
New Insights on Phytochemical Features and Biological Properties of Alnus glutinosa Stem Bark

 ,
, 
 and
and
Abstract
1. Introduction
2. Results
2.1. Phytochemical Profile
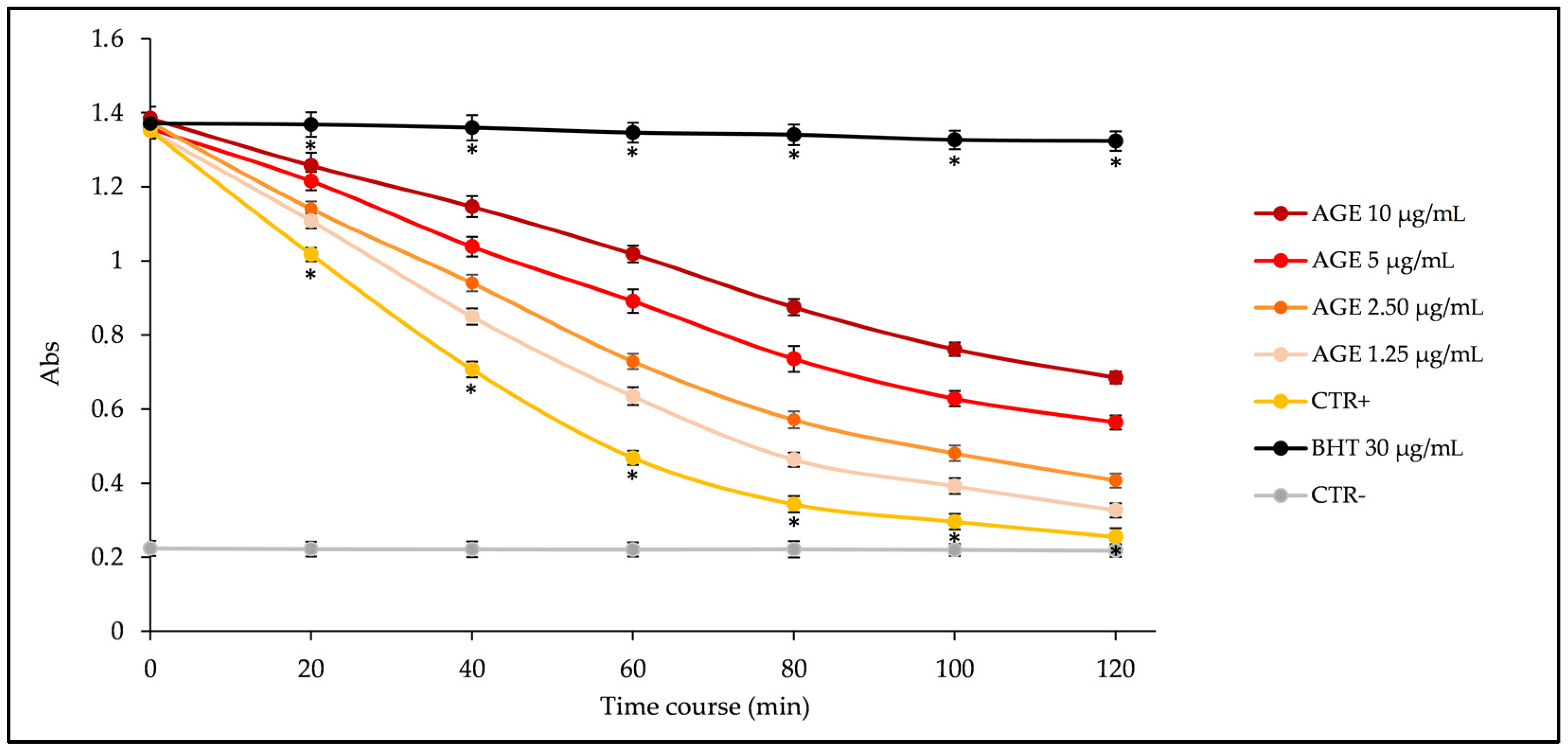
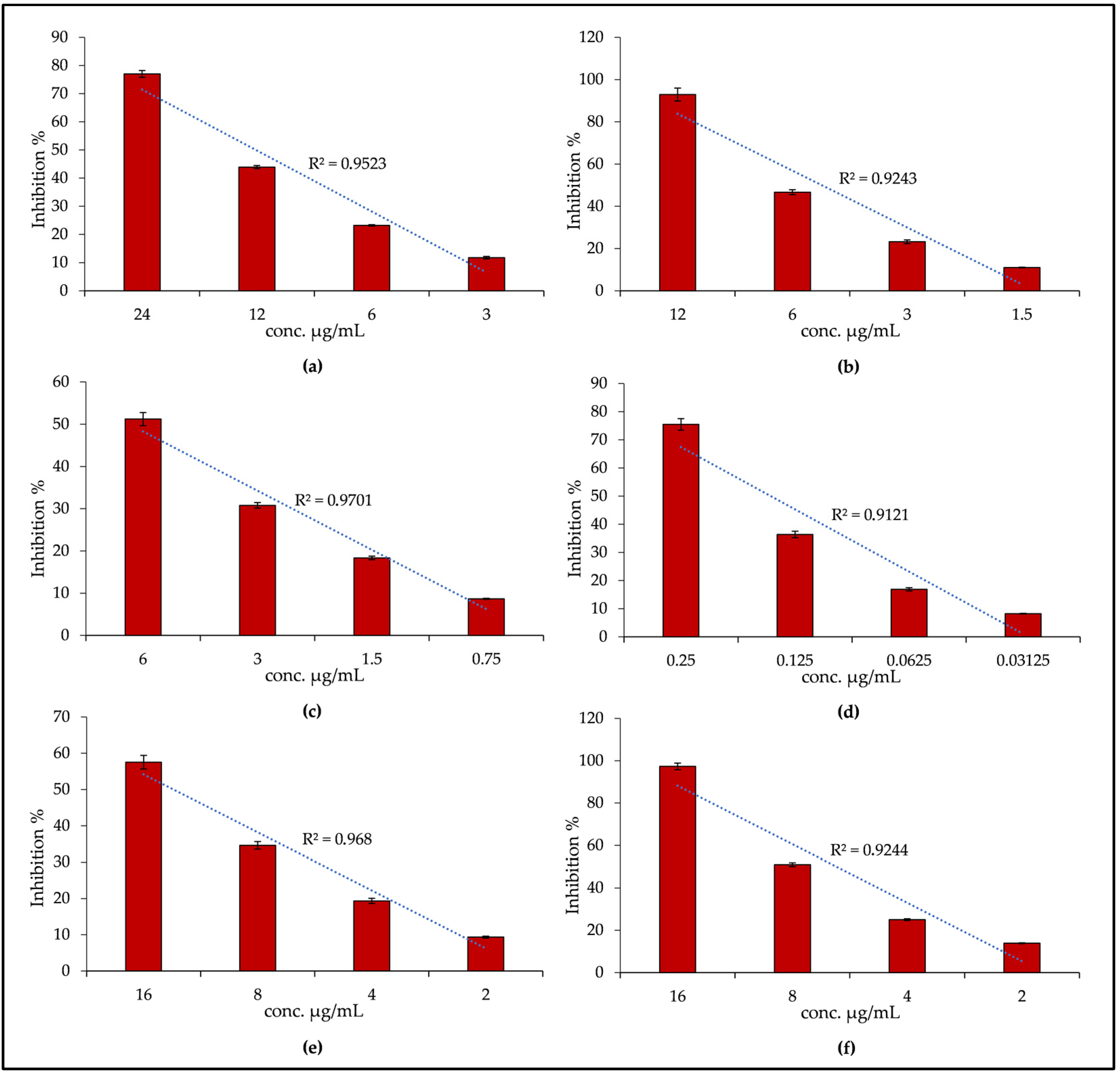
2.2. Biological Properties
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material and Sample Preparation
3.3. Total Phenols
3.4. Phytochemical Analysis
3.5. In Vitro Antioxidant and Anti-Inflammatory Activity
3.5.1. BCB Assay
3.5.2. DPPH Assay
3.5.3. FRAP Assay
3.5.4. TEAC Assay
3.5.5. ORAC Assay
3.5.6. Ferrozine Assay
3.5.7. BSA Denaturation Assay
3.5.8. Protease Inhibition Assay
3.6. In Vivo Anti-Angiogenic Activity
3.6.1. CAM Assay
3.6.2. EAP Activity on Zebrafish Embryos
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ren, X.; He, T.; Chang, Y.; Zhao, Y.; Chen, X.; Bai, S.; Wang, L.; Shen, M.; She, G. The Genus Alnus, a Comprehensive Outline of Its Chemical Constituents and Biological Activities. Molecules 2017, 22, 1383. [Google Scholar] [CrossRef] [PubMed]
- Lauberts, M.; Pals, M. Antioxidant Activity of Different Extracts from Black Alder (Alnus glutinosa) Bark with Greener Extraction Alternative. Plants 2021, 10, 2531. [Google Scholar] [CrossRef] [PubMed]
- Sati, S.; Sati, O.; Sati, N. Bioactive constituents and medicinal importance of genus Alnus. Pharmacogn. Rev. 2011, 5, 174. [Google Scholar] [CrossRef] [PubMed]
- Novaković, M.; Stanković, M.; Vučković, I.; Todorović, N.; Trifunović, S.; Tešević, V.; Vajs, V.; Milosavljević, S. Diarylheptanoids from Alnus glutinosa Bark and Their Chemoprotective Effect on Human Lymphocytes DNA. Planta Med. 2013, 79, 499–505. [Google Scholar] [CrossRef]
- Folkman, J. Angiogenesis in cancer, vascular, rheumatoid and other diseases. Nat. Med. 1995, 1, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Angiogenesis inhibitors: A new class of drugs. Cancer Biol. Ther. 2003, 2, S127–S133. [Google Scholar] [CrossRef]
- Agarwal, C.; Hofmann, T.; Visi-Rajczi, E.; Pásztory, Z. Low-frequency, green sonoextraction of antioxidants from tree barks of Hungarian woodlands for potential food applications. Chem. Eng. Process. Process Intensif. 2021, 159, 108221. [Google Scholar] [CrossRef]
- Skrypnik, L.; Grigorev, N.; Michailov, D.; Antipina, M.; Danilova, M.; Pungin, A. Comparative study on radical scavengingactivity and phenolic compounds content in water bark extracts of alder (Alnus glutinosa (L.) Gaertn.), oak (Quercus robur L.) and pine (Pinus sylvestris L.). Eur. J. Wood Wood Prod. 2019, 77, 879–890. [Google Scholar] [CrossRef]
- Di Vito, M.; Gentile, M.; Mattarelli, P.; Barbanti, L.; Micheli, L.; Mazzuca, C.; Garzoli, S.; Titubante, M.; Vitali, A.; Cacaci, M.; et al. Phytocomplex Influences Antimicrobial and Health Properties of Concentrated Glycerine Macerates. Antibiotics 2020, 9, 858. [Google Scholar] [CrossRef]
- Altınyay, Ç.; Eryılmaz, M.; Yazgan, A.N.; Sever Yılmaz, B.; Altun, M.L. Antimicrobial activity of some Alnus species. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4671–4674. [Google Scholar]
- Tung, N.H.; Kim, S.K.; Ra, G.C.; Zhao, Y.Z.; Sohn, D.H.; Kim, Y.H. Antioxidative and hepatoprotective diarylheptanoids from the bark of Alnus japonica. Planta Med. 2010, 76, 626–629. [Google Scholar] [CrossRef]
- Telysheva, G.; Dizhbite, T.; Bikovens, O.; Ponomarenko, J.; Janceva, S.; Krasilnikova, J. Structure and antioxidant activity of diarylheptanoids extracted from bark of grey alder (Alnus incana) and potential of biorefinery-based bark processing of European trees. Holzforschung 2011, 65, 623–629. [Google Scholar] [CrossRef]
- Ponomarenko, J.; Trouillas, P.; Martin, N.; Dizhbite, T.; Krasilnikova, J.; Telysheva, G. Elucidation of antioxidant properties of wood bark derived saturated diarylheptanoids: A comprehensive (DFT-supported) understanding. Phytochemistry 2014, 103, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.C.; Wang, M.H. Antioxidative activity and anti-inflammatory effects of diarylheptanoids isolated from Alnus hirsuta. J. Wood Sci. 2011, 57, 323–330. [Google Scholar] [CrossRef]
- Kuroyanagl, M.; Shimomae, M.; Nagashima, Y.; Muto, N.; Okuda, T.; Kawahara, N.; Nakane, T.; Sano, T. New diarylheptanoids from Alnus japonica and their antioxidative activity. Chem. Pham. Bull. 2005, 53, 1519–1523. [Google Scholar] [CrossRef]
- Kim, H.J.; Yeom, S.H.; Kim, M.K.; Paek, I.N.; Lee, M.W. Nitric oxide and prostaglandin E2 synthesis inhibitory activities of diarylheptanoids from the barks of Alnus japonica Steudel. Arch. Pharm. Res. 2005, 28, 177–179. [Google Scholar]
- Lee, M.A.; Lee, H.K.; Kim, S.H.; Kim, Y.C.; Sung, S.H. Chemical constituents of Alnus firma and their inhibitory activity on lipopolysaccharide-induced nitric oxide production in BV2 microglia. Planta Med. 2010, 76, 1007–1010. [Google Scholar] [CrossRef]
- Lee, C.J.; Lee, S.S.; Chen, S.C.; Ho, F.M.; Lin, W.W. Oregonin inhibits lipopolysaccharide-induced iNOS gene transcription and upregulates HO-1 expression in macrophages and microglia. Br. J. Pharmacol. 2005, 146, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Lundqvist, A.; Magnusson, L.U.; Ullstrom, C.; Krasilnikova, J.; Telysheva, T.; Dizhbite, T.; Hulten, L.M. Oregonin reduces lipid accumulation and proinflammatory responses in primary human macrophages. Biochem. Bioph. Res. Commun. 2015, 458, 693–699. [Google Scholar] [CrossRef]
- Lee, C.S.; Jang, E.R.; Kim, Y.J.; Lee, M.S.; Seo, S.J.; Lee, M.W. Hirsutenone inhibits lipopolysaccharide-activated NF-kB-induced inflammatory mediator production by suppressing Toll-like receptor 4 and ERK activation. Int. Immunopharmacol. 2010, 10, 520–525. [Google Scholar] [CrossRef]
- Lee, S.C.; Ko, H.H.; Seo, S.J.; Choi, Y.W.; Lee, M.W.; Myung, S.C.; Bang, H. Diarylheptanoid hirsutenone prevents tumor necrosis factor--stimulated production of inflammatory mediators in human keratinocytes through NF-kB inhibition. Int. Immunopharmacol. 2009, 9, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.Y.; Cai, X.F.; Na, M.K.; Lee, J.J.; Bae, K.W. Diarylheptanoids from Alnus hirsuta inhibit the NF-Kb activation and NO and TNF- production. Biol. Pharm. Bull. 2007, 30, 810–813. [Google Scholar] [CrossRef] [PubMed]
- Sajida, M.; Khana, M.R.; Shah, S.A.; Majid, M.; Ismail, H.; Maryam, S.; Batool, R.; Younis, T. Investigations on anti-inflammatory and analgesic activities of Alnus nitida Spach (Endl). stem bark in Sprague Dawley rats. J. Ethnopharmacol. 2017, 198, 407–416. [Google Scholar] [CrossRef]
- Velatooru, L.R.; Vakamullu, S.; Penugurti, V. Alpinoid c analog inhibits angiogenesis and induces apoptosis in COLO205 cell line. Chem. Interact. 2019, 308, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Smeriglio, A.; D’Angelo, V.; Denaro, M.; Trombetta, D.; Raimondo, F.M.; Germanò, M.P. Polyphenol Characterization, Antioxidant and Skin Whitening Properties of Alnus cordata Stem Bark. Chem. Biodivers. 2019, 16, e1900314. [Google Scholar] [CrossRef]
- Cornara, L.; Ambu, G.; Alberto, A.; Trombetta, D.; Smeriglio, A. Characterization of Ingredients Incorporated in the Traditional Mixed-Salad of the Capuchin Monks. Plants 2022, 11, 301. [Google Scholar] [CrossRef]
- Smeriglio, A.; Denaro, M.; Trombetta, D.; Ragusa, S.; Circosta, C. New Insights on Euphorbia dendroides L. (Euphorbiaceae): Polyphenol Profile and Biological Properties of Hydroalcoholic Extracts from Aerial Parts. Plants 2021, 10, 1621. [Google Scholar] [CrossRef]
- Muscarà, C.; Smeriglio, A.; Trombetta, D.; Mandalari, G.; La Camera, E.; Grassi, G.; Circosta, C. Phytochemical characterization and biological properties of two standardized extracts from a non-psychotropic Cannabis sativa L. cannabidiol (CBD)-chemotype. Phytother. Res. 2021, 35, 5269–5281. [Google Scholar] [CrossRef]
- Smeriglio, A.; Bonasera, S.; Germanò, M.P.; D’Angelo, V.; Barreca, D.; Denaro, M.; Monforte, M.T.; Galati, E.M.; Trombetta, D. Opuntia ficus-indica (L.) Mill. fruit as source of betalains with antioxidant, cytoprotective, and anti-angiogenic properties. Phytother. Res. 2019, 33, 1526–1537. [Google Scholar] [CrossRef]
- Denaro, M.; Smeriglio, A.; Trombetta, D. Antioxidant and Anti-Inflammatory Activity of Citrus Flavanones Mix and Its Stability after In Vitro Simulated Digestion. Antioxidants 2021, 10, 140. [Google Scholar] [CrossRef]
- Danna, C.; Bazzicalupo, M.; Ingegneri, M.; Smeriglio, A.; Trombetta, D.; Burlando, B.; Cornara, L. Anti-Inflammatory and Wound Healing Properties of Leaf and Rhizome Extracts from the Medicinal Plant Peucedanum ostruthium (L.) W. D. J. Koch. Molecules 2022, 27, 4271. [Google Scholar] [CrossRef] [PubMed]
- Bazzicalupo, M.; Cornara, L.; Burlando, B.; Cascini, A.; Denaro, M.; Smeriglio, A.; Trombetta, D. Carpobrotus edulis (L.) N.E.Br. extract as a skin preserving agent: From traditional medicine to scientific validation. J. Integr. Med. 2021, 19, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Smeriglio, A.; D’Angelo, V.; Denaro, M.; Trombetta, D.; Germanò, M.P. The Hull of Ripe Pistachio Nuts (Pistacia vera L.) as a Source of New Promising Melanogenesis Inhibitors. Plant Foods Hum. Nutr. 2021, 76, 111–117. [Google Scholar] [CrossRef]
- Smeriglio, A.; De Francesco, C.; Denaro, M.; Trombetta, D. Prickly Pear Betalain-Rich Extracts as New Promising Strategy for Intestinal Inflammation: Plant Complex vs. Main Isolated Bioactive Compounds. Front. Pharmacol. 2021, 12, 722398. [Google Scholar] [CrossRef] [PubMed]
- Smeriglio, A.; Denaro, M.; Di Gristina, E.; Mastracci, L.; Grillo, F.; Cornara, L.; Trombetta, D. Pharmacognostic approach to evaluate the micromorphological, phytochemical and biological features of Citrus lumia seeds. Food Chem. 2022, 375, 131855. [Google Scholar] [CrossRef] [PubMed]
- Smeriglio, A.; Denaro, M.; D’Angelo, V.; Germanò, M.P.; Trombetta, D. Antioxidant, Anti-Inflammatory and Anti-Angiogenic Properties of Citrus lumia Juice. Front. Pharmacol. 2020, 11, 593506. [Google Scholar] [CrossRef]
- Certo, G.; Costa, R.; D’Angelo, V.; Russo, M.; Albergamo, A.; Dugo, G.; Germanò, M.P. Anti-angiogenic activity and phytochemical screening of fruit fractions from Vitex agnus castus. Nat. Prod. Res. 2017, 31, 2850–2856. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # a | Compound | RT b | [M-H]− | MS/MS | λmax | AGE |
|---|---|---|---|---|---|---|
| (min) | (m/z) | (m/z) | (nm) | mg/g DE | ||
| 1 | Hirsutanonol 5-O-glucoside | 31.01 | 507 | 345 | 232; 282 | 13.95 ± 0.02 |
| 2 | Oregonin | 32.23 | 477 | 327 | 234; 282 | 484.18 ± 1.52 |
| 3 | Alnuside | 34.42 | 461 | 311 | 228; 254 | 5.02 ± 0.05 |
| 4 | Platyphylloside | 35.18 | 475 | 189 | 228; 248 | 3.36 ± 0.02 |
| 5 | Quercetin-3-sophoroside | 36.14 | 625 | 464 | 280; 330 | 1.66 ± 0.01 |
| 6 | Rubranoside A | 37.05 | 493 | 331 | 228; 282 | 3.76 ± 0.01 |
| 7 | 5-O-Methylhirsutanonol | 38.44 | 359 | 345 | 230; 282 | 17.91 ± 0.03 |
| 8 | Rubranoside B | 39.21 | 463 | 331 | 228; 282 | 27.21 ± 0.12 |
| 9 | Hirsutenone | 40.62 | 327 | 205 | 228; 282 | 33.71 ± 0.17 |
| 10 | Coumaroyl-oregonin | 42.63 | 623 | 477 | 222; 290 | 12.11 ± 0.05 |
| 11 | 5(S)-1,7-di(4-hydroxyphenyl)-5-O-β-D-[6-(E-3,4-DMC c-glucopyranosyl)] heptane-3-one | 45.44 | 665 | 475 | 223; 287 | 21.88 ± 0.08 |
| 12 | 5(S)-1-(4-hydroxyphenyl)-7-(3,4-dihydroxyphenyl)-5-O-β-D-[6-(3,4-DMC c-glucopyranosyl)] heptane-3-one | 47.53 | 681 | 475 | 223; 286 | 16.48 ± 0.14 |
| 13 | 3(R)-1,7-di(3,4-dihydroxyphenyl)-5-O-β-D-[6-(Z-3,4-DMC c-glucopyranosyl)] heptane | 49.42 | 683 | 493 | 224; 288 | 3.28 ± 0.03 |
| 14 | 3(R)-1,7-di(3,4-dihydroxyphenyl)-5-O-β-D-[6-(E-3,4,5-TMC d-glucopyranosyl)] heptane | 50.02 | 713 | 523 | 224; 289 | 3.61 ± 0.01 |
| Diarylheptanoids | 647.96 | |||||
| Oregonin | 484.18 | |||||
| Flavonoids | 1.66 | |||||
| Assay | AGE | Oregonin | RS b |
|---|---|---|---|
| IC50 (C.L. 95%) µg/mL | |||
| BCB | 8.95 (6.20–12.92) * | 9.50 (7.07–11.44) * | 0.24 (0.19–0.30) |
| DPPH | 12.21 (10.26–14.52) | 16.13 (13.85–18.67) * | 8.35 (2.07–13.66) |
| TEAC | 5.97 (4.69–7.60) | 6.46 (4.08–8.62) | 4.77 (3.85–5.90) |
| FRAP | 4.75 (1.89–7.95) | 7.01 (5.27–9.86) | 5.27 (4.09–6.79) |
| ORAC | 0.15 (0.12–0.17) * | 0.17 (0.12–0.20) * | 0.69 (0.31–1.53) |
| BSA a denaturation | 12.97 (10.40–16.18) | 9.35 (6.27–12.09) * | 17.82 (14.15–22.44) |
| Protease inhibitory activity | 5.47 (1.75–7.02) | 6.38 (4.18–8.77) | 6.33 (3.07–13.08) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smeriglio, A.; D’Angelo, V.; Cacciola, A.; Ingegneri, M.; Raimondo, F.M.; Trombetta, D.; Germanò, M.P. New Insights on Phytochemical Features and Biological Properties of Alnus glutinosa Stem Bark. Plants 2022, 11, 2499. https://doi.org/10.3390/plants11192499
Smeriglio A, D’Angelo V, Cacciola A, Ingegneri M, Raimondo FM, Trombetta D, Germanò MP. New Insights on Phytochemical Features and Biological Properties of Alnus glutinosa Stem Bark. Plants. 2022; 11(19):2499. https://doi.org/10.3390/plants11192499
Chicago/Turabian StyleSmeriglio, Antonella, Valeria D’Angelo, Anna Cacciola, Mariarosaria Ingegneri, Francesco Maria Raimondo, Domenico Trombetta, and Maria Paola Germanò. 2022. "New Insights on Phytochemical Features and Biological Properties of Alnus glutinosa Stem Bark" Plants 11, no. 19: 2499. https://doi.org/10.3390/plants11192499
APA StyleSmeriglio, A., D’Angelo, V., Cacciola, A., Ingegneri, M., Raimondo, F. M., Trombetta, D., & Germanò, M. P. (2022). New Insights on Phytochemical Features and Biological Properties of Alnus glutinosa Stem Bark. Plants, 11(19), 2499. https://doi.org/10.3390/plants11192499