
Effects of Low-Temperature Stress during the Anther Differentiation Period on Winter Wheat Photosynthetic Performance and Spike-Setting Characteristics


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Plant Measurement and Methods
2.3. Plant Sampling
2.4. Data Processing
3. Results
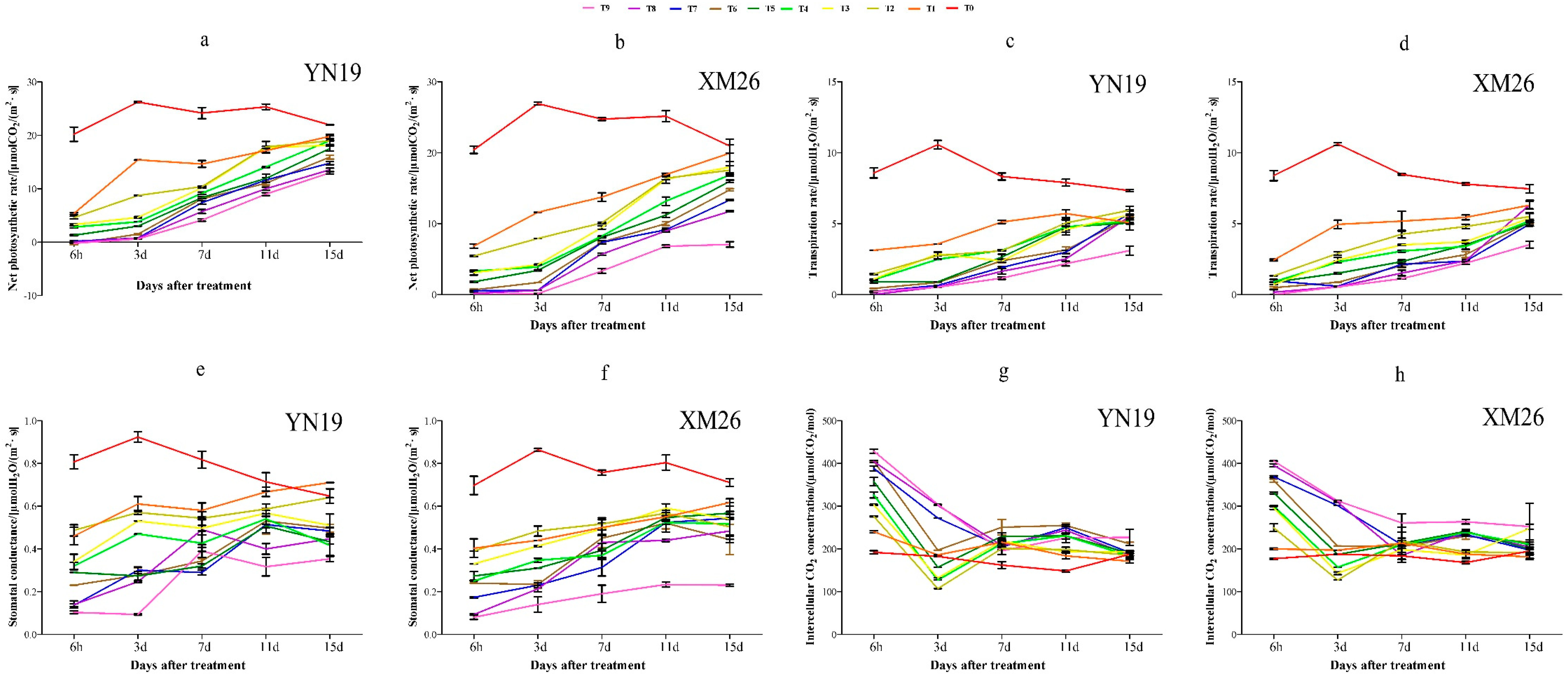
3.1. Photosynthetic Parameters of Wheat
3.1.1. Photosynthetic Parameters Relative to Variety, Treatment Temperature and Time
3.1.2. Changes to Photosynthetic Parameters of Wheat after Treatment
Net Photosynthetic Rate
Transpiration Rate
Stomatal Conductance
Intercellular CO2 Concentration
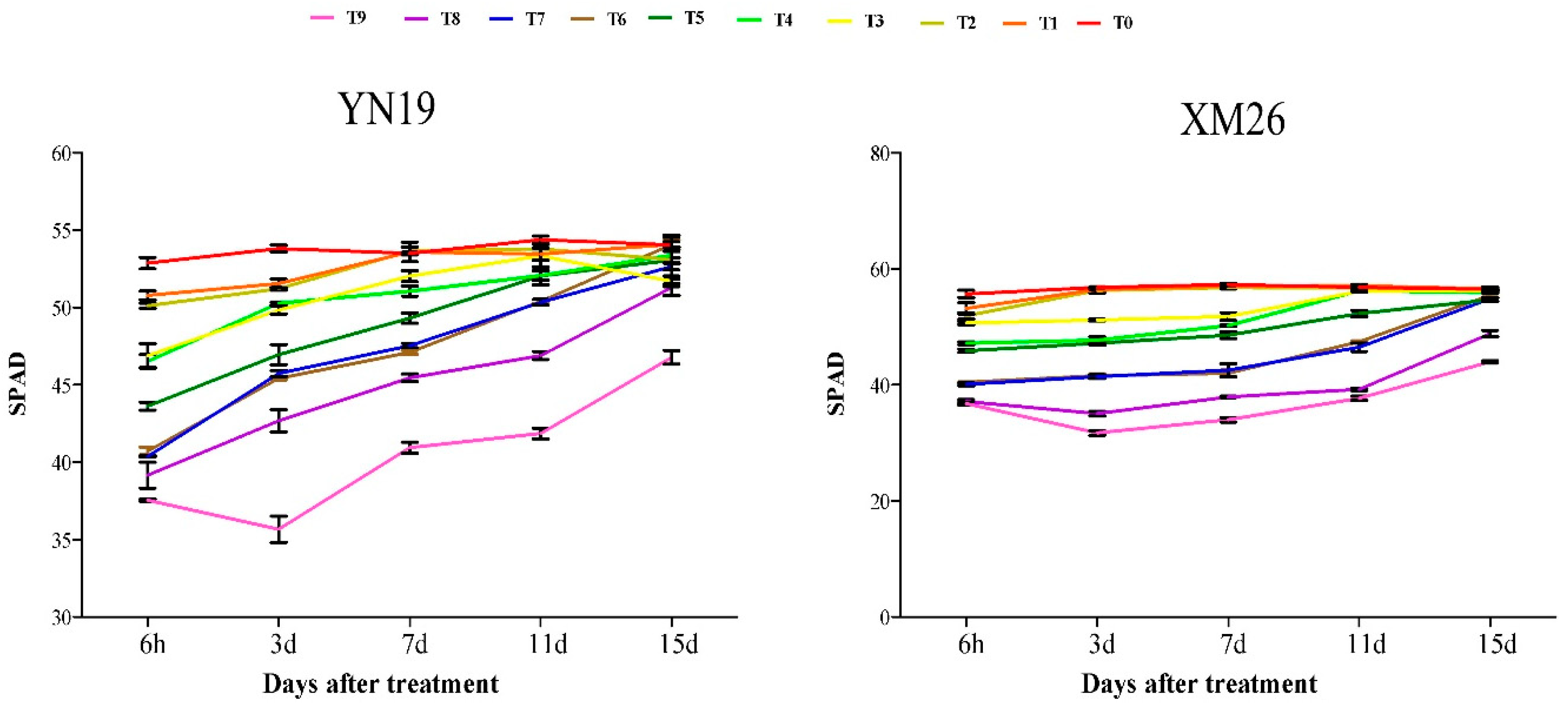
3.2. Relative Chlorophyll Content (SPAD) of Wheat
3.3. Wheat Spike-Setting Characteristics
3.3.1. Spike-Setting Characteristics and Yield Relative to Variety, Temperature Treatment, Treatment Time
3.3.2. Changes of Spike-Setting Characteristics
3.4. The Relationship between Photosynthetic Parameters and Wheat Spike-Setting Characteristics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bruinsma, J. The Resource Outlook to 2050. By How Much Do Land, Water Use and Crop Yields Need to Increase by 2050; Technical Papers from the Expert Meeting on How to Feed the World in 2050; FAO: Rome, Italy, 2009; pp. 1–33. [Google Scholar]
- FAO. The Future of Food and Agriculture—Trends and Challenges; FAO: Rome, Italy, 2017. [Google Scholar]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield Trends Are Insufficient to Double Global Crop Production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Special Report. In Global Warming of 1.5 °C; The Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2018.
- Barlow, K.M.; Christy, B.P.; O’Leary, G.J.; Riffkin, P.A.; Nuttall, J.G. Simulating the impact of extreme heat and frost events on wheat crop production: A review. Field Crop. Res. 2015, 171, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Tshewang, S.; Jessop, R.; Birchall, C. Effect of frost on triticale and wheat varieties at flowering in the north eastern Australian cereal belt. Cereal Res. Commun. 2017, 45, 655–664. [Google Scholar] [CrossRef]
- Crimp, S.J.; Zheng, B.; Khimashia, N.; Gobbett, D.L.; Chapman, S.; Howden, M.; Nicholls, N. Recent changes in southern Australian frost occurrence: Implications for wheat production risk. Crop Pasture Sci. 2016, 67, 801–811. [Google Scholar] [CrossRef]
- Frederiks, T.M.; Christopher, J.T.; Borrel, A.K. Investigation of Post Head-Emergence Frost Resistance in Several CIMMYT Synthetic and Queensland wheats. In Proceedings of the 4th International Crop Science Congress, Brisbane, Australia, 26 September–1 October 2004. [Google Scholar]
- Frederiks, T.M.; Christopher, J.T.; Borrell, A.K. Low Temperature Adaption of Wheat Post Head-Emergence in Northern Australia. In Proceedings of the 11th International Wheat Genetics Symposium, Brisbane, Australia, 24–29 August 2008; Sidney University Press: Sydney, Australia. [Google Scholar]
- Gu, L.; Hanson, P.J.; Post, W.M.; Kaiser, D.P.; Yang, B.; Nemani, R.; Pallardy, S.G.; Meyers, T. The 2007 Eastern US Spring Freeze: Increased Cold Damage in a Warming World? Bio Sci. 2008, 58, 253–262. [Google Scholar] [CrossRef]
- Trnka, M.; Rötter, R.; Ruiz-Ramos, M.; Kersebaum, K.C.; Olesen, J.E.; Žalud, Z.; Semenov, M. Adverse weather conditions for European wheat production will become more frequent with climate change. Nat. Clim. Chang. 2014, 4, 637–643. [Google Scholar] [CrossRef]
- Zhong, X.; Mei, X.; Li, Y.; Yoshida, H.; Zhao, P.; Wang, X.; Han, L.; Hu, X.; Huang, S.; Huang, J.; et al. Changes in Frost Resistance of Wheat Young Ears with Development During Jointing Stage. J. Agron. Crop Sci. 2008, 194, 343–349. [Google Scholar] [CrossRef]
- Holman, J.D.; Schlegel, A.J.; Thompson, C.R.; Lingenfelser, J.E. Influence of Precipitation, Temperature, and 56 Years on Winter Wheat Yields in Western Kansas. Crop Manag. 2011, 10, 11–21. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, X.C.; Hu, W.G.; Cao, T.; Liu, Z.; Chen, Y. Status and suggestion of wheat variety utilization in southern Huang-Huai wheat region. J. Seed Ind. Guide 2016, 45, 18–38. [Google Scholar] [CrossRef]
- Gao, Y.; Tang, J.W.; Yin, G.H.; Hart, Y.L.; Huang, F.; Wang, L.N.; Yu, H.F.; Li, N.N.; Zhang, Q.; Yang, G.Y.; et al. Effect of Different Periods and Frequency of Late Spring Coldness on Winter Wheat Yield Related Traits. J. Triticeae Crop. 2015, 35, 687–692. [Google Scholar] [CrossRef]
- Qian, Y.L.; Wang, J.L.; Zheng, C.L. Spatial-temporal change of low temperature disaster of winter wheat in North China in last 50 years. Chin. J. Ecol. 2014, 33, 3245–3253. [Google Scholar] [CrossRef]
- Zhang, W.J.; Huang, Z.L.; Wang, Q.; Guan, Y.N. Effects of Low Temperature on Leaf Anatomy and Photosynthetic Performance in Different Genotypes of Wheat Following a Rice Crop. Int. J. Agric. Biol. 2015, 17, 1165–1171. [Google Scholar] [CrossRef]
- Wang, W.; Wang, X.; Zhang, J.; Huang, M.; Cai, J.; Zhou, Q.; Dai, T.; Jiang, D. Salicylic acid and cold priming induce late-spring freezing tolerance by maintaining cellular redox homeostasis and protecting photosynthetic apparatus in wheat. Plant Growth Regul. 2019, 90, 109–121. [Google Scholar] [CrossRef]
- Thakur, P.; Kumar, S.; Malik, J.A.; Berger, J.D.; Nayyar, H. Cold stress effects on reproductive development in grain crops: An overview. Environ. Exp. Bot. 2010, 67, 429–443. [Google Scholar] [CrossRef]
- Li, C.-Y.; Xu, W.; Liu, L.-W.; Yang, J.; Zhu, X.-K.; Guo, W.-S. Changes of endogenous hormone contents and antioxidative enzyme activities in wheat leaves under low temperature stress at jointing stage. Chin. J. Appl. Ecol. 2015, 26, 2015–2022. [Google Scholar] [CrossRef]
- Li, X.; Topbjerg, H.B.; Jiang, D.; Liu, F. Drought priming at vegetative stage improves the antioxidant capacity and photosynthesis performance of wheat exposed to a short-term low temperature stress at jointing stage. Plant Soil 2015, 393, 307–318. [Google Scholar] [CrossRef]
- Zheng, J.; Liu, T.; Zheng, Q.; Li, J.; Qian, Y.; Li, J.; Zhan, Q. Identification of cold tolerance and analysis of genetic diversity for major wheat varieties in Jianghuai region of China. Pak. J. Bot. 2019, 52, 839–849. [Google Scholar] [CrossRef]
- Wang, R.-X.; Yan, C.-S.; Zhang, X.-Y.; Sun, G.-Z.; Qian, Z.-G.; Qi, X.-L.; Mou, Q.-H.; Xiao, S.-H. Effect of Low Temperature in Spring on Yield and Photosynthetic Characteristics of Wheat. Acta Agron. Sin. 2018, 44, 288–296. [Google Scholar] [CrossRef]
- Tao, H.Z.; Zhao, C.L.; Li, W.Q. Photosynthetic Response to Low Temperature in Plant. Chin. J. Biochem. Mol. Biol. 2012, 28, 501–508. [Google Scholar] [CrossRef]
- Xu, C.C.; Zhang, J.H. Effect of drought on chlorophyll fluorescence and xanthophylls cycle components in winter wheat leaves with different ages. Acta Photophysiol. Sin. 1999, 25, 29–37. [Google Scholar] [CrossRef]
- Ge, J.; Jiang, X.J.; Ren, D.C.; Chen, S.D. Effects of Low Temperature Stress on Antioxidant System and Photosynthetic Capacity of Wheat at Jointing Stage. Tianjin Agric. Sci. 2021, 27, 5–9. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.Y.; Qian, J.W.; Liang, C.D. Characteristics and main cultivation techniques of Yannong 19 wheat variety in Xuhuai Area of Jiangsu Province. J. Triticeae Crops 2003, 23, 96. [Google Scholar] [CrossRef]
- Notice of the Ministry of Agriculture of People’s Republic of China No. 1505; Ministry of Agriculture of the PRC: Beijing, China, 2010.
- Li, X.; Pu, H.; Liu, F.; Zhou, Q.; Cai, J.; Dai, T.; Cao, W.; Jiang, D. Winter Wheat Photosynthesis and Grain Yield Responses to Spring Freeze. Agron. J. 2015, 107, 1002–1010. [Google Scholar] [CrossRef]
- Fan, Q.H.; Sun, W.C.; Li, Z.J.; Liang, Y.C. Effects of silicon on photosynthesis and its major relevant enzyme activities in wheat leaves under short-term cold stress. Plant Nutr. Fertil. Sci. 2009, 15, 544–550. [Google Scholar] [CrossRef]
- Guan, Y.-N.; Huang, Z.-L.; Zhang, W.-J.; Shi, X.-D.; Zhang, P.-P. Effects of low temperature stress on photosynthetic performance of different genotypes wheat cultivars. Chin. J. Appl. Ecol. 2013, 24, 1895–1899. [Google Scholar] [CrossRef]
- Marcellos, H. Wheat frost injury-freezing stress and photosynthesis. Aust. J. Agric. Res. 1977, 28, 557–564. [Google Scholar] [CrossRef]
- Fuller, M.; Fuller, A.M.; Kaniouras, S.; Christophers, J.; Fredericks, T. The freezing characteristics of wheat at ear emergence. Eur. J. Agron. 2007, 26, 435–441. [Google Scholar] [CrossRef]
- Cui, B.; Cheng, S.L.; Yuan, X.Y.; Zhou, Y.R.; Hao, P.A.; Li, J.L.; Ma, J. Effects of Low Temperature Stress on the Photosynthetic Characteristics and Chlorophyll Fluorescence Parameters of Bletilla striata. Chin. J. Trop. Crop. 2019, 40, 891–897. [Google Scholar] [CrossRef]
- Guo, Z.-J.; Yu, Z.-W.; Shi, Y.; Zhao, J.-Y.; Zhang, Y.-L.; Wang, D. Photosynthesis Characteristics of Flag Leaf and Dry Matter Accumulation and Allocation in Winter Wheat under Supplemental Irrigation after Measuring Moisture Content in Different Soil Layers. Acta Agron. Sin. 2014, 40, 731–738. [Google Scholar] [CrossRef]
- Yuanyuan, K.; Xiang, C.; Lele, Z.; Yan, Z.; Hui, X.; Hassan, M.A.; Gang, J.; Feifei, L.; Fengzhen, W.; Jincai, L. Effects of low temperature stress at another connective stage on dry matter translocation and distribution and grain yield of wheat. J. Anhui Agric. Univ. 2021, 48, 701–706. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, Y.; Ma, Q.; Su, S.; Li, C.; Ding, J.; Zhu, M.; Zhu, X.; Guo, W. Effects of low temperature in spring on fertility of pollen and formation of grain number in wheat. Acta Agron. Sin. 2020, 47, 104–115. [Google Scholar] [CrossRef]
- Xiao, L.; Liu, B.; Zhang, H.; Gu, J.; Fu, T.; Asseng, S.; Liu, L.; Tang, L.; Cao, W.; Zhu, Y. Modeling the response of winter wheat phenology to low temperature stress at elongation and booting stages. Agric. For. Meteorol. 2021, 303, 108376. [Google Scholar] [CrossRef]
- Ji, H.; Xiao, L.; Xia, Y.; Song, H.; Liu, B.; Tang, L.; Cao, W.; Zhu, Y.; Liu, L. Effects of jointing and booting low temperature stresses on grain yield and yield components in wheat. Agric. For. Meteorol. 2017, 243, 33–42. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, G.; Gao, X.; Chu, C. Photosynthetic decline in flag leaves of two field-grown spring wheat cultivars with different senescence properties. S. Afr. J. Bot. 2006, 72, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Slafer, G.; Abeledo, L.; Miralles, D.; González, F.G.; Whitechurch, E. Photoperiod sensitivity during stem elongation as an avenue to raise potential yield in wheat. Euphytica 2001, 119, 191–197. [Google Scholar] [CrossRef]
- Pan, Y.H. Effects of Potassium Deficiency on Leaf Area Establishment and Photosynthesis Characteristics of Brassica napus. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2017. [Google Scholar]
- Li, W.M.; Zhang, J.B.; Zhu, A.N. Effects of air temperature and humidity on the photosynthesis of winter wheat. J. Irrig. Drain. 2008, 27, 90–92. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cultivar | Breeding Unit | Certification Unit | Information |
|---|---|---|---|
| Yannong 19 | Yantai Academy of Agricultural Sciences, Shandong Province, China | Shandong Provincial Crop Variety Certification Committee, China | Medium-gluten winter wheat variety. The main wheat variety planted in Anhui Province [27]. |
| Xinmai 26 | Xinxiang Academy of Agricultural Sciences, Henan Province, China | China National Crop Variety Examination and Approving Committee | Super stringy semi-winter wheat variety. Suitable for planting in the south of Huang-huai winter wheat area, China. Attention should be paid the pre-vention of spring low-temperature stress after sowing [28]. |
| Treatment | Day/Night Temperature (°C) | Day/Night Time (h) |
|---|---|---|
| T0(CK) | 15/15 | |
| T1 | 15/2 | 12/12 (1 day) |
| T2 | 15/2 | 12/12 (2 days) |
| T3 | 15/2 | 12/12 (3 days) |
| T4 | 15/0 | 12/12 (1 day) |
| T5 | 15/0 | 12/12 (2 days) |
| T6 | 15/0 | 12/12 (3 days) |
| T7 | 15/−2 | 12/12 (1 day) |
| T8 | 15/−2 | 12/12 (2 days) |
| T9 | 15/−2 | 12/12 (3 days) |
| Pn | Tr | ||||||||||
| Source | df | 6 h | 3 days | 7 days | 11 days | 15 days | 6 h | 3 days | 7 days | 11 days | 15 days |
| Variety (V) | 1 | 5.061 * | 24.679 ** | 1.742 | 23.700 ** | 0.702 | 1.754 | 5.150 * | 4.459 * | 26.250 ** | 212.654 ** |
| Time (T) | 2 | 107.372 ** | 4817.742 ** | 180.756 ** | 272.750 ** | 71.056 ** | 72.772 ** | 400.596 ** | 144.267 ** | 148.309 ** | 64.425 ** |
| TEMP | 3 | 2094.333 ** | 46,721.242 ** | 1876.492 ** | 919.008 ** | 143.510 ** | 2689.090 ** | 5683.113 ** | 1180.115 ** | 593.546 ** | 396.052 ** |
| V × T | 2 | 0.294 | 67.738 ** | 0.211 | 1.319 | 14.917 ** | 3.563 * | 1.841 | 7.468 ** | 3.597 * | 17.443 ** |
| V × TEMP | 3 | 0.385 | 58.008 ** | 2.474 | 1.95 | 2.034 | 0.455 | 3.651 * | 0.238 | 2.51 | 45.499 ** |
| T × TEMP | 6 | 14.837 ** | 970.723 ** | 27.565 ** | 34.058 ** | 8.570 ** | 16.031 ** | 58.279 ** | 20.988 ** | 19.242 ** | 10.964 ** |
| V × T × TEMP | 6 | 0.491 | 30.416 ** | 0.155 | 0.738 | 2.770 * | 1.919 | 6.823 ** | 2.494 * | 2.128 | 15.581 ** |
| Gs | Ci | ||||||||||
| Source | df | 6 h | 3 days | 7 days | 11 days | 15 days | 6 h | 3 days | 7 days | 11 days | 15 days |
| Varieties (V) | 1 | 23.092 ** | 256.971 ** | 6.766 * | 0.79 | 0.564 | 217.686 ** | 1974.806 ** | 0.562 | 4.692 * | 6.934 * |
| Time (T) | 2 | 134.894 ** | 200.395 ** | 36.961 ** | 39.917 ** | 20.439 ** | 1590.326 ** | 48,057.871 ** | 0.989 | 114.223 ** | 1.196 |
| TEMP | 3 | 553.162 ** | 719.595 ** | 205.825 ** | 105.302 ** | 42.544 ** | 2772.767 ** | 1652.910 ** | 19.683 ** | 195.804 ** | 8.332 ** |
| V × T | 2 | 0.834 | 7.642 ** | 4.161 * | 0.521 | 2.979 | 4.764 * | 47.854 ** | 2.829 | 0.75 | 0.164 |
| V × TEMP | 3 | 4.410 ** | 12.159 ** | 0.782 | 5.186 ** | 2.123 | 3.665 * | 166.138 ** | 1.728 | 4.434 ** | 0.576 |
| T × TEMP | 6 | 15.926 ** | 23.716 ** | 9.296 ** | 10.376 ** | 5.653 ** | 188.097 ** | 7786.114 ** | 1.13 | 13.244 ** | 0.669 |
| V × T × TEMP | 6 | 1.065 | 4.407 ** | 3.918 ** | 1.288 | 2.851 * | 5.384 ** | 50.951 ** | 3.708 ** | 6.360 ** | 0.643 |
| Source | df | Grains per Spike | Grains per Upper Spikelet | Grains per Middle Spikelet | Grains per Lower Spikelet | Yield per Plant (g) | 1000−Grain Weight | Upper Spikelet Yield (g) | Middle Spikelet Yield (g) | Lower Spikelet Yield (g) |
|---|---|---|---|---|---|---|---|---|---|---|
| Variety (V) | 1 | 5.646 * | 5.608 * | 481.909 ** | 3.354 | 51.246 ** | 1645.898 ** | 6.806 * | 0.515 | 2.472 |
| Time (T) | 2 | 187.029 ** | 136.636 ** | 63.950 ** | 7.805 ** | 56.403 ** | 2729.459 ** | 60.831 ** | 153.725 ** | 26.966 ** |
| TEMP | 3 | 1834.842 ** | 1203.659 ** | 610.441 ** | 339.778 ** | 2275.089 ** | 1100.179 ** | 261.209 ** | 537.149 ** | 1889.350 ** |
| V × T | 2 | 6.557 ** | 10.399 ** | 10.297 ** | 0.094 | 7.921 ** | 1117.622 ** | 4.474 * | 6.626 ** | 1.131 |
| V × TEMP | 3 | 2.804 * | 2.880 * | 1.177 | 1.925 | 12.260 ** | 90.969 ** | 3.254 * | 6.026 ** | 8.190 ** |
| T × TEMP | 6 | 24.301 ** | 16.326 ** | 7.532 ** | 4.590 ** | 7.963 ** | 346.694 ** | 8.733 ** | 18.726 ** | 5.529 ** |
| V × T × TEMP | 6 | 1.533 | 1.319 | 1.652 | 0.491 | 1.264 | 154.425 ** | 1.038 | 1.287 | 0.262 |
| Variety | Treatment | Grains per Spike | Grain−Setting Loss Rate | Grains per Upper Spikelet | Upper Spikelet Grain−Setting Loss Rate | Grains per Middle Spikelet | Middle Spikelet Grain−Setting Loss Rate | Grains per Lower Spikelet | Lower Spikelet Grain−Setting Loss Rate |
|---|---|---|---|---|---|---|---|---|---|
| YN19 | 72 h −2 °C | 3.40 ± 1.00 e | 52.02% | 0.00 ± 0.00 g | 100.00% | 1.40 ± 1.14 g | 92.55% | 2.00 ± 2.35 bcd | 80.00% |
| 72 h 0 °C | 4.00 ± 0.00 e | 51.26% | 0.00 ± 0.00 g | 100.00% | 4.00 ± 0.71 f | 78.72% | 0.00 ± 0.00 f | 100.00% | |
| 72 h 2 °C | 4.20 ± 0.58 e | 51.01% | 0.00 ± 0.00 g | 100.00% | 4.20 ± 1.63 f | 77.66% | 0.00 ± 0.00 f | 100.00% | |
| 48 h −2 °C | 9.60 ± 0.58 d | 44.19% | 0.00 ± 0.00 g | 100.00% | 7.80 ± 2.95 cde | 58.51% | 0.60 ± 0.55 def | 94.00% | |
| 48 h 0 °C | 9.20 ± 0.00 d | 44.70% | 1.20 ± 0.84 fg | 91.30% | 7.40 ± 0.55 de | 60.64% | 1.80 ± 1.64 bcdef | 82.00% | |
| 48 h 2 °C | 13.80 ± 0.58 c | 38.89% | 1.40 ± 0.89 fg | 89.86% | 10.00 ± 1.22 c | 46.81% | 2.40 ± 2.51 bc | 76.00% | |
| 24 h −2 °C | 14.80 ± 1.00 c | 37.63% | 4.60 ± 1.52 cd | 66.67% | 9.00 ± 0.71 cde | 52.13% | 1.20 ± 1.1 bcdef | 88.00% | |
| 24 h 0 °C | 20.20 ± 1.00 b | 30.81% | 5.60 ± 1.52 bc | 59.42% | 12.00 ± 1.41 b | 36.17% | 2.60 ± 1.52 b | 74.00% | |
| 24 h 2 °C | 21.40 ± 0.58 b | 29.29% | 6.60 ± 1.34 b | 52.17% | 12.40 ± 0.89 b | 34.04% | 2.40 ± 0.89 bc | 76.00% | |
| ck | 44.60 ± 0.58 a | 13.80 ± 0.84 a | 18.80 ± 1.30 a | 10.00 ± 1.87 a | |||||
| XM26 | 72 h −2 °C | 3.00 ± 0.00 f | 53.79% | 0.00 ± 0.00 g | 100.00% | 3.00 ± 0.71 fg | 84.38% | 0.00 ± 0.00 f | 100.00% |
| 72 h 0 °C | 4.00 ± 1.00 f | 52.53% | 0.00 ± 0.00 g | 100.00% | 4.00 ± 1.00 f | 79.17% | 0.00 ± 0.00 f | 100.00% | |
| 72 h 2 °C | 4.00 ± 1.00 f | 52.53% | 0.00 ± 0.00 g | 100.00% | 4.00 ± 1.00 f | 79.17% | 0.00 ± 0.00 f | 100.00% | |
| 48 h −2 °C | 8.00 ± 1.00 e | 47.47% | 0.60 ± 1.34 fg | 95.77% | 7.20 ± 3.70 e | 62.50% | 0.20 ± 0.45 ef | 97.96% | |
| 48 h 0 °C | 9.80 ± 0.58 de | 45.20% | 1.20 ± 0.84 fg | 91.55% | 7.80 ± 0.83 cde | 59.38% | 0.80 ± 0.84 cdef | 91.84% | |
| 48 h 2 °C | 14.00 ± 0.58 bc | 39.90% | 1.20 ± 0.84 fg | 91.55% | 10.00 ± 1.58 c | 47.92% | 2.80 ± 0.84 b | 71.43% | |
| 24 h −2 °C | 12.40 ± 1.00 cd | 41.92% | 2.80 ± 1.1 e | 80.28% | 9.00 ± 1.41 cde | 53.13% | 0.60 ± 0.89 def | 93.88% | |
| 24 h 0 °C | 15.80 ± 0.58 b | 37.63% | 3.60 ± 0.89 de | 74.65% | 9.60 ± 1.34 cd | 50.00% | 1.40 ± 0.55 bcdef | 85.71% | |
| 24 h 2 °C | 15.00 ± 0.58 bc | 38.64% | 3.80 ± 1.3 de | 73.24% | 9.80 ± 2.28 c | 48.96% | 2.60 ± 1.14 b | 73.47% | |
| ck | 45.60 ± 0.58 a | 14.20 ± 1.3 a | 19.20 ± 1.10 a | 9.80 ± 1.79 a |
| Variety | Treatment | 1000−Grain Weight (g) | Yield per Plant (g) | Yield per Plant Yield Reduction Rate (%) | Upper Spikelets Yield (g) | Middle Spikelets Yield (g) | Lower Spikelets Yield (g) | Upper Spikelets Yield Reduction Rate (%) | Middle Spikelets Yield Reduction Rate (%) | Lower Spikelets Yield Reduction Rate (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| YN19 | 72 h −2 °C | 35.16 ± 0.30 h | 0.05 ± 0.08 h | 99.53 | 0.00 ± 0.00 f | 0.05 ± 0.04 i | 0.07 ± 0.08 fg | 100.00 | 94.10 | 84.00 |
| 72 h 0 °C | 35.97 ± 0.64 g | 0.07 ± 0.13 h | 99.44 | 0.00 ± 0.00 f | 0.14 ± 0.02 h | 0.00 ± 0.00 g | 100.00 | 82.77 | 100.00 | |
| 72 h 2 °C | 36.01 ± 0.00 g | 0.20 ± 0.09 h | 98.22 | 0.00 ± 0.00 f | 0.15 ± 0.05 h | 0.00 ± 0.00 g | 100.00 | 81.88 | 100.00 | |
| 48 h −2 °C | 39.60 ± 0.15 f | 0.45 ± 0.12 gh | 92.54 | 0.00 ± 0.00 f | 0.31 ± 0.1 g | 0.05 ± 0.04 fg | 100.00 | 63.00 | 88.14 | |
| 48 h 0 °C | 43.42 ± 0.15 e | 0.41 ± 0.03 gh | 90.59 | 0.03 ± 0.02 def | 0.32 ± 0.02 fg | 0.07 ± 0.07 fg | 95.70 | 61.51 | 83.78 | |
| 48 h 2 °C | 45.00 ± 0.01 c | 1.08 ± 0.22 def | 80.50 | 0.06 ± 0.04 d | 0.45 ± 0.05 d | 0.11 ± 0.11 ef | 89.61 | 46.09 | 75.42 | |
| 24 h −2 °C | 45.52 ± 0.30 b | 1.15 ± 0.50 def | 84.13 | 0.06 ± 0.04 de | 0.41 ± 0.03 def | 0.21 ± 0.07 cd | 90.99 | 50.93 | 52.34 | |
| 24 h 0 °C | 46.99 ± 0.01 a | 1.43 ± 0.39 cd | 66.47 | 0.12 ± 0.06 c | 0.56 ± 0.06 c | 0.26 ± 0.07 bc | 79.85 | 32.45 | 40.11 | |
| 24 h 2 °C | 47.05 ± 0.06 a | 1.79 ± 0.10 c | 60.48 | 0.11 ± 0.04 c | 0.58 ± 0.04 c | 0.31 ± 0.06 b | 81.38 | 30.12 | 29.33 | |
| ck | 43.94 ± 0.09 d | 6.79 ± 0.33 a | 0.61 ± 0.03 b | 0.83 ± 0.05 b | 0.44 ± 0.08 a | |||||
| XM26 | 72 h −2 °C | 41.41 ± 0.58 h | 0.05 ± 0.09 h | 99.59 | 0.00 ± 0.00 f | 0.12 ± 0.03 hi | 0.00 ± 0.00 g | 100.00 | 86.40 | 100.00 |
| 72 h 0 °C | 43.00 ± 0.00 f | 0.05 ± 0.09 h | 99.43 | 0.00 ± 0.00 f | 0.17 ± 0.04 h | 0.00 ± 0.00 g | 100.00 | 81.17 | 100.00 | |
| 72 h 2 °C | 42.00 ± 0.06 g | 0.17 ± 0.147 h | 98.89 | 0.00 ± 0.00 f | 0.17 ± 0.04 h | 0.00 ± 0.00 g | 100.00 | 81.61 | 100.00 | |
| 48 h −2 °C | 42.70 ± 0.46 f | 0.34 ± 0.32 gh | 97.75 | 0.01 ± 0.02 ef | 0.31 ± 0.14 g | 0.03 ± 0.06 g | 98.74 | 66.35 | 94.51 | |
| 48 h 0 °C | 43.42 ± 0.15 e | 0.70 ± 0.39 fg | 92.99 | 0.04 ± 0.03 def | 0.34 ± 0.32 efg | 0.05 ± 0.04 fg | 94.86 | 62.92 | 88.82 | |
| 48 h 2 °C | 44.63 ± 0.12 c | 0.96 ± 0.56 def | 87.66 | 0.05 ± 0.03 def | 0.45 ± 0.06 d | 0.13 ± 0.04 ef | 92.07 | 51.14 | 73.20 | |
| 24 h −2 °C | 43.63 ± 0.00 e | 0.39 ± 0.35 gh | 92.88 | 0.03 ± 0.03 def | 0.39 ± 0.05 defg | 0.12 ± 0.05 ef | 96.13 | 57.01 | 73.80 | |
| 24 h 0 °C | 44.22 ± 0.00 d | 0.76 ± 0.07 efg | 88.50 | 0.12 ± 0.05 c | 0.42 ± 0.05 de | 0.16 ± 0.04 de | 82.98 | 53.53 | 65.86 | |
| 24 h 2 °C | 45.73 ± 0.01 b | 1.23 ± 0.23 de | 79.67 | 0.06 ± 0.02 d | 0.45 ± 0.09 d | 0.17 ± 0.06 de | 90.52 | 50.94 | 62.73 | |
| ck | 47.58 ± 0.11 a | 5.75 ± 0.07 b | 0.68 ± 0.06 a | 0.91 ± 0.05 a | 0.45 ± 0.09 a |
| Pn (6 h) | Pn (3 d) | Pn (7 d) | Pn (11 d) | Pn (15 d) | |||
|---|---|---|---|---|---|---|---|
| R | Grains per spike | YN19 | 0.971 ** | 0.969 ** | 0.9773 ** | 0.9606 ** | 0.8835 ** |
| XM26 | 0.9875 ** | 0.9744 ** | 0.9667 ** | 0.9355 ** | 0.7255 * | ||
| 1000−grain weight | YN19 | 0.8858 ** | 0.9288 ** | 0.5585 | 0.695 * | 0.8825 ** | |
| XM26 | 0.9044 ** | 0.9203 ** | 0.9218 ** | 0.9359 ** | 0.8662 ** | ||
| Yield | YN19 | 0.9965 ** | 0.9453 ** | 0.9656 ** | 0.907 ** | 0.7703 ** | |
| XM26 | 0.9784 ** | 0.9625 ** | 0.9523 ** | 0.8547 ** | 0.6112 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Liu, L.; Chen, X.; Li, J. Effects of Low-Temperature Stress during the Anther Differentiation Period on Winter Wheat Photosynthetic Performance and Spike-Setting Characteristics. Plants 2022, 11, 389. https://doi.org/10.3390/plants11030389
Zhang Y, Liu L, Chen X, Li J. Effects of Low-Temperature Stress during the Anther Differentiation Period on Winter Wheat Photosynthetic Performance and Spike-Setting Characteristics. Plants. 2022; 11(3):389. https://doi.org/10.3390/plants11030389
Chicago/Turabian StyleZhang, Yan, Lvzhou Liu, Xiang Chen, and Jincai Li. 2022. "Effects of Low-Temperature Stress during the Anther Differentiation Period on Winter Wheat Photosynthetic Performance and Spike-Setting Characteristics" Plants 11, no. 3: 389. https://doi.org/10.3390/plants11030389
APA StyleZhang, Y., Liu, L., Chen, X., & Li, J. (2022). Effects of Low-Temperature Stress during the Anther Differentiation Period on Winter Wheat Photosynthetic Performance and Spike-Setting Characteristics. Plants, 11(3), 389. https://doi.org/10.3390/plants11030389