d.A.d.; et al. Salicylic Acid as a Salt Stress Mitigator on Chlorophyll Fluorescence, Photosynthetic Pigments, and Growth of Precocious-Dwarf Cashew in the Post-Grafting Phase. Plants 2023, 12, 2783. https://doi.org/10.3390/plants12152783
Arruda TFdL, Lima GSd, Silva AARd, Azevedo CAVd, Souza ARd, Soares LAdA, Gheyi HR, Lima VLAd, Fernandes PD, Silva FdAd, et al. Salicylic Acid as a Salt Stress Mitigator on Chlorophyll Fluorescence, Photosynthetic Pigments, and Growth of Precocious-Dwarf Cashew in the Post-Grafting Phase. Plants. 2023; 12(15):2783. https://doi.org/10.3390/plants12152783
Chicago/Turabian StyleArruda, Thiago Filipe de Lima, Geovani Soares de Lima, André Alisson Rodrigues da Silva, Carlos Alberto Vieira de Azevedo, Allesson Ramos de Souza, Lauriane Almeida dos Anjos Soares, Hans Raj Gheyi, Vera Lúcia Antunes de Lima, Pedro Dantas Fernandes, Francisco de Assis da Silva, and et al. 2023. "Salicylic Acid as a Salt Stress Mitigator on Chlorophyll Fluorescence, Photosynthetic Pigments, and Growth of Precocious-Dwarf Cashew in the Post-Grafting Phase" Plants 12, no. 15: 2783. https://doi.org/10.3390/plants12152783
APA StyleArruda, T. F. d. L., Lima, G. S. d., Silva, A. A. R. d., Azevedo, C. A. V. d., Souza, A. R. d., Soares, L. A. d. A., Gheyi, H. R., Lima, V. L. A. d., Fernandes, P. D., Silva, F. d. A. d., Dias, M. d. S., Chaves, L. H. G., & Saboya, L. M. F. (2023). Salicylic Acid as a Salt Stress Mitigator on Chlorophyll Fluorescence, Photosynthetic Pigments, and Growth of Precocious-Dwarf Cashew in the Post-Grafting Phase. Plants, 12(15), 2783. https://doi.org/10.3390/plants12152783