
Morphological and Eco-Geographic Variation in Algerian Wild Olives

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
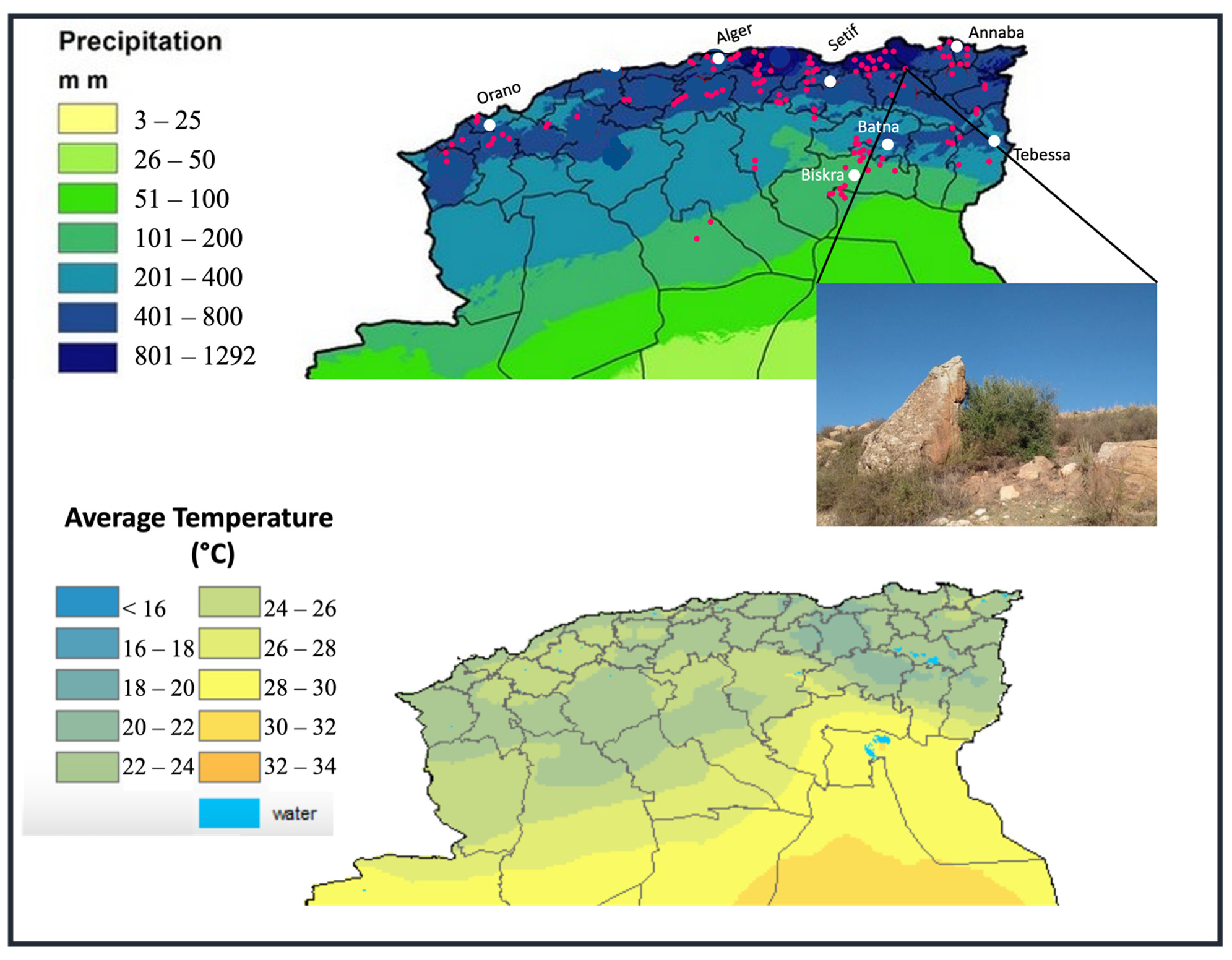
2.1. Field Surveys and Sampling
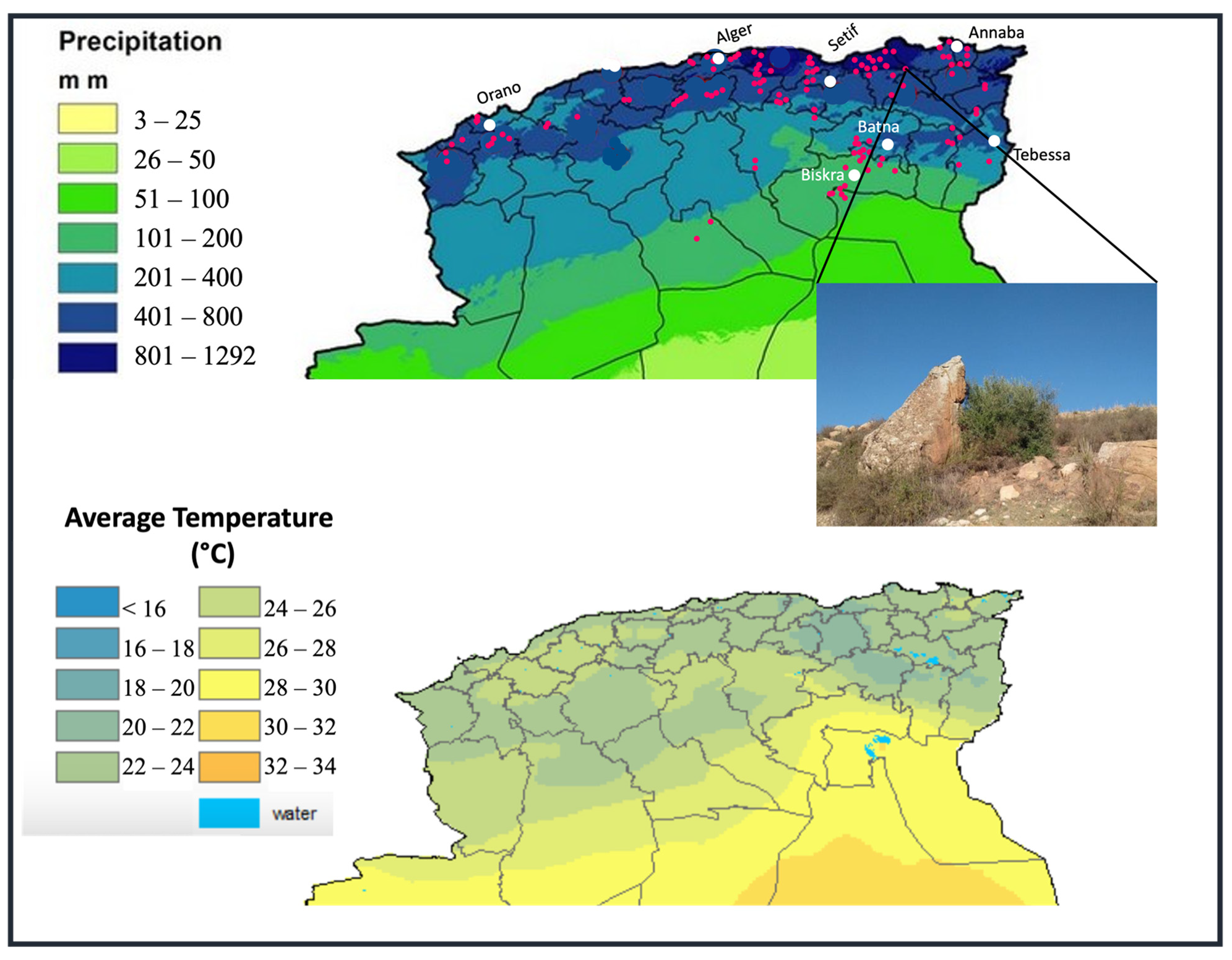
2.2. Ecogeographic Data
2.3. Morphological Quantitative and Qualitative Traits
2.4. Data Analysis
3. Results
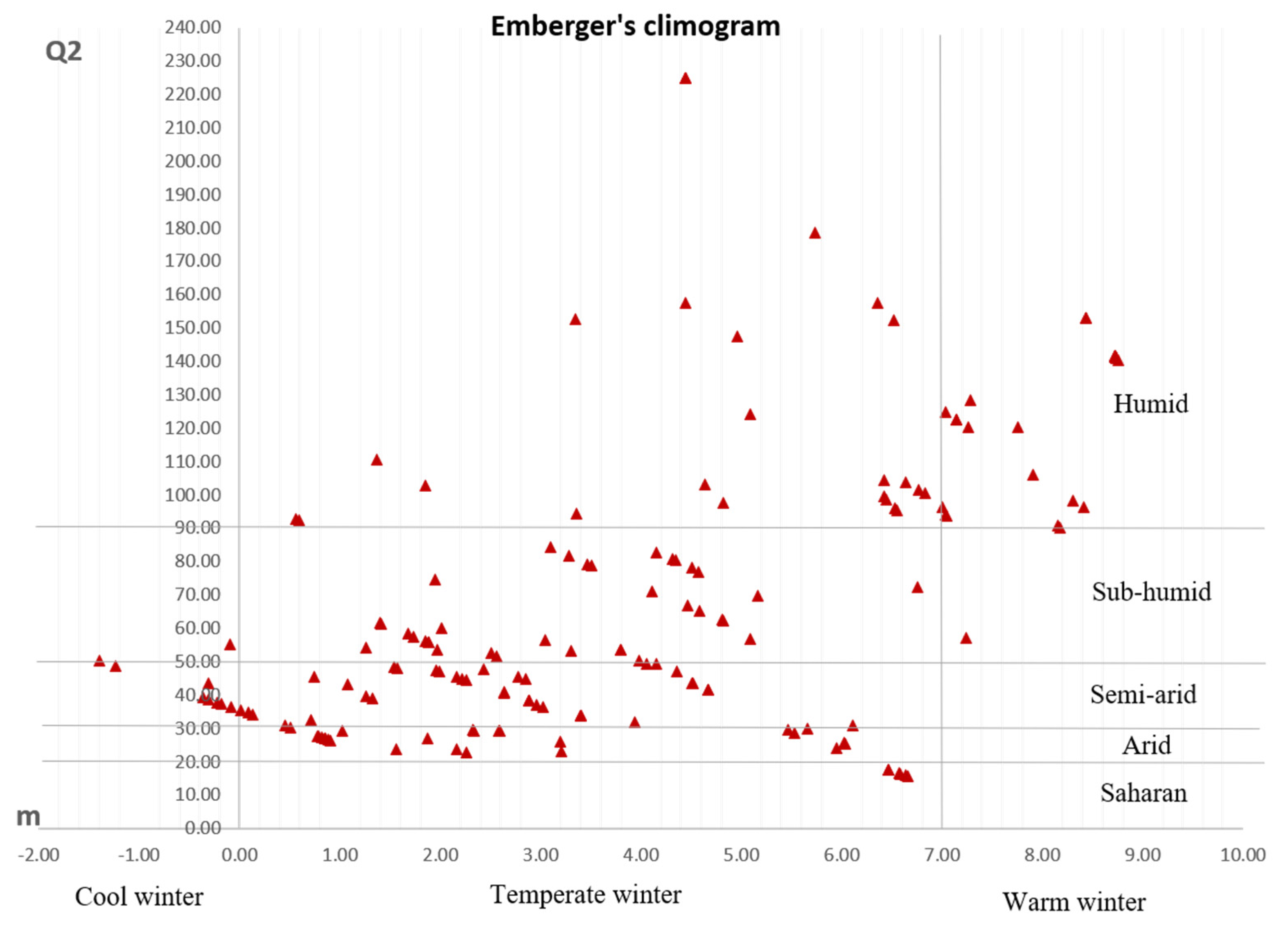
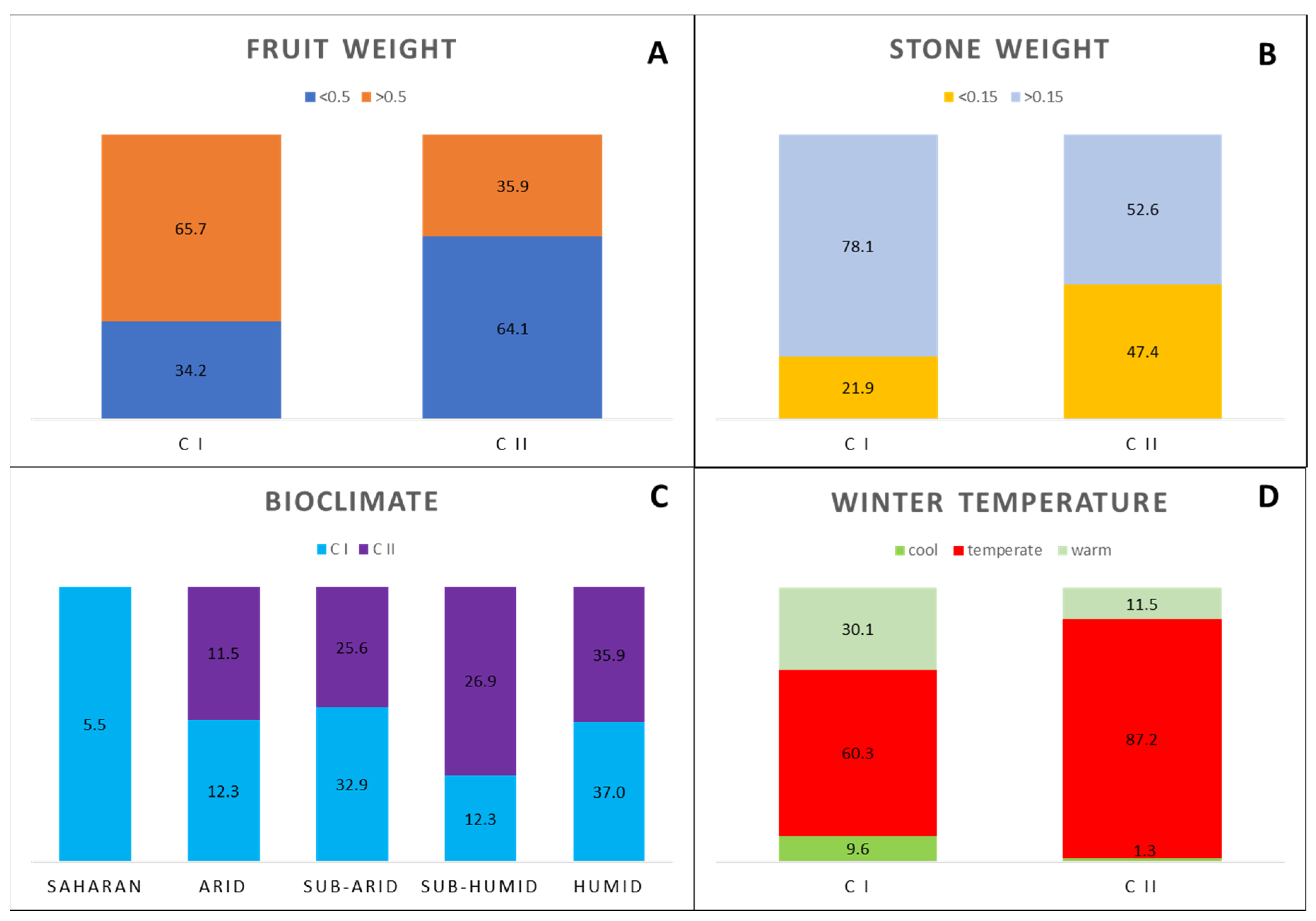
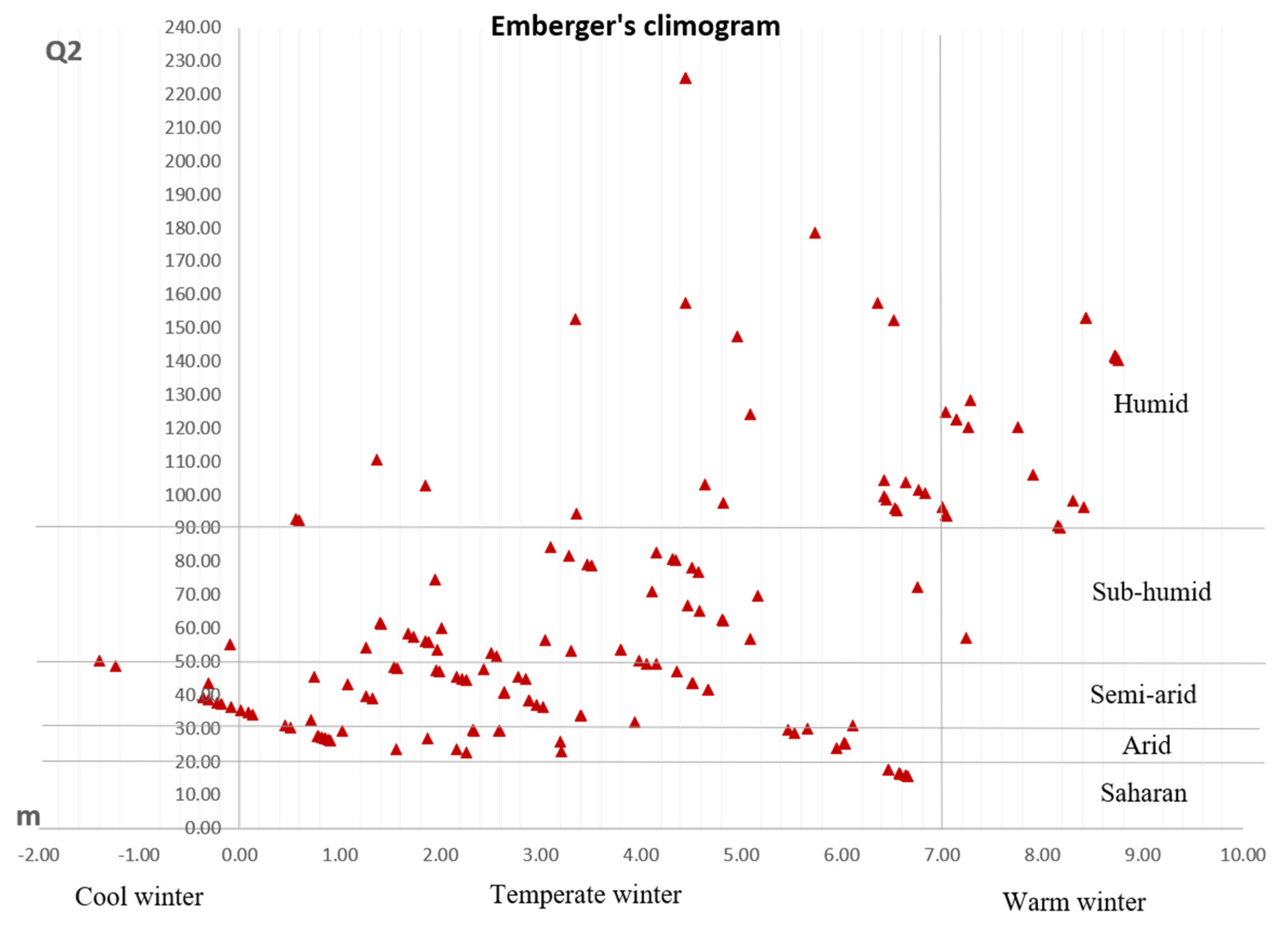
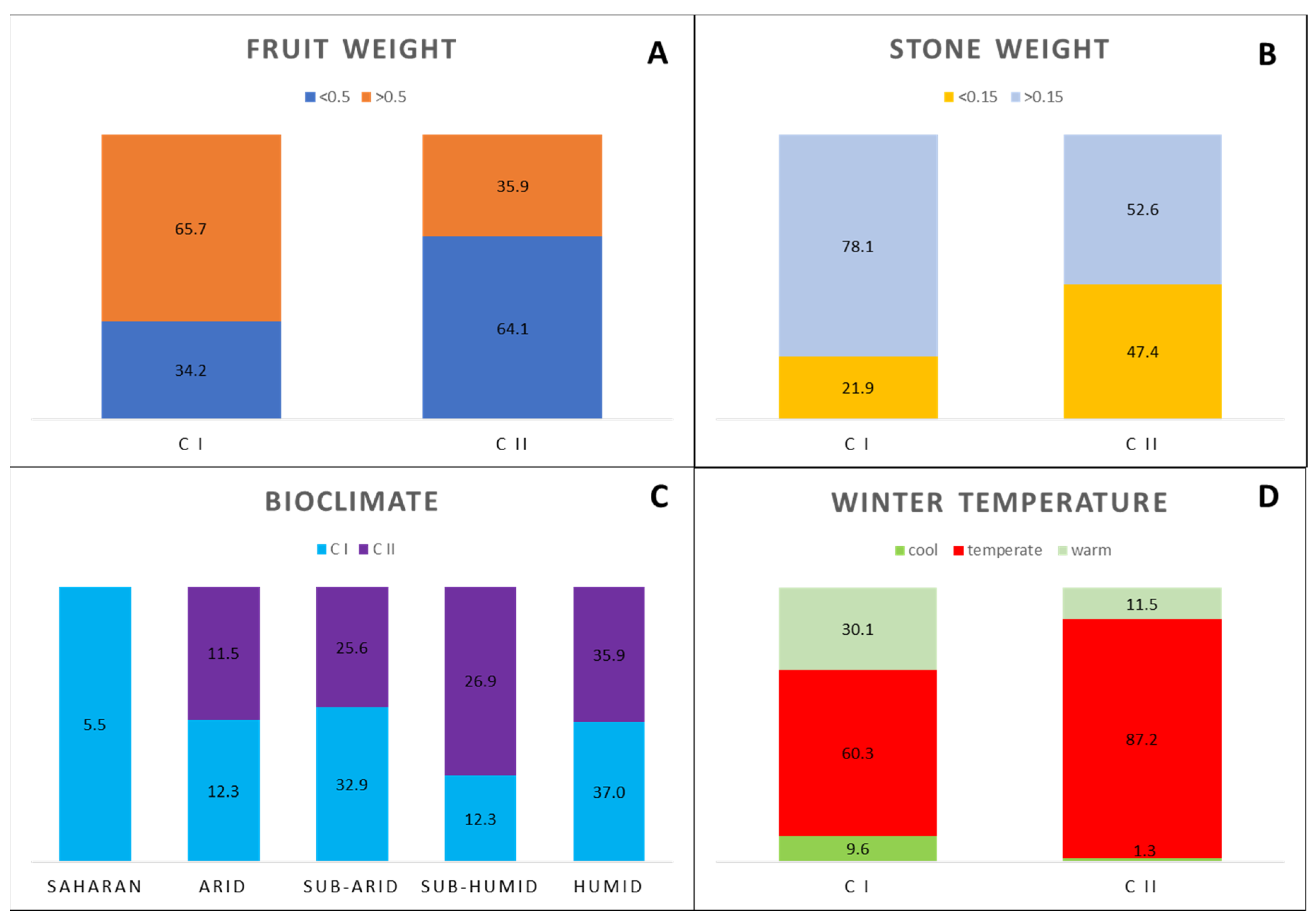
3.1. Ecogeographic Data Analysis
3.2. Quantitative Traits Analysis
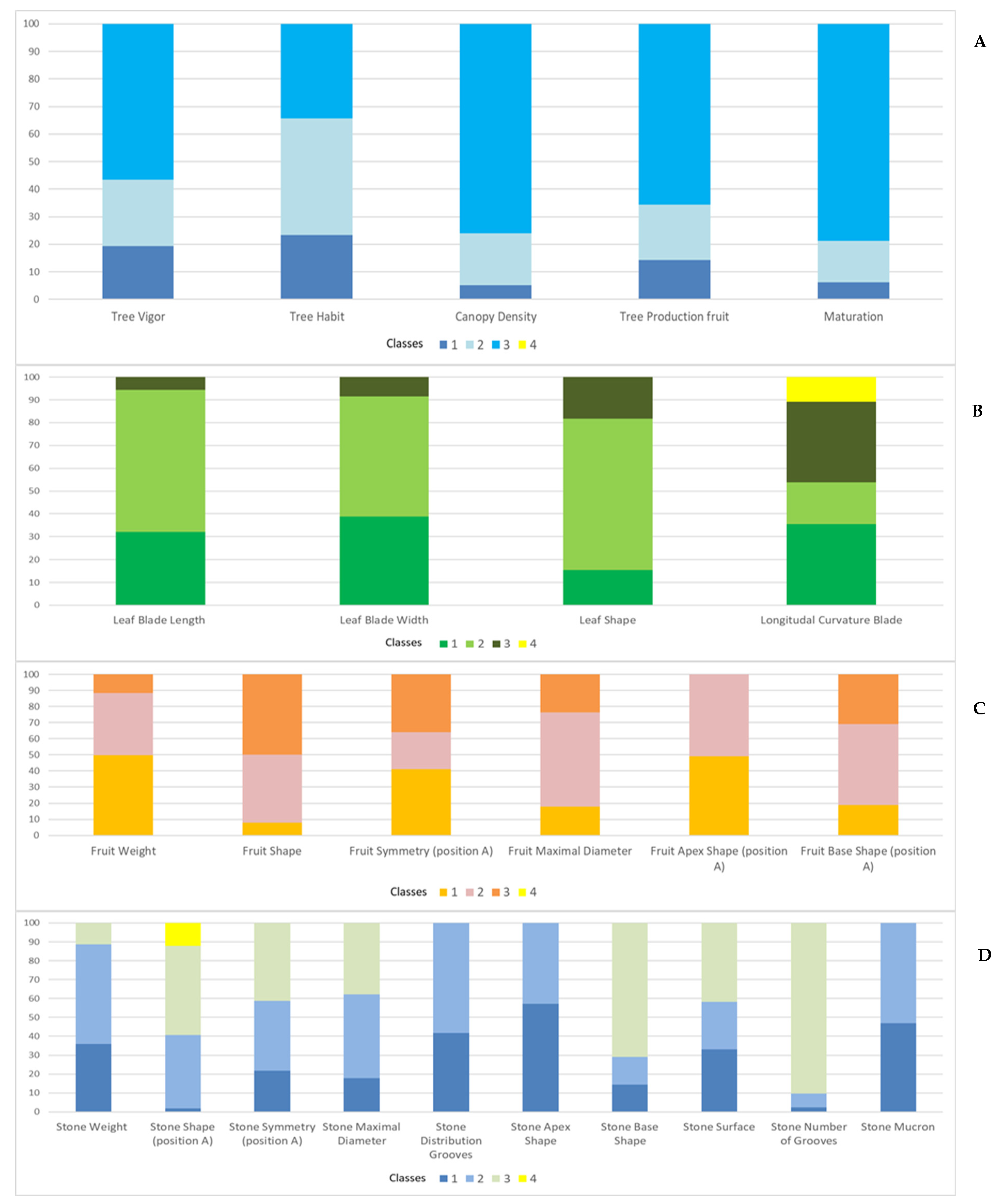
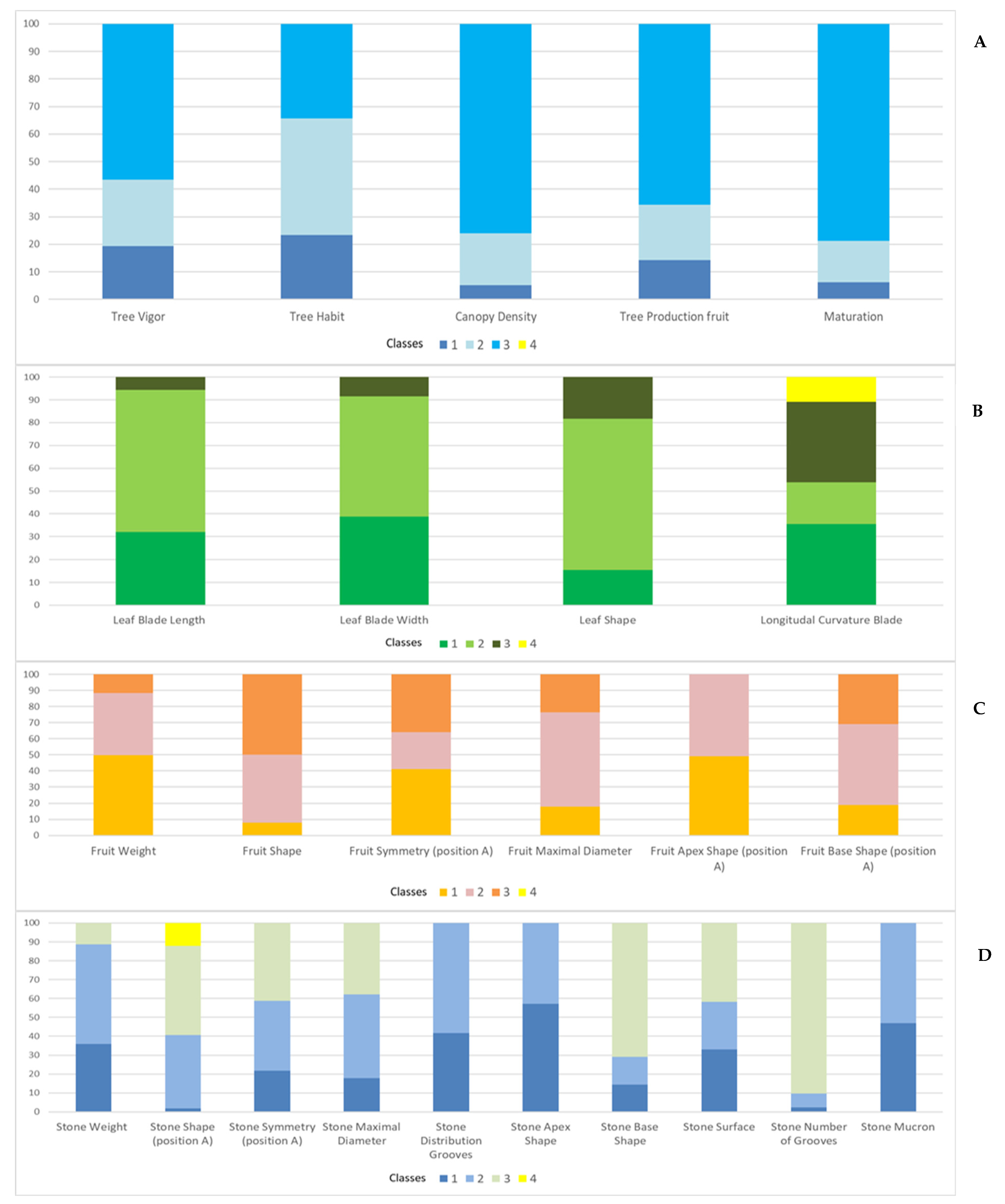
3.3. Qualitative Traits
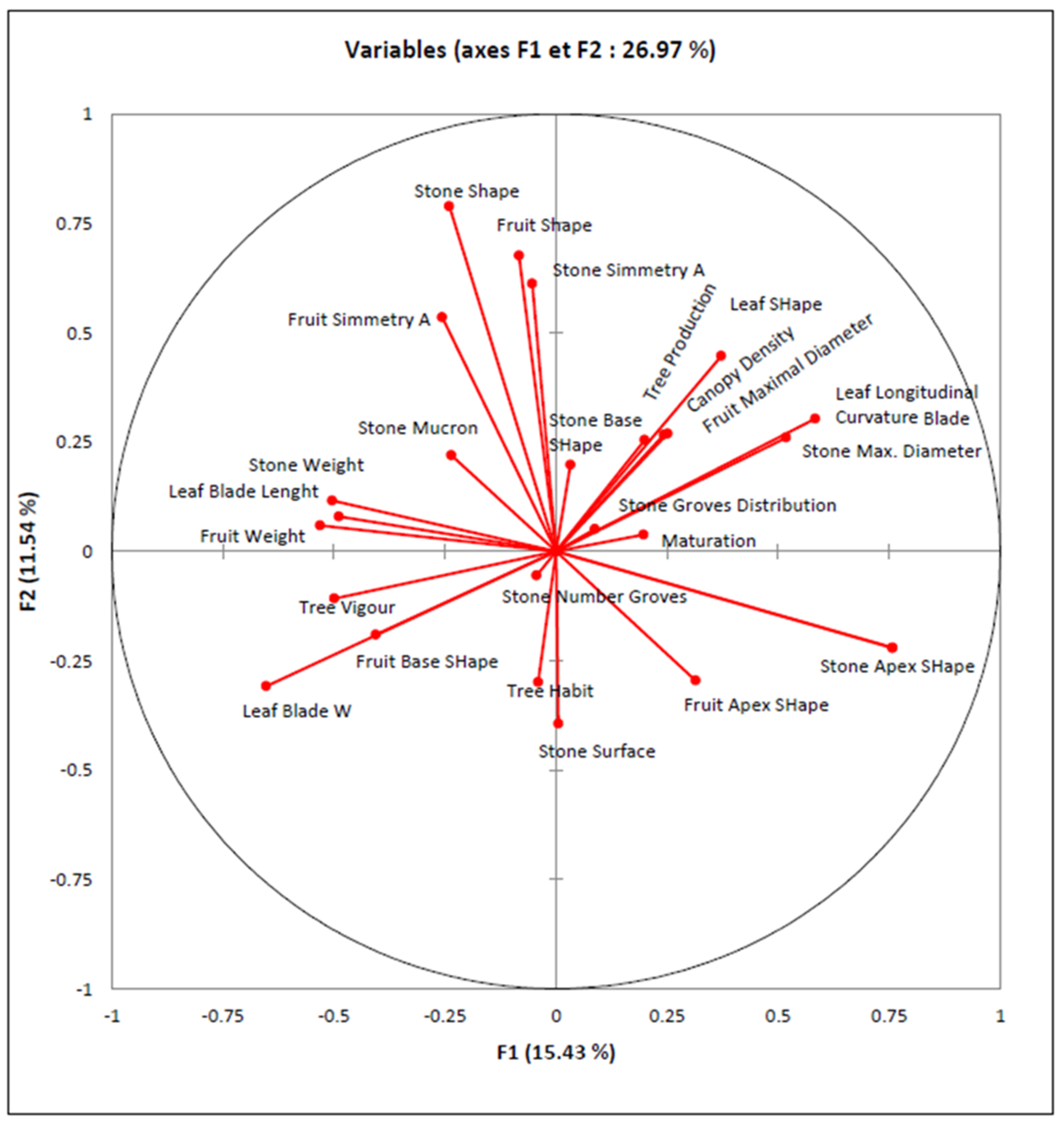
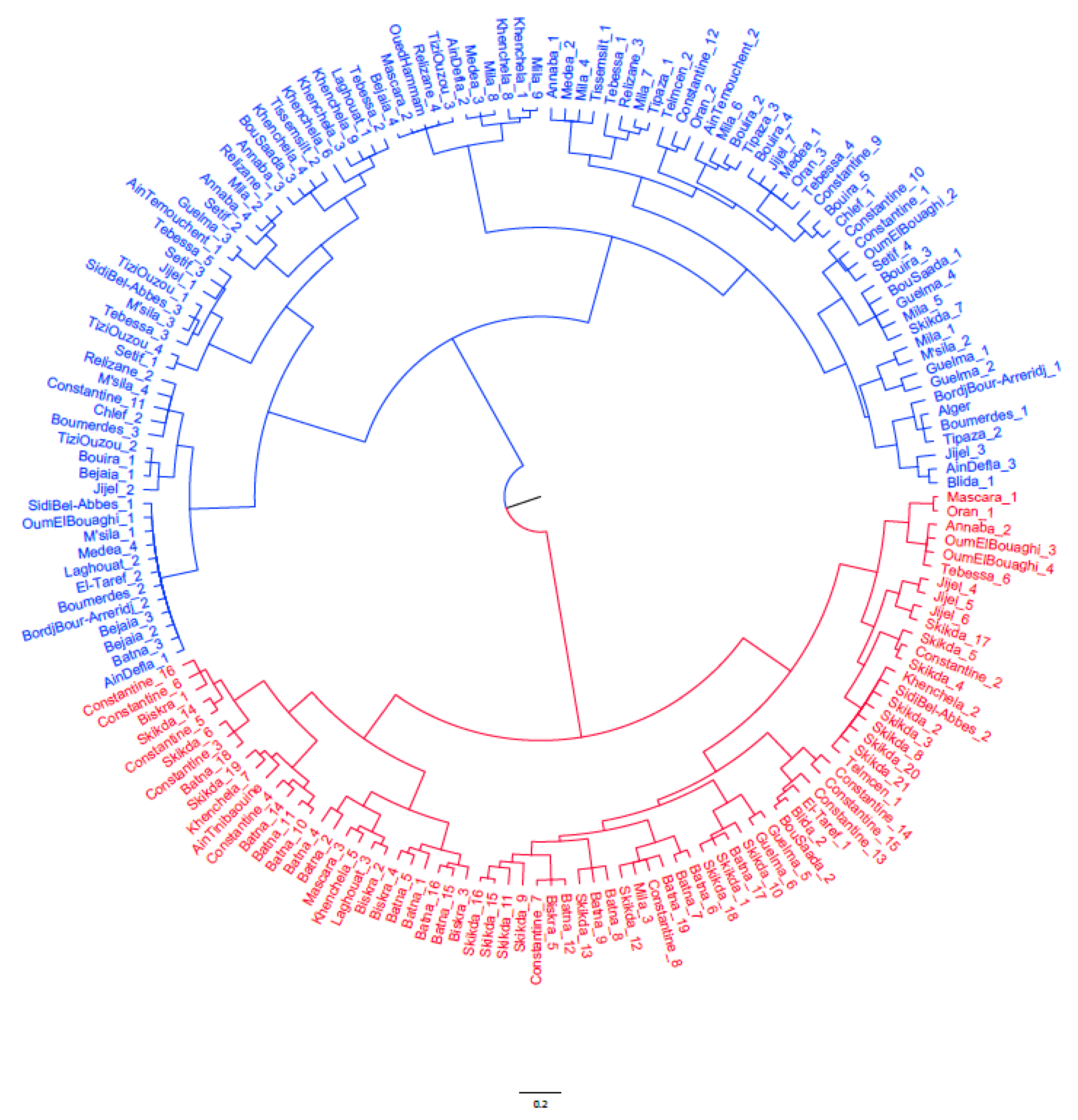
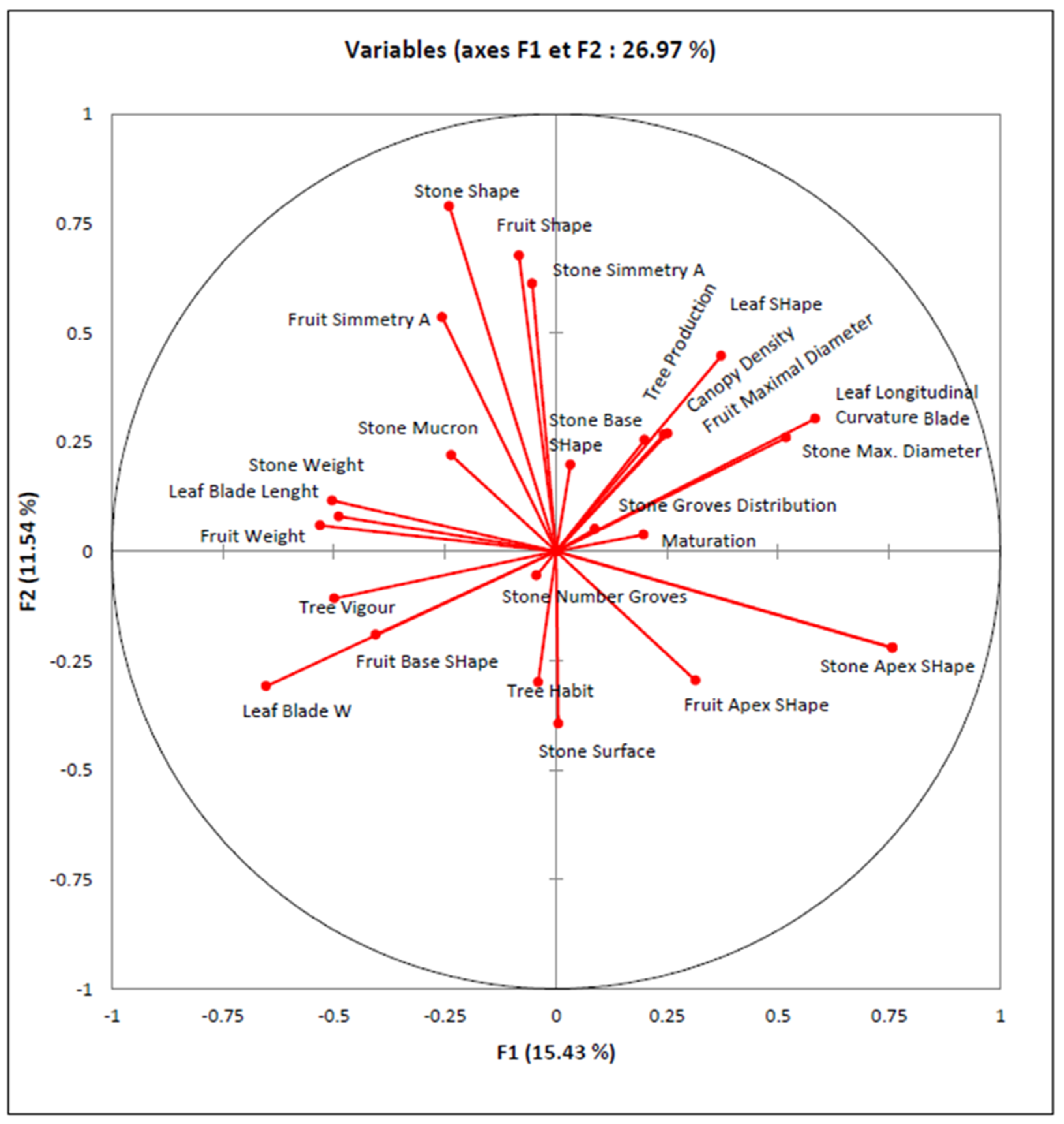
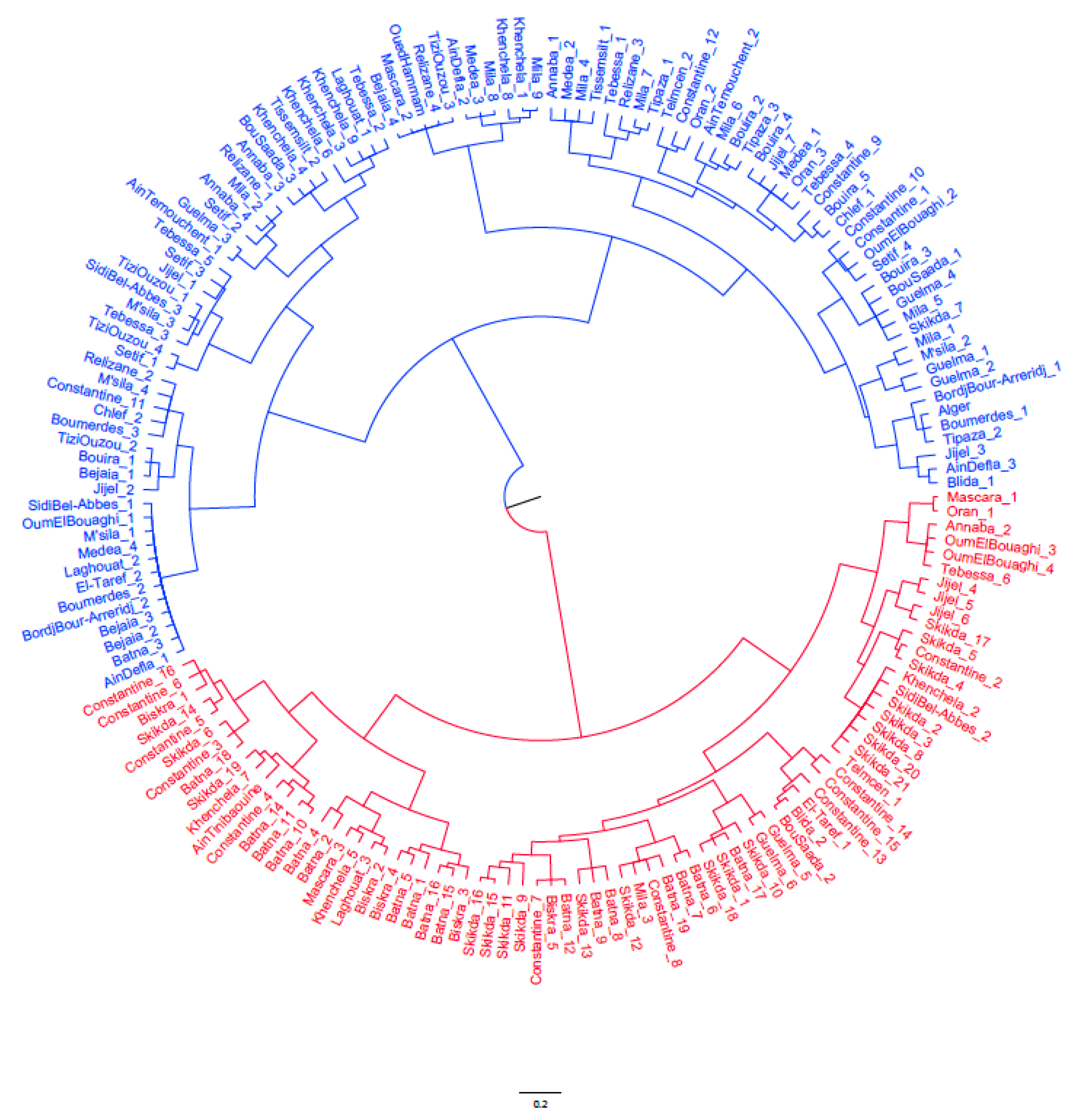
3.4. Cluster Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jukić Špika, M.; Liber, Z.; Montemurro, C.; Miazzi, M.M.; Ljubenkov, I.; Soldo, B.; Žanetić, M.; Vitanović, E.; Politeo, O.; Škevin, D. Quantitatively Unraveling Hierarchy of Factors Impacting Virgin Olive Oil Phenolic Profile and Oxidative Stability. Antioxidants 2022, 11, 594. [Google Scholar] [CrossRef] [PubMed]
- Green, P.S. A revision of Olea L. (Oleaceae). Kew Bull. 2002, 57, 91–140. [Google Scholar] [CrossRef]
- Besnard, G.; Rubio de Casas, R. Single vs. multiple independent olive domestications: The jury is (still) out. New Phytol. 2016, 209, 466–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terral, J.F.; Arnold-Simard, G. Beginnings of olive cultivation in eastern Spain in relation to Holocene bioclimatic changes. Quat. Res. 1996, 46, 176–185. [Google Scholar] [CrossRef]
- Baldoni, L.; Tosti, N.; Ricciolini, C.; Belaj, A.; Arcioni, S.; Pannelli, G.; Germanà, M.A.; Mulas, M.; Porceddu, A. Genetic structure of wild and cultivated olives in the Central Mediterranean Basin. Ann. Bot. 2006, 98, 935–942. [Google Scholar] [CrossRef]
- Klepo, T.; De la Rosa, R.; Satovic, Z.; León, L.; Belaj, A. Utility of wild germplasm in olive breeding. Sci. Hortic. 2013, 152, 92–101. [Google Scholar] [CrossRef]
- Besnard, G.; Khadari, B.; Navascués, M.; Fernández-Mazuecos, M.; El Bakkali, A.; Arrigo, N.; Baali-Cherif, D.; De Caraffa, V.B.-B.; Santoni, S.; Vargas, P.; et al. The complex history of the olive tree: From Late Quaternary diversification of Mediterranean lineages to primary domestication in the northern Levant. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122833. [Google Scholar] [CrossRef] [Green Version]
- Aranda, S.; Montes-Borrego, M.; Jiménez-Díaz, R.M.; Landa, B.B. Microbial communities associated with the root system of wild olives (Olea europaea L. subsp. europaea var. sylvestris) are good reservoirs of bacteria with antagonistic potential against Verticillium dahliae. Plant Soil 2011, 343, 329–345. [Google Scholar]
- Arias-Calderón, R.; Rodríguez-Jurado, D.; León, L.; Bajarano-Alcázar, J.; De la Rosa, R.; Belaj, A. Pre-breeding for resistance to Verticillium wilt in olive: Fishing in the wild relative gene pool. Crop Prot. 2015, 75, 25–33. [Google Scholar] [CrossRef]
- Díaz-Rueda, P.; Franco-Navarro, J.; Messora, R.; Espartero, J.; Rivero-Núñez, C.; Aleza, P.; Capote, N.; Cantos, M.; García-Fernández, J.L.; de Cires, A.; et al. SILVOLIVE, a germplasm collection of wild subspecies with high genetic variability as a source of rootstocks and resistance genes for olive breeding. Front. Plant Sci. 2020, 11, 629. [Google Scholar] [CrossRef]
- Hannachi, H.; Nasri, N.; Elfalleh, W.; Tlili, N.; Ferchichi, A.; Msallem, M. Fatty acids, sterols, polyphenols and chlorophylls of olive oils obtained from Tunisian wild olive trees (Olea europaea L. var. sylvestris). Int. J. Food Prop. 2013, 16, 1271–1283. [Google Scholar] [CrossRef]
- Baccouri, B.; Guerfel, M.; Zarrouk, W.; Taamalli, W.; Daoud, D.; Zarrouk, M. Wild olive (Olea europaea L.) selection for quality oil production. J. Food Biochem. 2011, 35, 161–176. [Google Scholar] [CrossRef]
- Dabbou, S.; Dabbou, S.; Selvaggini, R.; Urbani, S.; Taticchi, A.; Servili, M.; Hammami, M. Comparison of the chemical composition and the organoleptic profile of virgin olive oil from two wild and two cultivated Tunisian Olea Europaea. Chem. Biodivers. 2011, 8, 189–202. [Google Scholar] [CrossRef]
- Baali-Cherif, D.; Besnard, G. High Genetic Diversity and Clonal Growth in Relict Populations of Olea europaea subsp. laperrinei (Oleaceae) from Hoggar, Algeria. Ann. Bot. 2005, 96, 823–830. [Google Scholar] [PubMed]
- Besnard, G.; Anthelme, F.; Baali-Cherif, D. The Laperrine’s olive tree (Oleaceae): A wild genetic resource of the cultivated olive and a model-species for studying the biogeography of the Saharan Mountains. Acta Bot. Gall. 2012, 159, 319–328. [Google Scholar] [CrossRef]
- Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 28 May 2022).
- Mendil, M.; Sebai, A. Catalogue des Variétés Algérienne de L’olivier: L’olivier en Algérie; Institut Technique de l’Arboriculture Fruitière et de la vigne (ITAF): Algiers, Algeria, 2006; Dépôt légal N° 1840. [Google Scholar]
- Boucheffa, S.; Miazzi, M.M.; di Rienzo, V.; Mangini, G.; Fanelli, V.; Tamendjari, A.; Pignone, D.; Montemurro, C. The coexistence of oleaster and traditional varieties affects genetic diversity and population structure in Algerian olive (Olea europaea) germplasm. Genet. Resour. Crop Evol. 2017, 64, 379–390. [Google Scholar] [CrossRef]
- Di Rienzo, V.; Sion, S.; Taranto, F.; D’Agostino, N.; Montemurro, C.; Fanelli, V.; Sabetta, W.; Boucheffa, S.; Tamendjari, A.; Pasqualone, A.; et al. Genetic flow among olive populations within the Mediterranean basin di Rienzo. PeerJ 2018, 2018, e5260. [Google Scholar] [CrossRef] [Green Version]
- Di Rienzo, V.; Miazzi, M.M.; Fanelli, V.; Sabetta, W.; Montemurro, C. The preservation and characterization of Apulian olive germplasm biodiversity. Acta Hortic. 2018, 1199, 1–6. [Google Scholar] [CrossRef]
- Saddoud Debbabi, O.S.; Miazzi, M.M.; Elloumi, O.; Fendri, M.; Ben Amar, F.; Savoia, M.; Sion, S.; Souabni, H.; Mnasri, S.R.; Ben Abdelaali, S.; et al. Recovery, assessment, and molecular characterization of minor olive genotypes in Tunisia. Plants 2020, 9, 382. [Google Scholar] [CrossRef] [Green Version]
- Saddoud Deddabi, O.; Montemurro, C.; Ben Maachia, S.; Ben Amar, F.; Fanelli, V.; Gadaleta, S.; El Riachy, M.; Chehade, A.; Siblini, M.; Boucheffa, S.; et al. A hot spot of olive biodiversity in the Tunisian Oasis of Degache. Diversity 2020, 12, 358. [Google Scholar] [CrossRef]
- Miazzi, M.M.; di Rienzo, V.; Mascio, I.; Montemurro, C.; Sion, S.; Sabetta, W.; Vivaldi, G.A.; Camposeo, S.; Caponio, F.; Squeo, G.; et al. Ger.OP: An Integrated Project for the Recovery of Ancient and Rare Olive Germplasm. Front. Plant Sci. 2020, 11, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boucheffa, S.; Tamendjari, A.; Sanchez-Gimeno, A.C.; Rovellini, P.; Venturini, S.; di Rienzo, V.; Miazzi, M.M.; Montemurro, C. Diversity assessment of Algerian wild and cultivated olives (Olea europaea L.) by molecular, morphological, and chemical traits. Eur. J. Lipid Sci. Technol. 2019, 121, 1800302. [Google Scholar] [CrossRef] [Green Version]
- Falek, W.; Sion, S.; Montemurro, C.; Mascio, I.; Gadaleta, S.; Fanelli, V.; Savoia, M.A.; Piarulli, L.; Bechkri, S.; Khelifi, D.; et al. Molecular diversity and ecogeographic distribution of Algerian wild olives (Olea europaea subsp. europaea var. sylvestris). Sci. Agric. 2022, 79, e20200308. [Google Scholar] [CrossRef]
- D’Agostino, N.; Taranto, F.; Camposeo, S.; Mangini, G.; Fanelli, V.; Gadaleta, S.; Miazzi, M.M.; Pavan, S.; di Rienzo, V.; Sabetta, W.; et al. GBS-derived SNP catalogue unveiled wide genetic variability and geographical relationships of Italian olive cultivars. Sci. Rep. 2018, 8, 15877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taranto, F.; D’Agostino, N.; Pavan, S.; Fanelli, V.; Di Rienzo, V.; Sabetta, W.; Miazzi, M.M.; Zelasco, S.; Perri, E.; Montemurro, C. Single nucleotide polymorphism (SNP) diversity in an olive germplasm collection. VIII Int. Olive Symp. 2016, 27–31. [Google Scholar] [CrossRef]
- Sion, S.; Savoia, M.A.; Gadaleta, S.; Piarulli, L.; Mascio, I.; Fanelli, V.; Montemurro, C.; Miazzi, M.M. How to choose a good marker to analyze the olive germplasm (Olea europaea L.) and derived products. Genes 2021, 12, 1474. [Google Scholar] [CrossRef]
- Unver, T.; Wu, Z.; Sterck, L.; Turktas, M.; Lohaus, R.; Li, Z.; Yang, M.; He, L.; Deng, T.; Escalante, F.J.; et al. Wild olive genome and oil biosynthesis. Proc. Natl. Acad. Sci. USA 2017, 114, E9413–E9422. [Google Scholar] [CrossRef] [Green Version]
- Boukhari, R.; Ameur, A.A.; Innal, H.; Gaouar, S.B.S. First morphological characterization of autochthonous olive (Olea europaea L.) denominations from central and eastern of Algeria. Acta Agric. Slov. 2020, 116, 311–325. [Google Scholar] [CrossRef]
- Khouatmiani, K.; Belhadj, S.; Tonetto, A.; Assie, A.; Mevy, J.P.; Gauquelin, T. Variability of eight Algerian oleaster ecotypes (Olea europaea subsp. europaea var. sylvestris [Mill.] Lehr): Pollen and exine morphology in relation to geo-climatic effect. Grana 2021, 60, 256–270. [Google Scholar]
- UPOV. Guidelines for the Conduct of Tests for Distinctness, Homogeneity and Stability. 1985. Available online: www.upov.int (accessed on 20 October 2017).
- Bechkri, S.; Khelifi, D. Variation in Vicia sativa s.l. from Algeria based on morphological characters and ecogeographic parameters. Genet. Resour. Crop Evol. 2017, 64, 815–832. [Google Scholar] [CrossRef]
- Emberger, L. Une classification bio-géographique des climats. Rech. Trav. Lab. Bot. Fac. Sci. Montp. Série Bot. 1955, 19, 3–43. [Google Scholar]
- Belaj, A.; León, L.; Satovic, Z.; De la Rosa, R. Variability of wild olives (Olea europaea subsp. europaea var. sylvestris) analyzed by agro—Morphological traits and SSR markers. Sci. Hortic. 2011, 129, 561–569. [Google Scholar]
- Sokal, R.R.; Braumann, C.A. Significance tests for coefficients of variation and variability profiles. Syst. Zool. 1980, 29, 50–66. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenALEx 6.5, genetic analysis in Excel. Population genetic software for teaching and research. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Tessier, C.; David, J.; This, P.; Boursiquot, J.M.; Charrier, A. Optimization of the choice of molecular markers for varietal identification in Vitis vinifera L. Theor. Appl. Genet. 1999, 98, 171–177. [Google Scholar] [CrossRef]
- Jolliffe, I.T.; Cadima, J. Principal component analysis: A review and recent developments. Phil. Trans. R. Soc. A 2016, 374, 2015020220150202. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Saponari, M.; Boscia, D.; Altamura, G.; Loconsole, G.; Zicca, S.; D’Attoma, G.; Morelli, M.; Palmisano, F.; Saponari, A.; Tavano, D.; et al. Isolation and pathogenicity of Xylella fastidiosa associated to the olive quick decline syndrome in southern Italy. Sci. Rep. 2017, 7, 17723. [Google Scholar] [CrossRef]
- Fanelli, V.; Mascio, I.; Falek, W.; Miazzi, M.M.; Montemurro, C. Current Status of Biodiversity Assessment and Conservation of Wild Olive (Olea europaea L. subsp. europaea var. sylvestris). Plants 2022, 11, 480. [Google Scholar]
- Sion, S.; Taranto, F.; Montemurro, C.; Mangini, G.; Camposeo, S.; Falco, V.; Gallo, A.; Mita, G.; Saddoud Debbabi, O.; Ben Amar, F.; et al. Genetic characterization of Apulian olive germplasm as potential source in new breeding programs. Plants 2019, 8, 268. [Google Scholar] [CrossRef] [Green Version]
- Besnard, G.; Baradat, P.; Bervillé, A. Genetic relationships in the olive (Olea europaea L.) reflect multilocal selection of cultivars. Theor. Appl. Genet. 2001, 102, 251–258. [Google Scholar] [CrossRef]
- Breton, C.; Tersac, M.; Bervillé, A. Genetic diversity and gene flow between the wild olive (oleaster, Olea europaea L.) and the olive: Several Plio-Pleistocene refuge zones in the Mediterranean basin suggested by simple sequence repeats analysis. J. Biogeogr. 2006, 33, 1916–1928. [Google Scholar] [CrossRef]
- Kassa, A.; Konrad, H.; Geburek, T. Molecular diversity and gene flow within and among different subspecies of the wild olive (Olea europaea L.): A review. Flora 2019, 250, 18–26. [Google Scholar] [CrossRef]
- Sorkheh, K.; Khaleghi, E. Molecular characterization of genetic variability and structure of olive (Olea europaea L.) germplasm collection analyzed by agromorphological traits and microsatellite markers. Turk. J. Agric. For. 2016, 40, 583–596. [Google Scholar] [CrossRef]
- Koubouris, G.C.; Avramidou, E.; Metzidakis, I.T.; Petrakis, P.V.; Sergentani, C.K.; Doulis, A.G. Phylogenetic and evolutionary applications of analyzing endocarp morphological characters by classification binary tree and leaves by SSR markers for the characterization of olive germplasm. Tree Genet. Genomes 2019, 15, 26. [Google Scholar] [CrossRef]
- Fendri, M.; Trujillo, I.; Trigui, A.; Rodríguez-García, M.; Ramírez, J.D. Simple Sequence Repeat Identification and Endocarp Characterization of Olive Tree Accessions in a Tunisian Germplasm Collection. HortScience 2010, 45, 1429–1436. [Google Scholar] [CrossRef] [Green Version]
- D’Imperio, M.; Viscosi, V.; Scarano, M.T.; D’Andrea, M.; Zullo, B.A.; Pilla, F. Integration between molecular and morphological markers for the exploitation of olive germoplasm (Olea europaea). Sci. Hortic. 2011, 130, 229–240. [Google Scholar] [CrossRef]
- Trujillo, I.; Ojeda, M.A.; Urdiroz, N.M.; Potter, D.; Barranco, D.; Rallo, L.; Diez, C.M. Identification of the worldwide olive germplasm bank of Córdoba (Spain) using SSR and morphological markers. Tree Genet. Genomes 2013, 10, 141–155. [Google Scholar] [CrossRef]
- Hannachi, H.; Breton, C.; Msallem, M.; El Hadj, S.B.; El Gazzah, M.; Bervillé, A. Differences between native and introduced olive cultivars as revealed by morphology of drupes, oil composition and SSR polymorphism: A case study in Tunisia. Sci. Hort. 2008, 116, 280–290. [Google Scholar] [CrossRef]
- Rodrigues, N.; Pinho, T.; Casal, S.; Peres, A.M.; Baptista, P.; Pereira, J.A. Chemical Characterization of Oleaster, Olea europaea var. sylvestris (Mill.) Lehr., Oils from Different Locations of Northeast Portugal. Appl. Sci. 2020, 10, 6414. [Google Scholar] [CrossRef]
- Hannachi, H.; Gómez, J.J.M.; Saadaoui, E.; Cervantes, E. Stone diversity in wild and cultivated olive trees (Olea europaea L.). Dendrobiology 2017, 77, 19–32. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organs | Trait | Trait Code | Classes | |||
|---|---|---|---|---|---|---|
| TREE ARCHITECTURE | Tree Vigor | TV 1 | 1 = Weak | 2 = Medium | 3 = Strong | |
| Tree Habit | TH 1 | 1 = Falling down | 2 = Spread out | 3 = Erected | ||
| Canopy Density | CD 1 | 1 = Loose | 2 = Medium | 3 = Compact | ||
| Tree Production fruit | TP 1 | 1 = Weak | 2 = Medium | 3 = Strong | ||
| Maturation | M 1 | 1 = Early | 2 = Medium | 3 = Late | ||
| LEAF SHAPE AND SIZE | Shape | LSH 3 | 1 = Elliptic (Length/Width < 4) | 2 = Elliptic-lanceolate (Length/Width 4–6) | 3 = Lanceolate (Length/Width > 6) | |
| Blade Length (cm) | LBL 3 | 1 = Short (<5 cm) | 2 = Medium (5–7 cm) | 3 = Long (>7 cm) | ||
| Blade Width (cm) | LBW 3 | 1 = Narrow (<1 cm) | 2 = Medium (1–1.5 cm) | 3 = Wide (>1.50 cm) | ||
| Longitudinal Curvature Blade | LCB 1 | 1 = Epinastic | 2 = Flat | 3 = Hyponastic | 4 = Helicoidal | |
| FRUIT SHAPE AND SIZE | Weight (g) | FW 3 | 1 = Low (<0.5 g) | 2 = Medium (0.5–1 g) | 3 = High (>1 g) | |
| Shape | FSH 3 | 1 = Spherical (L/A < 1.25) | 2 = Oval (Length/Width 1.25–1.45) | 3 = Longer (Length/Width > 1.45) | ||
| Symmetry (position A) | FSA 1 | 1 = Symmetrical | 2 = Slightly Asymmetrical | 3 = Asymmetrical | ||
| Maximal Diameter | FMD 1 | 1 = Toward base | 2 = Central | 3 = Toward apex | ||
| Apex Shape (position A) | FASH 1 | 1 = Pointed | 2 = Rounded | |||
| Base Shape (position A) | FBSH 1 | 1 = Rounded | 2 = Truncate | 3 = Pointed | ||
| Nipple | NI 1 | 1 = Absent | 2 = Tenuous | |||
| Lenticels | LE 1 | 1 = Few | 2 = Many | |||
| Dimension Lenticels | DLE 1 | 1 = Small | 2 = Large | |||
| Length (cm) | FL 2 | / | / | / | ||
| Width (cm) | FWI 2 | / | / | / | ||
| STONE SHAPE AND SIZE | Weight (g) | SW 3 | 1 = Low (<0.15 g) | 2 = Medium (0.15–0.30 g) | 3 = High (>0.3 g) | |
| Shape (position A) | SSH 3 | 1 = Spherical (Length/Width < 1.4) | 2 = Oval (Length/Width 1.4–1.8) | 3 = Elliptic (Length/Width 1.8–2.2) | 4 = Longer (Length/Width > 2.2) | |
| Symmetry (position A) | SSA 1 | 1 = Symmetrical | 2 = Slightly Asymmetrical | 3 = Asymmetrical | ||
| Maximal Diameter | SMD 1 | 1 = Toward base | 2 = Central | 3 = Toward apex | ||
| Groves distribution | SDG 1 | 1 = Regular | 2 = Irregular | |||
| Apex Shape | SASH 1 | 1 = Pointed | 2 = Rounded | |||
| Base Shape | SBSH 1 | 1 = Rounded | 2 = Truncate | 3 = Pointed | ||
| Surface | SS 1 | 1 = Smooth | 2 = Rugose | 3 = Scabrous | ||
| Number of Grooves | SNG 3 | 1 = Reduced (<6) | 2 = Medium (6–7) | 3 = High (>7) | ||
| Mucron | SM 1 | 1 = Without mucron | 2 = With mucron | |||
| Length (cm) | SL 2 | / | / | / | ||
| Width (cm) | SWI 2 | / | / | / | ||
| Trait | Range | Mean | Standard Deviation | CV (%) |
|---|---|---|---|---|
| Leaf Blade Length (cm) | 2.97–8.99 | 5.38 | 0.96 | 17.87% |
| Leaf Blade Width (cm) | 0.56–2.05 | 1.11 | 0.29 | 26.19% |
| Leaf Shape (L/I) | 2.87–9.31 | 5.14 | 1.24 | 24.09% |
| Fruit Weight (g) | 0.11–1.87 | 0.61 | 0.35 | 57.82% |
| Fruit Length (cm) | 0.64–2.19 | 1.22 | 0.24 | 19.81% |
| Fruit Width (cm) | 0.48–1.36 | 0.84 | 0.16 | 19.52% |
| Fruit Shape (L/I) | 1.09–2.13 | 1.48 | 0.18 | 11.86% |
| Stone Weight (g) | 0.04–0.98 | 0.19 | 0.10 | 51.57% |
| Stone Length (cm) | 0.56–1.64 | 1.04 | 0.18 | 17.16% |
| Stone Width (cm) | 0.38–0.81 | 0.56 | 0.07 | 11.70% |
| Stone Shape (L/I) | 1.32–2.93 | 1.87 | 0.26 | 13.93% |
| Stone Number of Grooves (SNG) | 4.18–14.65 | 9.57 | 1.89 | 19.73% |
| Descriptors | LBW | LSH | FW | FL | FWI | FSH | SW | SL | SWl | SSH | SNG |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Leaf Blade Length (cm) (LBL) | 0.57 *** | 0.15 | 0.41 *** | 0.45 *** | 0.42 *** | 0.08 | 0.32 *** | 0.35 *** | 0.22 ** | 0.25 *** | 0.00 |
| Leaf Blade Width (cm) (LBW) | −0.67 *** | 0.36 *** | 0.37 *** | 0.45 *** | −0.14 | 0.33 *** | 0.18 * | 0.26 *** | 0.01 | −0.10 | |
| Leaf Shape (L/I) (LSH) | −0.10 | −0.07 | −0.20 ** | 0.27 *** | −0.11 | 0.08 | −0.13 | 0.21 ** | 0.13 | ||
| Fruit Weight (g) (FW) | 0.87 *** | 0.90 *** | −0.03 | 0.72 *** | 0.69 *** | 0.71 *** | 0.26 *** | 0.05 | |||
| Fruit Length (cm) (FL) | 0.86 *** | 0.30 *** | 0.72 *** | 0.89 *** | 0.69 *** | 0.55 *** | 0.08 | ||||
| Fruit Width (cm) (FWI) | −0.22 ** | 0.71 *** | 0.60 *** | 0.76 *** | 0.12 | 0.03 | |||||
| Fruit Shape (L/I) (FSH) | 0.03 | 0.56 *** | −0.11 | 0.85 *** | 0.11 | ||||||
| Stone Weight (g) (SW) | 0.64 *** | 0.75 *** | 0.17 | 0.14 | |||||||
| Stone Length (cm) (SL) | 0.66 *** | 0.74 *** | 0.09 | ||||||||
| Stone Width (cm) (SW) | 0.00 | 0.04 | |||||||||
| Stone Shape (L/I) (SSH) | 0.07 |
| Trait | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bioclimate | Leaf Blade Length (cm) | Leaf Blade Width (cm) | Leaf Shape (L/I) | Fruit Weight (g) | Fruit Length (cm) | Fruit Width (cm) | Fruit Shape (L/I) | Stone Weight (g) | Stone Length (cm) | Stone Width (cm) | Stone Shape (L/I) | Stone Number of Grooves |
| Sub-humid | 5.25 ± 0.83 a | 1.09 ± 0.26 a | 5.13 ± 1.37 a,b | 0.59 ± 0.31 a | 1.22 ± 0.21 a | 0.85 ± 0.16 a | 1.46 ± 0.17 a,b | 0.22 ± 0.16 a | 1.05 ± 0.17 a | 0.58 ± 0.07 a | 1.84 ± 0.23 a | 9.71 ± 2.08 a |
| Humid | 5.26 ± 0.87 a | 1.09 ± 0.30 a | 5.12 ± 1.38 a,b | 0.61 ± 0.34 a | 1.20 ± 0.22 a | 0.83 ± 0.16 a | 1.47 ± 0.17 a,b | 0.18 ± 0.07 a | 1.01 ± 0.16 a | 0.54 ± 0.06 a | 1.87 ± 0.25 a | 9.55 ± 1.71 a |
| Semi-arid | 5.56 ± 1.10 a | 1.16 ± 0.27 a | 4.93 ± 0.99 a,b | 0.57 ± 0.35 a | 0.57 ± 0.26 a | 0.81 ± 0.15 a | 1.48 ± 0.17 a,b | 0.18 ± 0.08 a | 1.04 ± 0.20 a | 0.56 ± 0.07 a | 1.86 ± 0.30 a | 9.42 ± 2.16 a |
| Arid | 5.34 ± 0.91 a | 0.99 ± 0.28 a | 5.74 ± 1.06 a | 0.69 ± 0.45 a | 1.30 ± 0.30 a | 0.85 ± 0.20 a | 1.55 ± 0.20 a | 0.19 ± 0.07 a | 1.10 ± 0.18 a | 0.55 ± 0.06 a | 1.98 ± 0.26 a | 9.77 ± 1.59 a |
| Saharan | 6.64 ± 1.25 b | 1.56 ± 0.27 b | 4.49 ± 0.48 b | 0.89 ± 0.28 a | 1.41 ± 0.17 a | 1.04 ± 0.12 b | 1.37 ± 0.19 b | 0.25 ± 0.09 a | 1.05 ± 0.21 a | 0.60 ± 0.05 a | 1.74 ± 0.21 a | 9.11 ± 1.38 a |
| Trait | Trait Code | KT | Kj | Hj | Dj |
|---|---|---|---|---|---|
| Tree Vigor | TV | 3 | 3 | 0.979 | 0.582 |
| Tree Habit | TH | 3 | 3 | 1.071 | 0.649 |
| Canopy Density | CD | 3 | 3 | 0.682 | 0.390 |
| Tree Production fruit | TP | 3 | 3 | 0.881 | 0.511 |
| Maturation | M | 3 | 3 | 0.635 | 0.346 |
| Leaf Shape | LSH | 3 | 3 | 0.868 | 0.501 |
| Leaf Blade Length (cm) | LBL | 3 | 3 | 0.841 | 0.513 |
| Leaf Blade Width (cm) | LBW | 3 | 3 | 0.920 | 0.566 |
| Longitudinal Curvature Blade | LCB | 4 | 4 | 1.281 | 0.701 |
| Fruit Weight (g) | FW | 3 | 3 | 0.988 | 0.601 |
| Fruit Shape | FSH | 3 | 3 | 0.910 | 0.566 |
| Fruit Symmetry (position A) | FSA | 3 | 3 | 1.066 | 0.646 |
| Fruit Maximal Diameter | FMD | 3 | 3 | 0.956 | 0.565 |
| Fruit Apex Shape (position A) | FASH | 2 | 2 | 0.693 | 0.500 |
| Fruit Base Shape (position A) | FBSH | 3 | 3 | 1.021 | 0.614 |
| Nipple | NI | 2 | 1 | - | - |
| Lenticels | LE | 2 | 1 | - | - |
| Dimension Lenticels | DLE | 2 | 1 | - | - |
| Stone Weight (g) | SW | 3 | 3 | 0.979 | 0.594 |
| Stone Shape (position A) | SSH | 4 | 4 | 1.046 | 0.609 |
| Stone Symmetry (position A) | SSA | 3 | 3 | 1.063 | 0.645 |
| Stone Maximal Diameter | SMD | 3 | 3 | 1.034 | 0.627 |
| Stone Distribution Grooves | SDG | 2 | 2 | 0.682 | 0.489 |
| Stone Apex Shape | SASH | 2 | 2 | 0.680 | 0.487 |
| Stone Base Shape | SBSH | 3 | 3 | 0.795 | 0.448 |
| Stone Surface | SS | 3 | 3 | 1.081 | 0.655 |
| Stone Number of Grooves | SNG | 3 | 3 | 0.366 | 0.175 |
| Stone Mucron | SM | 2 | 2 | 0.691 | 0.498 |
| Average | 0.89 | 0.54 | |||
| Min | 0.37 | 0.18 | |||
| Max | 1.28 | 0.70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falek, W.; Mascio, I.; Gadaleta, S.; Fanelli, V.; Bechkri, S.; Khelifi, D.; Miazzi, M.M.; Montemurro, C. Morphological and Eco-Geographic Variation in Algerian Wild Olives. Plants 2022, 11, 1803. https://doi.org/10.3390/plants11141803
Falek W, Mascio I, Gadaleta S, Fanelli V, Bechkri S, Khelifi D, Miazzi MM, Montemurro C. Morphological and Eco-Geographic Variation in Algerian Wild Olives. Plants. 2022; 11(14):1803. https://doi.org/10.3390/plants11141803
Chicago/Turabian StyleFalek, Wahiba, Isabella Mascio, Susanna Gadaleta, Valentina Fanelli, Sakina Bechkri, Douadi Khelifi, Monica Marilena Miazzi, and Cinzia Montemurro. 2022. "Morphological and Eco-Geographic Variation in Algerian Wild Olives" Plants 11, no. 14: 1803. https://doi.org/10.3390/plants11141803
APA StyleFalek, W., Mascio, I., Gadaleta, S., Fanelli, V., Bechkri, S., Khelifi, D., Miazzi, M. M., & Montemurro, C. (2022). Morphological and Eco-Geographic Variation in Algerian Wild Olives. Plants, 11(14), 1803. https://doi.org/10.3390/plants11141803