
Current Status of Biodiversity Assessment and Conservation of Wild Olive (Olea europaea L. subsp. europaea var. sylvestris)

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Morphological and Genetic Characterization of Oleaster
2.1. Morphological Evaluation
2.2. Genetic Diversity Assessment
2.3. Correlation between the Oleaster Population Genetic Differentiation and the Country of Origin
3. Conservation of Oleaster Genetic Resources
4. Conclusions and Future Perspective
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carrión, Y.; Ntinou, M.; Badal, E. Olea europaea L. in the North Mediterranean Basin during the Pleniglacial and the Early–Middle Holocene. Quat. Sci. Rev. 2010, 29, 952–968. [Google Scholar] [CrossRef] [Green Version]
- Green, P.S. A revision of Olea L. (Oleaceae). Kew Bull. 2002, 57, 91–140. [Google Scholar] [CrossRef]
- Kassa, A.; Konrad, H.; Geburek, T. Molecular diversity and gene flow within and among different subspecies of the wild olive (Olea europaea L.): A review. Flora 2019, 250, 18–26. [Google Scholar] [CrossRef]
- Besnard, G.; Khadari, B.; Navascues, M.; Fernandez-Mazuecos, M.; El Bakkali, A.; Arrigo, N.; Baali-Cherif, D.; Brunini-Bronzini de Caraffa, V.; Santoni, S.; Vargas, P.; et al. The complex history of the olive tree: From Late Quaternary diversification of Mediterranean lineages to primary domestication in the northern Levant. Proc. R. Soc. B. 2013, 280, 20122833. [Google Scholar] [CrossRef] [Green Version]
- Diez, C.M.; Trujillo, I.; Martinez-Uriroz, N.; Barranco, D.; Rallo, L. Olive domestication and diversification in the Mediterranean Basin. New Phytol. 2015, 206, 436–447. [Google Scholar] [CrossRef]
- Arenas-Castro, S.; Gonçalves, J.F.; Moreno, M.; Villar, R. Projected climate changes are expected to decrease the suitability and production of olive varieties in southern Spain. Sci. Total Environ. 2020, 709, 136161. [Google Scholar] [CrossRef]
- International Olive Council (IOC). Available online: https://www.internationaloliveoil.org (accessed on 10 December 2021).
- Gianguzzi, L.; Bazan, G. The Olea europaea L. var. sylvestris (Mill.) Lehr. forests in the Mediterranean area. Plant Sociol. 2019, 56, 3–34. [Google Scholar]
- Belaj, A.; Gurbuz Veral, M.; Sikaoui, H.; Moukhli, A.; Khadari, B.; Mariotti, R.; Baldoni, L. Olive Genetic Resources. In The Olive Tree Genome, Compendium of Plant Genomes; Rugini, E., Baldoni, L., Muleo, R., Sebastiani, L., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 27–54. [Google Scholar]
- Turrill, W.B. Wild and cultivated olives. Kew Bull. 1951, 6, 437–442. [Google Scholar] [CrossRef]
- Besnard, G.; Bervillé, A. Multiple origins for Mediterranean olive (Olea europaea L. ssp. europaea) based upon mitochondrial DNA polymorphisms. C. R. Acad. Sci. Ser. III Sci. Vie. 2000, 323, 173–181. [Google Scholar] [CrossRef]
- Bronzini de Caraffa, V.; Maury, J.; Gambotti, C.; Breton, C.; Bervillé, A.; Giannettini, J. Mitochondrial DNA variation and RAPD mark oleasters, olive and feral olive from Western and Eastern Mediterranean. Theor. Appl. Genet. 2002, 104, 1209–1216. [Google Scholar] [CrossRef]
- Chiappetta, A.; Muto, A.; Muzzalupo, R.; Muzzalupo, I. New rapid procedure for genetic characterization of Italian wild olive (Olea europaea) and traceability of virgin olive oils by means of SSR markers. Sci. Hortic. 2017, 226, 42–49. [Google Scholar] [CrossRef]
- Falek, W.; Sion, S.; Montemurro, C.; Mascio, I.; Gadaleta, S.; Fanelli, V.; Savoia, M.A.; Piarulli, L.; Bechkri, S.; Khelifi, D.; et al. Molecular diversity and ecogeographic distribution of Algerian wild olives (Olea europaea subsp. europaea var. sylvestris). Sci. Agric. 2022, 79, e20200308. [Google Scholar] [CrossRef]
- Gualdi, S.; Somot, S.; Li, L.; Artale, V.; Adani, M.; Bellucci, A.; Braun, A.; Calmanti, S.; Carillo, A.; Dell’Aquila, A.; et al. The CIRCE simulations: Regional climate change projections with realistic representation of the Mediterranean Sea. Bull. Am. Meteorol. Soc. 2013, 94, 65–81. [Google Scholar] [CrossRef]
- Carlucci, A.; Raimondo, M.L.; Cibelli, F.; Phillips, A.J.L.; Lops, F. Pleurostomophora richardsiae, Neofusicoccum parvum and Phaeoacremonium aleophilum associated with a decline of olives in southern Italy. Phytopathol. Mediterr. 2013, 52, 517–527. [Google Scholar]
- White, S.M.; Navas-Cortés, J.A.; Bullock, J.M.; Boscia, D.; Chapman, D.S. Estimating the epidemiology of emerging Xylella fastidiosa outbreaks in olives. Plant Pathol. 2020, 69, 1403–1413. [Google Scholar] [CrossRef]
- Colella, C.; Miacola, C.; Amenduni, M.; D’Amico, M.; Bubici, G.; Cirulli, M. Sources of verticillium wilt resistance in wild olive germplasm from the Mediterranean region. Plant Pathol. 2008, 57, 533–539. [Google Scholar] [CrossRef]
- Díaz-Rueda, P.; Aguado, A.; Romero-Cuadrado, L.; Capote, N.; Colmenero-Flores, J.M. Wild olive genotypes as a valuable source of resistance to defoliating Verticillium dahliae. Front. Plant Sci. 2021, 12, 662060. [Google Scholar] [CrossRef]
- Hernández-Santana, V.; Diaz-Rueda, P.; Diaz-Espejo, A.; Raya-Sereno, M.D.; Gutierrez-Gordillo, S.; Montero, A.; Perez-Martin, A.; Colmenero-Flores, J.M.; Rodriguez-Dominguez, C.M. Hydraulic traits emerge as relevant determinants of growth patterns in wild olive genotypes under water stress. Front. Plant Sci. 2019, 10, 291. [Google Scholar] [CrossRef]
- Palomares-Rius, J.E.; Belaj, A.; León, L.; de la Rosa, R.; Rapoport, H.F.; Castillo, P. Evaluation of the phytopathological reaction of wild and cultivated olives as a means of finding promising new sources of genetic diversity for resistance to root-knot nematodes. Plant Dis. 2019, 103, 2559–2568. [Google Scholar] [CrossRef] [Green Version]
- León, L.; Díaz-Rueda, P.; Belaj, A.; De la Rosa, R.; Carrascosa, C.; Colmenero-Flores, J.M. Evaluation of early vigor traits in wild olive germplasm. Sci. Horticult. 2020, 264, 109–157. [Google Scholar] [CrossRef] [Green Version]
- Sion, S.; Taranto, F.; Montemurro, C.; Mangini, G.; Camposeo, S.; Falco, V.; Gallo, A.; Mita, G.; Saddoud Debbabi, O.; Ben Amar, F.; et al. Genetic characterization of Apulian olive germplasm as potential source in new breeding programs. Plants 2019, 8, 268. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Rueda, P.; Franco-Navarro, J.; Messora, R.; Espartero, J.; Rivero-Núñez, C.; Aleza, P.; Capote, N.; Cantos, M.; García-Fernández, J.L.; de Cires, A.; et al. SILVOLIVE, a germplasm collection of wild subspecies with high genetic variability as a source of rootstocks and resistance genes for olive breeding. Front. Plant Sci. 2020, 11, 629. [Google Scholar] [CrossRef]
- Pavan, S.; Vergine, M.; Nicolì, F.; Sabella, E.; Aprile, A.; Negro, C.; Fanelli, V.; Savoia, M.A.; Montilon, V.; Susca, L.; et al. Screening of olive biodiversity defines genotypes potentially resistant to Xylella fastidiosa. Front. Plant Sci. 2021, 12, 723879. [Google Scholar] [CrossRef]
- Sebastiani, L.; Busconi, M. Recent developments in olive (Olea europaea L.) genetics and genomics: Applications in taxonomy, varietal identification, traceability and breeding. Plant Cell Rep. 2017, 36, 1345–1360. [Google Scholar] [CrossRef]
- León, L.; de la Rosa, R.; Velasco, L.; Belaj, A. Using wild olives in breeding programs: Implications on oil quality composition. Front. Plant Sci. 2018, 9, 232. [Google Scholar] [CrossRef]
- Espínola, F.; Vidal, A.M.; Espínola, J.M.; Moya, M. Processing effect and characterization of olive oils from Spanish wild olive trees (Olea europaea var. sylvestris). Molecules 2021, 26, 1304. [Google Scholar] [CrossRef]
- Amane, M.; Ouazzani, N.; Lumaret, R.; Debain, C. Chloroplast-DNA variation in the wild and cultivated olives (Olea europaea L.) of Morocco. Euphytica 2000, 116, 59–64. [Google Scholar] [CrossRef]
- Lumaret, R.; Amane, M.; Ouazzani, N.; Baldoni, L.; Debain, C. Chloroplast DNA variation in the cultivated and wild olive taxa of the genus Olea L. Theor. Appl. Genet. 2000, 101, 547–553. [Google Scholar] [CrossRef]
- Besnard, G.; Baradat, P.H.; Chevalier, D.; Tagmount, A.; Berville, A. Genetic differentiation in the olive complex (Olea europaea) revealed by RAPDs and RFLPs in the rRNA genes. Genet. Resour. Crop Evol. 2001, 48, 165–182. [Google Scholar] [CrossRef]
- Vargas, P.; Kadereit, J.W. Molecular fingerprinting evidence (ISSR, Inter-Simple Sequence Repeats) for a wild status of Olea europaea L. (Oleaceae) in the Eurosiberian North of the Iberian Peninsula. Flora 2001, 196, 142–152. [Google Scholar] [CrossRef]
- Besnard, G.; Bervillé, A. On chloroplast DNA variations in the olive (Olea europaea L.) complex: Comparison of RFLP and PCR polymorphisms. Theor. Appl. Genet. 2002, 104, 1157–1163. [Google Scholar] [CrossRef]
- Baldoni, L.; Tosti, N.; Ricciolini, C.; Belaj, A.; Arcioni, S.; Pannelli, G.; Germana, M.A.; Mulas, M.; Porceddu, A. Genetic structure of wild and cultivated olives in the Central Mediterranean Basin. Ann. Bot. 2006, 98, 935–942. [Google Scholar] [CrossRef]
- Breton, C.; Tersac, M.; Bervillé, A. Genetic diversity and gene flow between the wild olive (oleaster, Olea europaea L.) and the olive: Several Plio-Pleistocene refuge zones in the Mediterranean basin suggested by simple sequence repeats analysis. J. Biogeogr. 2006, 33, 1916–1928. [Google Scholar] [CrossRef]
- Belaj, A.; Muñoz-Diez, C.; Baldoni, L.; Porceddu, A.; Barranco, D.; Satovic, Z. Genetic diversity and population structure of wild olives from the north-western Mediterranean assessed by SSR markers. Ann. Bot. 2007, 100, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Brito, G.; Loureiro, J.; Lopes, T.; Rodriguez, E.; Santos, C. Genetic characterisation of olive trees from Madeira Archipelago using flow cytometry and microsatellite markers. Genet. Resour. Crop Evol. 2008, 55, 657–664. [Google Scholar] [CrossRef]
- Hannachi, H.; Breton, C.; Msallem, M.; El Hadj, S.B.; El Gazzah, M.; Berville, A. Differences between native and introduced olive cultivars as revealed by morphology of drupes, oil composition and SSR polymorphisms: A case study in Tunisia. Sci. Horticult. 2008, 116, 280–290. [Google Scholar] [CrossRef]
- Hannachi, H.; Sommerlatte, H.; Breton, C.; Msallem, M.; El Gazzah, M.; El Hadj, S.B.; Berville, A. Oleaster (var. sylvestris) and subsp. cuspidata are suitable genetic resources for improvement of the olive (Olea europaea subsp. europaea var. europaea). Genet. Resour. Crop Evol. 2009, 56, 393–403. [Google Scholar] [CrossRef]
- Belaj, A.; Munoz-Diez, C.; Baldoni, L.; Satovic, Z.; Barranco, D. Genetic diversity and relationships of wild and cultivated olives at regional level in Spain. Sci. Hortic. 2010, 124, 323–330. [Google Scholar] [CrossRef]
- Erre, P.; Chessa, I.; Munoz-Diez, C.; Belaj, A.; Rallo, L.; Trujillo, I. Genetic diversity and relationships between wild and cultivated olives (Olea europaea L.) in Sardinia as assessed by SSR markers. Genet. Resour. Crop Evol. 2010, 57, 41–54. [Google Scholar] [CrossRef]
- Hannachi, H.; Breton, C.; Msallem, M.; Sommerlatte, H.; El Hadj, S.B.; El Gazzah, M.; Berville, A. Genetic relationships between cultivated and wild olive trees (Olea europaea L. var. europaea and var. sylvestris) based on nuclear and chloroplast SSR markers. Nat. Resour. 2010, 1, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Sesli, M.; Yegenoglu, E.D. Comparison of Manzanilla and wild type olives by RAPD-PCR analysis. Afr. J. Biotechnol. 2010, 9, 986–990. [Google Scholar]
- Belaj, A.; León, L.; Satovic, Z.; de la Rosa, R. Variability of wild olives (Olea europaea subsp. europaea var. sylvestris) analyzed by agro-morphological traits and SSR markers. Sci. Horticult. 2011, 129, 561–569. [Google Scholar] [CrossRef]
- Ismaili, H.; Cantini, C.; Ianni, G.; Lloshi, I. Exploration and selection of the wild olive genotypes. J. Int. Environ. Appl. Sci. 2012, 7, 841–846. [Google Scholar]
- Besnard, G.; El Bakkali, A.; Haouane, H.; Baali-Cherif, D.; Moukhli, A.; Khadari, B. Population genetics of Mediterranean and Saharan olives: Geographic patterns of differentiation and evidence for early generations of admixture. Ann. Bot. 2013, 112, 1293–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beghè, D.; Piotti, A.; Satovic, Z.; de la Rosa, R.; Belaj, A. Pollen-mediated gene flow and fine-scale spatial genetic structure in Olea europaea subsp. europaea var. sylvestris. Ann. Bot. 2017, 119, 671–679. [Google Scholar]
- Boucheffa, S.; Miazzi, M.M.; di Rienzo, V.; Mangini, G.; Fanelli, V.; Tamendjari, A.; Pignone, D.; Montemurro, C. The coexistence of oleaster and traditional varieties affects genetic diversity and population structure in Algerian olive (Olea europaea) germplasm. Genet. Resour. Crop Evol. 2017, 64, 379–390. [Google Scholar] [CrossRef]
- Belaj, A.; De La Rosa, R.; Lorite, I.J.; Mariotti, R.; Cultrera, N.G.; Beuzón, C.R.; González-Plaza, J.J.; Muñoz-Mérida, A.; Trelles, O.; Baldoni, L. Usefulness of a new large set of high throughput EST-SNP markers as a tool for olive germplasm collection management. Front. Plant Sci. 2018, 9, 1320. [Google Scholar] [CrossRef] [Green Version]
- Di Rienzo, V.; Sion, S.; Taranto, F.; D’Agostino, N.; Montemurro, C.; Fanelli, V.; Sabetta, W.; Boucheffa, S.; Tamendjari, A.; Pasqualone, A.; et al. Genetic flow among olive populations within the Mediterranean basin. PeerJ 2018, 6, e5260. [Google Scholar] [CrossRef] [Green Version]
- Ismaili, H.; Veizi, A.; Lani, V. Collection and assessment of olive biodiversity. Int. J. Curr. Microbiol. App. Sci. 2018, 7, 1716–1726. [Google Scholar] [CrossRef]
- Boucheffa, S.; Tamendjari, A.; Sanchez-Gimeno, A.C.; Rovellini, P.; Venturini, S.; di Rienzo, V.; Miazzi, M.M.; Montemurro, C. Diversity assessment of Algerian wild and cultivated olives (Olea europaea L.) by molecular, morphological, and chemical traits. Eur. J. Lipid Sci. Technol. 2019, 121, 1800302. [Google Scholar] [CrossRef] [Green Version]
- Gros-Balthazard, M.; Besnard, G.; Sarah, G.; Holtz, Y.; Leclercq, J.; Santoni, S.; Wegmann, D.; Glémin, S.; Khadari, B. Evolutionary transcriptomics reveals the origins of olives and the genomic changes associated with their domestication. Plant J. 2019, 100, 143–157. [Google Scholar] [CrossRef]
- Mariotti, R.; Belaj, A.; De La Rosa, R.; Leòn, L.; Brizioli, F.; Baldoni, L.; Mousavi, S. EST–SNP study of Olea europaea L. uncovers functional polymorphisms between cultivated and wild olives. Genes 2020, 11, 916. [Google Scholar] [CrossRef]
- Rodrigues, N.; Pinho, T.; Casal, S.; Peres, A.M.; Baptista, P.; Pereira, J.A. Chemical characterization of Oleaster, Olea europaea var. sylvestris (Mill.) Lehr., oils from different locations of Northeast Portugal. Appl. Sci. 2020, 10, 6414. [Google Scholar] [CrossRef]
- Dervishi, A.; Jakše, J.; Ismaili, H.; Javornik, B.; Štajner, N. Genetic structure and core collection of olive germplasm from Albania revealed by microsatellite markers. Genes 2021, 12, 256. [Google Scholar] [CrossRef]
- Khouatmiani, K.; Belhadj, S.; Tonetto, A.; Assie, A.; Mevy, J.P.; Gauquelin, T. Variability of eight Algerian oleaster ecotypes (Olea europaea subsp. europaea var. sylvestris [Mill.] Lehr): Pollen and exine morphology in relation to geo-climatic effect. Grana 2021, 60, 256–270. [Google Scholar] [CrossRef]
- Sion, S.; Savoia, M.A.; Gadaleta, S.; Piarulli, L.; Mascio, I.; Fanelli, V.; Montemurro, C.; Miazzi, M.M. How to choose a good marker to analyze the olive germplasm (Olea europaea L.) and derived products. Genes 2021, 12, 1474. [Google Scholar] [CrossRef]
- Rossi, R.; Leolini, C.; Costafreda-Aumedes, S.; Leolini, L.; Bindi, M.; Zaldei, A.; Moriondo, M. Performances evaluation of a low-cost platform for high-resolution plant phenotyping. Sensors 2020, 20, 3150. [Google Scholar] [CrossRef]
- Gómez-Gálvez, F.J.; Pérez-Mohedano, D.; de la Rosa-Navarro, R.; Belaj, A. High-throughput analysis of the canopy traits in the worldwide olive germplasm bank of Córdoba using very high-resolution imagery acquired from unmanned aerial vehicle (UAV). Sci. Hortic. 2021, 278, 109851. [Google Scholar] [CrossRef]
- Belaj, A.; Satovic, Z.; Rallo, L.; Trujillo, I. Genetic diversity and relationships in olive (Olea europaea L.) germplasm collections as determined by randomly amplified polymorphic DNA. Theor. Appl. Genet. 2002, 105, 638–644. [Google Scholar] [CrossRef]
- Besnard, G.; Hernández, P.; Khadari, B.; Dorado, G.; Savolainen, V. Genomic profiling of plastid DNA variation in the Mediterranean olive tree. BMC Plant Biol. 2011, 11, 80. [Google Scholar] [CrossRef] [Green Version]
- Niu, E.; Jiang, C.; Wang, W.; Zhang, Y.; Zhu, S. Chloroplast genome variation and evolutionary analysis of Olea europaea L. Genes 2020, 11, 879. [Google Scholar] [CrossRef]
- Cruz, F.; Julca, I.; Gómez-Garrido, J.; Loska, D.; Marcet-Houben, M.; Cano, E.; Galán, B.; Frias, L.; Ribeca, P.; Derdak, S.; et al. Genome sequence of the olive tree, Olea europaea. Gigascience 2016, 5, 29. [Google Scholar] [CrossRef]
- Unver, T.; Wu, Z.; Sterck, L.; Turktas, M.; Lohaus, R.; Li, Z.; Yang, M.; He, L.; Deng, T.; Escalante, F.J.; et al. Wild olive genome and oil biosynthesis. Proc. Natl. Acad. Sci. USA 2017, 114, E9413–E9422. [Google Scholar] [CrossRef] [Green Version]
- Rao, G.; Zhang, J.; Liu, X.; Lin, C.; Xin, H.; Xue, L.; Wang, C. De novo assembly of a new Olea europaea genome accession using nanopore sequencing. Hortic. Res. 2021, 8, 64. [Google Scholar] [CrossRef]
- Biton, I.; Doron-Faigenboim, A.; Jamwal, M.; Mani, Y.; Eshed, R.; Rosen, A.; Sherman, A.; Ophir, R.; Lavee, S.; Avidan, B.; et al. Development of a large set of SNP markers for assessing phylogenetic relationships between the olive cultivars composing the Israeli olive germplasm collection. Mol. Breed. 2015, 35, 107. [Google Scholar] [CrossRef]
- D’Agostino, N.; Taranto, F.; Camposeo, S.; Mangini, G.; Fanelli, V.; Gadaleta, S.; Miazzi, M.M.; Pavan, S.; di Rienzo, V.; Sabetta, W.; et al. GBS-derived SNP catalogue unveiled wide genetic variability and geographical relationships of Italian olive cultivars. Sci. Rep. 2018, 8, 15877. [Google Scholar] [CrossRef] [Green Version]
- Taranto, F.; D’Agostino, N.; Pavan, S.; Fanelli, V.; di Rienzo, V.; Sabetta, W.; Miazzi, M.M.; Zelasco, S.; Perri, E.; Montemurro, C. Single nucleotide polymorphism (SNP) diversity in an olive germplasm collection. Acta Hortic. 2018, 1199, 27–32. [Google Scholar] [CrossRef]
- Zhu, S.; Niu, E.; Shi, A.; Mou, B. Genetic diversity analysis of olive germplasm (Olea europaea L.) with genotyping-by-sequencing technology. Front. Genet. 2019, 10, 755. [Google Scholar] [CrossRef] [Green Version]
- Kaya, H.B.; Akdemir, D.; Lozano, R.; Cetin, O.; Kaya, H.S.; Sahin, M.; Smith, J.L.; Tanyolac, B.; Jannink, J.L. Genome wide association study of 5 agronomic traits in olive (Olea europaea L.). Sci. Rep. 2019, 9, 18764. [Google Scholar] [CrossRef] [Green Version]
- Kyriakopoulou, C.I.; Kalogianni, D.P. Genetic identification of the wild form of olive (Olea europaea var. sylvestris) using allele-specific real-time PCR. Foods 2020, 9, 467. [Google Scholar] [CrossRef] [Green Version]
- Julca, I.; Marcet-Houben, M.; Cruz, F.; Gómez-Garrido, J.; Gaut, B.S.; Díez, C.M.; Gut, I.G.; Alioto, T.S.; Vargas, P.; Gabaldón, T. Genomic evidence for recurrent genetic admixture during the domestication of Mediterranean olive trees (Olea europaea L.). BMC Biol. 2020, 18, 148. [Google Scholar] [CrossRef] [PubMed]
- Saddoud Debbabi, O.; Montemurro, C.; Maachia, S.B.; Amar, F.B.; Fanelli, V.; Gadaleta, S.; El Riachy, M.; Chehade, A.; Siblini, M.; Boucheffa, S.; et al. A hot spot of olive biodiversity in the Tunisian Oasis of Degache. Diversity 2020, 12, 358. [Google Scholar] [CrossRef]
- Wright, S. The genetical structure of populations. Ann. Eugen. 1949, 15, 323–354. [Google Scholar] [CrossRef]
- Nei, M. Definition and estimation of fixation indices. Evolution 1986, 40, 643–645. [Google Scholar] [CrossRef]
- Neyeloff, J.L.; Fuchs, S.C.; Moreira, L.B. Meta-analyses and Forest plots using a Microsoft excel spreadsheet: Step-by-step guide focusing on descriptive data analysis. BMC Res. Notes 2012, 5, 52. [Google Scholar] [CrossRef] [Green Version]
- Montemurro, C.; Dambruoso, G.; Bottalico, G.; Sabetta, W. Self-incompatibility assessment of some Italian olive genotypes (Olea europaea L.) and cross-derived seedling selection by SSR markers on seed endosperms. Front. Plant Sci. 2019, 451, 10. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, D. Effects of inbreeding on the genetic diversity of populations. Phil. Trans. R. Soc. B 2003, 358, 1051–1070. [Google Scholar] [CrossRef] [Green Version]
- Keller, L.F.; Waller, D.M. Inbreeding effects in wild populations. Trends Ecol. Evol. 2002, 17, 230–241. [Google Scholar] [CrossRef]
- Maxted, N.; Kell, S.; Ford-Lloyd, B.; Dulloo, E.; Toledo, A. Toward the systematic conservation of Global Crop Wild Relative Diversity. Crop Sci. 2012, 52, 774–785. [Google Scholar] [CrossRef]
- Castañeda-Álvarez, N.P.; Khoury, C.K.; Achicanoy, H.A.; Bernau, V.; Dempewolf, H.; Eastwood, R.J.; Guarino, L.; Harker, R.H.; Jarvis, A.; Maxted, N.; et al. Global conservation priorities for crop wild relatives. Nat. Plants 2016, 2, 16022. [Google Scholar] [CrossRef]
- Di Rienzo, V.; Miazzi, M.M.; Fanelli, V.; Sabetta, W.; Montemurro, C. The preservation and characterization of Apulian olive germplasm biodiversity. Acta Hortic. 2018, 1199, 1–6. [Google Scholar] [CrossRef]
- Lotti, C.; Iovieno, P.; Centomani, I.; Marcotrigiano, A.R.; Fanelli, V.; Mimiola, G.; Summo, C.; Pavan, S.; Ricciardi, L. Genetic, bio-agronomic, and nutritional characterization of kale (Brassica oleracea L. var. acephala) diversity in Apulia, Southern Italy. Diversity 2018, 10, 25. [Google Scholar] [CrossRef] [Green Version]
- Fanelli, V.; Volpicella, M.; Giampetruzzi, A.; Saldarelli, P.; Leoni, C.; Ceci, L.R.; di Rienzo, V.; Venerito, P.; Taranto, F.; Giannini, P.; et al. Valorization of autochthonous Apulian grapevine cultivars for spumante production. Acta Hortic. 2019, 1248, 457–462. [Google Scholar] [CrossRef]
- Wang, Y.; Rahsid, M.A.R.; Li, X.; Yao, C.; Lu, L.; Bai, J.; Li, Y.; Xu, N.; Ynag, Q.; Zhang, L.; et al. Collection and evaluation of genetic diversity and population structure of potato landraces and varieties in China. Front. Plant Sci. 2019, 10, 139. [Google Scholar] [CrossRef] [Green Version]
- Volynkin, V.; Polulyakh, A.; Levchenko, S.; Vasylyk, I.; Likhovskoi, V. Autochthonous grape species, varieties and cultivars of Crimea. Acta Hortic. 2019, 1259, 91–98. [Google Scholar] [CrossRef]
- Miazzi, M.M.; di Rienzo, V.; Mascio, I.; Montemurro, C.; Sion, S.; Sabetta, W.; Vivaldi, G.A.; Camposeo, S.; Caponio, F.; Squeo, G.; et al. Re.Ger.OP: An Integrated Project for the Recovery of Ancient and Rare Olive Germplasm. Front. Plant Sci. 2020, 11, 73. [Google Scholar] [CrossRef] [Green Version]
- De la Rosa, R.; Klepo, T.; Arias-Calderón, R.; Toumi, A.; Domínguez-García, M.; Leon, L.; Belaj, A.; Mariotti, R.; Baldoni, L.; Satovic, Z.; et al. Current status of conservation, evaluation and usefulness of wild olive germplasm. Acta Hortic. 2014, 1057, 515–519. [Google Scholar] [CrossRef] [Green Version]
- El Bakkali, A.; Essalouh, L.; Tollon, C.; Rivallan, R.; Mournet, P.; Moukhli, A.; Zaher, H.; Mekkaoui, A.; Hadidou, A.; Sikaoui, L.; et al. Characterization of Worldwide Olive Germplasm Banks of Marrakech (Morocco) and Cordoba (Spain): Towards management and use of olive germplasm in breeding programs. PLoS ONE 2019, 14, e0223716. [Google Scholar] [CrossRef] [Green Version]
- López-Yerena, A.; Ninot, A.; Lozano-Castellón, J.; Escribano-Ferrer, E.; Romero-Aroca, A.J.; Belaj, A.; Vallverdú-Queralt, A.; Lamuela-Raventós, R.M. Conservation of Native Wild Ivory-White Olives from the MEDES Islands Natural Reserve to Maintain Virgin Olive Oil Diversity. Antioxidants 2020, 9, 1009. [Google Scholar] [CrossRef]
- Haouane, H.; El Bakkali, A.; Moukhli, A.; Tollon, C.; Santoni, S.; Oukabli, A.; El Modafar, C.; Khadari, B. Genetic structure and core collection of the World Olive Germplasm Bank of Marrakech: Towards the optimised management and use of Mediterranean olive genetic resources. Genetica 2011, 139, 1083–1094. [Google Scholar] [CrossRef] [Green Version]
- International Olive Council. Book of the IOC network of germplasm banks. In Proceedings of the International Seminar the IOC Network of Germplasm Banks and the True Healthy Olive Cultivars Project, Cordoba, Spain, 21–24 October 2019. [Google Scholar]
- Saddoud Debbabi, O.; Rahmani Mnasri, S.; Ben Amar, F.; Ben Naceur, M.; Montemurro, C.; Miazzi, M.M. Applications of microsatellite markers for the characterization of olive genetic resources of Tunisia. Genes 2021, 12, 286. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.M.; van der Werf, W.; Lansink, A.O. Assessment of the environmental impacts of Xylella fastidiosa subsp. pauca in Puglia. Crop. Prot. 2021, 142, 105519. [Google Scholar] [CrossRef]
- Barazani, O.; Westberg, E.; Hanin, N.; Dag, A.; Kerem, Z.; Tugendhaft, Y.; Hmidat, M.; Hijawi, T.; Kadereit, J.W. A comparative analysis of genetic variation in rootstocks and scions of old olive trees—A window into the history of olive cultivation practices and past genetic variation. BMC Plant Biol. 2014, 14, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
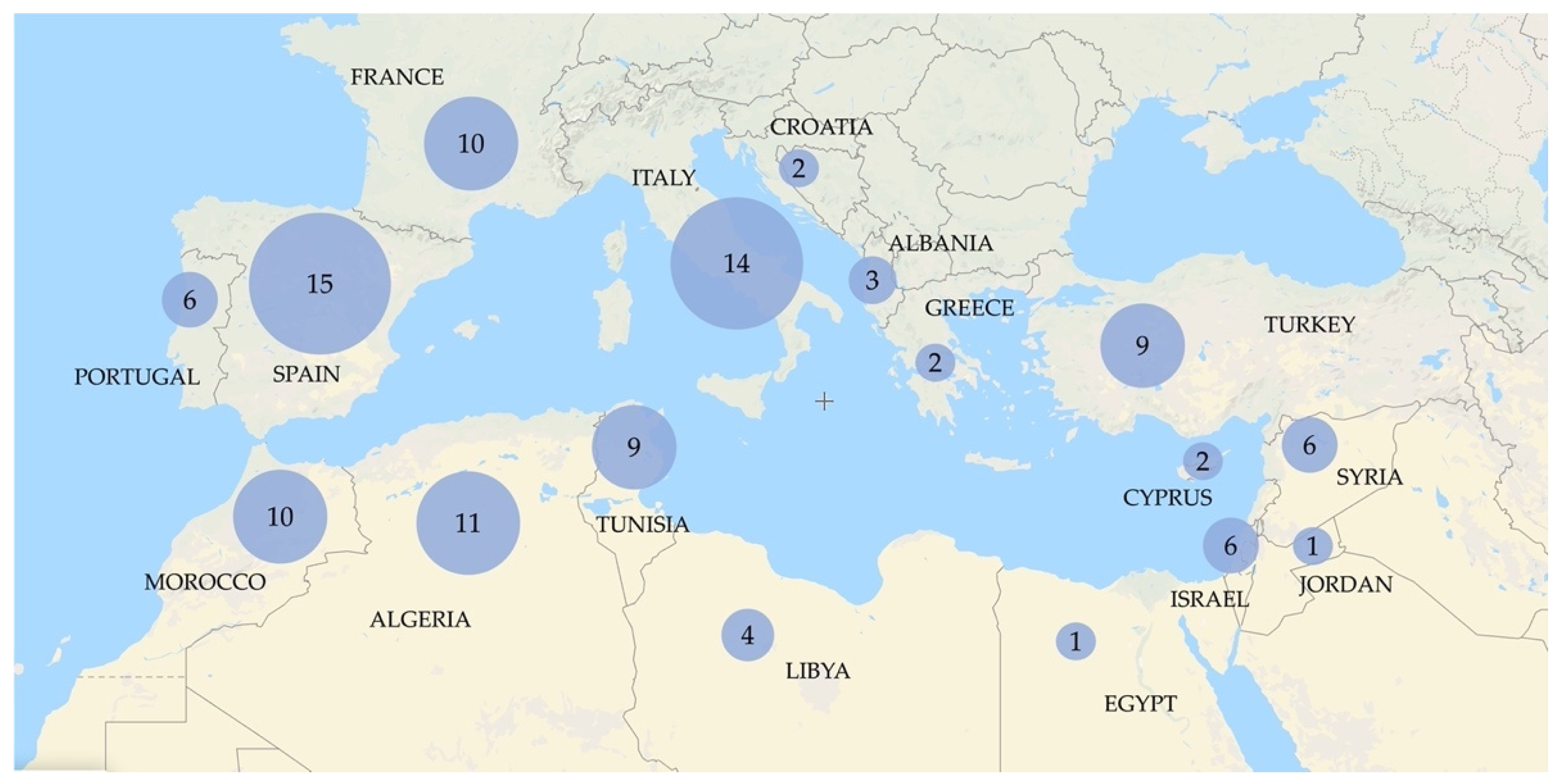
| Reference | Analyzed Subspecies | Analyzed Varieties of Subspecies europaea | Number of Analyzed Oleaster Accessions | Used Markers |
|---|---|---|---|---|
| Amane et al., 2000 [29] | europaea, laperrinei | europaea and sylvestris | 45 | RFLP |
| Besnard and Bervillé, 2000 [11] | europaea, maroccana, laperrinei, cerasiformis, cuspidata | europaea and sylvestris | 300 | RAPD, RFLP |
| Lumaret et al., 2000 [30] | europaea, maroccana, laperrinei, cuspidata | europaea and sylvestris | 101 | RFLP |
| Besnard et al., 2001 [31] | europaea, maroccana, laperrinei, cerasiformis, cuspidata | europaea and sylvestris | 292 | RAPD, RFLP |
| Vargas and Kadereit, 2001 [32] | europaea, maroccana, laperrinei, cerasiformis, cuspidata, guanchica | europaea and sylvestris | 26 | ISSR |
| Besnard and Bervillé, 2002 [33] | europaea, maroccana, laperrinei, cerasiformis, cuspidata | europaea and sylvestris | 7 | RFLP |
| Bronzini de Caraffa et al., 2002 [12] | europaea | europaea and sylvestris | 99 | RAPD, RFLP |
| Baldoni et al., 2006 [34] | europaea | europaea and sylvestris | 100 | AFLP |
| Breton et al., 2006 [35] | europaea | sylvestris | 166 | SSR |
| Belaj et al., 2007 [36] | europaea | sylvestris | 171 | SSR |
| Brito et al., 2008 [37] | europaea, cerasiformis, guanchica | europaea and sylvestris | 8 | SSR |
| Hannachi et al., 2008 [38] | europaea | europaea and sylvestris | 70 | Morphological, SSR |
| Hannachi et al., 2009 [39] | europaea, cuspidata | europaea and sylvestris | 12 | Morphological, SSR |
| Belaj et al., 2010 [40] | europaea | europaea and sylvestris | 107 | SSR |
| Erre et al., 2010 [41] | europaea | europaea and sylvestris | 21 | SSR |
| Hannachi et al., 2010 [42] | europaea | europaea and sylvestris | 52 | SSR |
| Sesli and Yegenoglu, 2010 [43] | europaea | europaea and sylvestris | 8 | RAPD |
| Belaj et al., 2011 [44] | europaea | sylvestris | 48 | Morphological, SSR |
| Ismaili et al., 2012 [45] | europaea | sylvestris | 27 | Morphological |
| Besnard et al., 2013 [46] | europaea, laperrinei | europaea and sylvestris | 390 | SSR |
| Diez et al., 2015 [5] | europaea | europaea and sylvestris | 96 | SSR |
| Beghè et al., 2017 [47] | europaea | sylvestris | 225 | SSR |
| Boucheffa et al., 2017 [48] | europaea | europaea and sylvestris | 16 | SSR |
| Chiappetta et al., 2017 [13] | europaea | europaea and sylvestris | 99 | SSR |
| Belaj et al., 2018 [49] | europaea | europaea and sylvestris | 89 | SNP |
| di Rienzo et al., 2018 [50] | europaea | europaea and sylvestris | 16 | SSR |
| Ismaili et al., 2018 [51] | europaea | europaea and sylvestris | 61 | Morphological, SSR |
| Boucheffa et al., 2019 [52] | europaea | europaea and sylvestris | 12 | Morphological, SSR |
| Gros-Balthazard et al., 2019 [53] | europaea, cuspidata | europaea and sylvestris | 27 | SNP |
| Díaz-Rueda et al., 2020 [24] | europaea, maroccana, laperrinei, cerasiformis, cuspidata, guanchica | europaea and sylvestris | 59 | Morphological, SSR |
| Mariotti et al., 2020 [54] | europaea, guanchica | europaea and sylvestris | 73 | SNP |
| Rodrigues et al., 2020 [55] | europaea | sylvestris | 12 | Morphological |
| Dervishi et al., 2021 [56] | europaea | europaea and sylvestris | 19 | SSR |
| Khouatmiani et al., 2021 [57] | europaea | sylvestris | 24 | Morphological |
| Falek et al., 2022 [14] | europaea | sylvestris | 174 | SSR |
| Key Findings | Reference |
|---|---|
| Multiple domestication events took place in olive | Diez et al., 2015 [5] Gros-Balthazard et al., 2019 [53] Julca et al., 2020 [73] |
| Wild olive includes feral forms and genuine wild olives | Bronzini de Caraffa et al., 2002 [12] Baldoni et al., 2006 [34] Breton et al., 2006 [35] Chiappetta et al., 2017 [13] |
| Genuine oleasters show a much higher level of morphological and genetic variability compared to cultivated olives | Hannachi et al., 2009 [39] Belaj et al., 2010 [40] Belaj et al., 2011 [44] Besnard et al., 2013 [46] Díaz-Rueda et al., 2020 [24] |
| A constant gene flow takes place in the regions in which wild and cultivated olives coexist, making the distinction difficult between genuinely wild and feral olive | Besnard et al., 2001 [31] Bronzini de Caraffa et al., 2002 [12] Breton et al., 2006 [35] Belaj et al., 2007 [36] Boucheffa et al., 2017 [48] |
| Wild olive is an important source of traits related to biotic and abiotic stress tolerances. | Beghè et al., 2017 [47] Mariotti et al., 2020 [54] Falek et al., 2022 [14] |
| Institution | Country | Strategy of Conservation |
|---|---|---|
| WOGC—IFAPA, Worldwide Olive Germplasm Bank of Córdoba | Spain | Ex situ |
| WOGB—INRA, Worldwide Olive Germplasm Bank of Marrakech | Morocco | Ex situ |
| WOGB—Worldwide Olive Germplasm Bank of Izmir | Turkey | Ex situ |
| National Gene Bank of Tunisia (NGBT) | Tunisia | Ex situ and in situ |
| Olive Gene Bank of Albania | Albania | Ex situ and in situ |
| National Olive Germplasm Bank of Turkey | Turkey | Ex situ and in situ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fanelli, V.; Mascio, I.; Falek, W.; Miazzi, M.M.; Montemurro, C. Current Status of Biodiversity Assessment and Conservation of Wild Olive (Olea europaea L. subsp. europaea var. sylvestris). Plants 2022, 11, 480. https://doi.org/10.3390/plants11040480
Fanelli V, Mascio I, Falek W, Miazzi MM, Montemurro C. Current Status of Biodiversity Assessment and Conservation of Wild Olive (Olea europaea L. subsp. europaea var. sylvestris). Plants. 2022; 11(4):480. https://doi.org/10.3390/plants11040480
Chicago/Turabian StyleFanelli, Valentina, Isabella Mascio, Wahiba Falek, Monica Marilena Miazzi, and Cinzia Montemurro. 2022. "Current Status of Biodiversity Assessment and Conservation of Wild Olive (Olea europaea L. subsp. europaea var. sylvestris)" Plants 11, no. 4: 480. https://doi.org/10.3390/plants11040480
APA StyleFanelli, V., Mascio, I., Falek, W., Miazzi, M. M., & Montemurro, C. (2022). Current Status of Biodiversity Assessment and Conservation of Wild Olive (Olea europaea L. subsp. europaea var. sylvestris). Plants, 11(4), 480. https://doi.org/10.3390/plants11040480