
Impacts of Microcystins on Morphological and Physiological Parameters of Agricultural Plants: A Review



 ,
,  , and
, and 
Abstract
1. Introduction
2. Plant Responses Related to Toxin Concentration and Chemical Structure
3. Plant Response Related with the Growth Conditions
4. Plant Responses Related with the Stage of Development and Genotype
5. Plant Responses Related to Combined Effects of MCs and Other Toxic Compounds and Stressors
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef]
- Svirčev, Z.B.; Tokodi, N.; Drobac, D.; Codd, G.A. Cyanobacteria in aquatic ecosystems in Serbia: Effects on water quality, human health and biodiversity. Syst. Biodivers. 2014, 12, 261–270. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef]
- Gobler, C.J. Climate Change and Harmful Algal Blooms: Insights and perspective. Harmful Algae 2020, 91, 101731. [Google Scholar] [CrossRef]
- Cordeiro-Araújo, M.K.; Chia, M.A.; do Carmo Bittencourt-Oliveira, M. Potential human health risk assessment of cylindrospermopsin accumulation and depuration in lettuce and arugula. Harmful Algae 2017, 68, 217–223. [Google Scholar] [CrossRef]
- Saqrane, S.; Oudra, B. CyanoHAB occurrence and water irrigation cyanotoxin contamination: Ecological impacts and potential health risks. Toxins 2009, 1, 113–122. [Google Scholar] [CrossRef]
- Van Apeldoorn, M.E.; Van Egmond, H.P.; Speijers, G.J.A.; Bakker, G.J.I. Toxins of cyanobacteria. Mol. Nutr. Food Res. 2007, 51, 7–60. [Google Scholar] [CrossRef] [PubMed]
- Catherine, A.; Bernard, C.; Spoof, L.; Bruno, M. Microcystins and Nodularins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Lisa Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 107–126. [Google Scholar]
- Díez-Quijada, L.; Prieto, A.I.; Guzmán-Guillén, R.; Jos, A.; Cameán, A.M. Occurrence and toxicity of microcystin congeners other than MC-LR and MC-RR: A review. Food Chem. Toxicol. 2019, 125, 106–132. [Google Scholar] [CrossRef]
- Graham, J.L.; Loftin, K.A.; Meyer, M.T.; Ziegler, A.C. Cyanotoxin mixtures and taste-and-odor compounds in cyanobacterial blooms from the midwestern united states. Environ. Sci. Technol. 2010, 44, 7361–7368. [Google Scholar] [CrossRef]
- Corbel, S.; Mougin, C.; Bouaïcha, N. Cyanobacterial toxins: Modes of actions, fate in aquatic and soil ecosystems, phytotoxicity and bioaccumulation in agricultural crops. Chemosphere 2014, 96, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K.; Jones, G. Cyanobacterial Toxins. In Toxic Cyanobacteria in Water: A guide to their public health consequences, monitoring and management; Chorus, I., Bartram, J., Eds.; E & FN Spon: London, UK, 1999; pp. 55–124. [Google Scholar]
- Craig, M.; Luu, H.A.; McCready, T.L.; Williams, D.; Andersen, R.J.; Holmes, C.F.B. Molecular mechanisms underlying the interaction of motuporin and microycystins with type-1 and type-2A protein phosphatases. Biochem. Cell Biol. Biol. Cell. 1996, 74, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Li, T.; Zhang, Y.-L.; Guo, Z.-L.; Xu, L.-H. Effect of microcystin-LR on protein phosphatase 2A and its function in human amniotic epithelial cells. J. Zhejiang Univ. Sci. B 2011, 12, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Christen, V.; Meili, N.; Fent, K. Microcystin-LR induces endoplasmatic reticulum stress and leads to induction of NFκB, interferon-alpha, and tumor necrosis factor-alpha. Environ. Sci. Technol. 2013, 47, 3378–3385. [Google Scholar] [CrossRef]
- Valério, E.; Vasconcelos, V.; Campos, A. New Insights on the Mode of Action of Microcystins in Animal Cells—A Review. Mini-Rev. Med. Chem. 2016, 16, 1032–1041. [Google Scholar] [CrossRef]
- IARC. Ingested Nitrate and Nitrite, and Cyanobacterial Peptide Toxins; International Agency for Research on Cancer: Lyon, France, 2010; No. 94. [Google Scholar]
- Abe, T.; Lawson, T.; Weyers, J.D.B.; Codd, G.A. Microcystin-LR inhibits photosynthesis of Phaseolus vulgaris primary leaves: Implications for current spray irrigation practice. New Phytol. 1996, 133, 651–658. [Google Scholar] [CrossRef]
- Kurki-Helasmo, K.; Meriluoto, J. Microcystin uptake inhibits growth and protein phosphatase activity in mustard (Sinapis alba L.) seedlings. Toxicon 1998, 36, 1921–1926. [Google Scholar] [CrossRef]
- McElhiney, J.; Lawton, L.A.; Leifert, C. Investigations into the inhibitory effects of microcystins on plant growth, and the toxicity of plant tissues following exposure. Toxicon 2001, 39, 1411–1420. [Google Scholar] [CrossRef]
- Liang, C.; Liu, H. Response of hormone in rice seedlings to irrigation contaminated with cyanobacterial extract containing microcystins. Chemosphere 2020, 256, 127157. [Google Scholar] [CrossRef]
- Garda, T.; Kónya, Z.; Tándor, I.; Beyer, D.; Vasas, G.; Erdodi, F.; Vereb, G.; Papp, G.; Riba, M.; M-Hamvas, M.; et al. Microcystin-LR induces mitotic spindle assembly disorders in Vicia faba by protein phosphatase inhibition and not reactive oxygen species induction. J. Plant. Physiol. 2016, 199, 1–11. [Google Scholar] [CrossRef]
- Llana-Ruiz-Cabello, M.; Jos, A.; Cameán, A.; Oliveira, F.; Barreiro, A.; Machado, J.; Azevedo, J.; Pinto, E.; Almeida, A.; Campos, A.; et al. Analysis of the Use of Cylindrospermopsin and/or Microcystin-Contaminated Water in the Growth, Mineral Content, and Contamination of Spinacia oleracea and Lactuca sativa. Toxins 2019, 11, 624. [Google Scholar] [CrossRef]
- Freitas, M.; Azevedo, J.; Pinto, E.; Neves, J.; Campos, A.; Vasconcelos, V. Effects of microcystin-LR, cylindrospermopsin and a microcystin-LR/cylindrospermopsin mixture on growth, oxidative stress and mineral content in lettuce plants (Lactuca sativa L.). Ecotoxicol. Environ. Saf. 2015, 116, 59–67. [Google Scholar] [CrossRef]
- Machado, J.; Azevedo, J.; Freitas, M.; Pinto, E.; Almeida, A.; Vasconcelos, V.; Campos, A. Analysis of the use of microcystin-contaminated water in the growth and nutritional quality of the root-vegetable, Daucus carota. Environ. Sci. Pollut. Res. 2017, 24, 752–764. [Google Scholar] [CrossRef]
- Zhang, Y.; Whalen, J.K.; Sauvé, S. Phytotoxicity and bioconcentration of microcystins in agricultural plants: Meta-analysis and risk assessment. Environ. Pollut. 2020, 272, 115966. [Google Scholar] [CrossRef]
- Malaissi, L.; Vaccarini, C.A.; Hernández, M.P.; Ruscitti, M.; Arango, C.; Busquets, F.; Arambarri, A.M.; Giannuzzi, L.; Andrinolo, D.; Sedan, D. [D-Leu1]MC-LR and MC-LR: A Small–Large Difference: Significantly Different Effects on Phaseolus vulgaris L. (Fabaceae) Growth and Phototropic Response after Single Contact during Imbibition with Each of These Microcystin Variants. Toxins 2020, 12, 585. [Google Scholar] [CrossRef]
- Sedan, D.; Malaissi, L.; Vaccarini, C.A.; Ventosi, E.; Laguens, M.; Rosso, L.; Giannuzzi, L.; Andrinolo, D. [D-Leu1]MC-LR Has Lower PP1 Inhibitory Capability and Greater Toxic Potency than MC-LR in Animal and Plant Tissues. Toxins 2020, 12, 632. [Google Scholar] [CrossRef] [PubMed]
- Davies, P.J. The plant hormones: Their nature, occurrence, and functions. In Plant Hormones; Davies, P.J., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 1–15. [Google Scholar]
- Cao, Q.; Rediske, R.R.; Yao, L.; Xie, L. Effect of microcystins on root growth, oxidative response, and exudation of rice (Oryza sativa). Ecotoxicol. Environ. Saf. 2018, 149, 143–149. [Google Scholar] [CrossRef]
- Cao, Q.; Steinman, A.D.; Yao, L.; Xie, L. Increment of root membrane permeability caused by microcystins result in more elements uptake in rice (Oryza sativa). Ecotoxicol. Environ. Saf. 2017, 145, 431–435. [Google Scholar] [CrossRef]
- Liang, C.; Wang, W.; Wang, Y. Effect of irrigation with microcystins-contaminated water on growth, yield and grain quality of rice (Oryza sativa). Environ. Earth Sci. 2016, 75, 505. [Google Scholar] [CrossRef]
- Liang, C.; Ma, X.; Liu, H. Effect of microcystins at different rice growth stages on its yield, quality, and safety. Environ. Sci. Pollut. Res. 2020, 28, 1–13. [Google Scholar] [CrossRef]
- Liang, C.; Wang, W. Response and recovery of rice (Oryza sativa) seedlings to irrigation with microcystin-contaminated water. Environ. Earth Sci. 2015, 73, 4573–4580. [Google Scholar] [CrossRef]
- Al-Sultan, E.; Yousif, A.; Hatem, M.T. Toxic effects of purified microcystins from soil blue-green alga oscillatoria pseudogeminata on tomato plant lycopersicon esculentum. Baghdad Sci. J. 2019, 16, 169–177. [Google Scholar]
- do Carmo Bittencourt-Oliveira, M.; Cordeiro-Araújo, M.K.; Chia, M.A.; de Toledo Arruda-Neto, J.D.; de Oliveira, Ê.T.; dos Santos, F. Lettuce irrigated with contaminated water: Photosynthetic effects, antioxidative response and bioaccumulation of microcystin congeners. Ecotoxicol. Environ. Saf. 2016, 128, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Corbel, S.; Bouaïcha, N.; Nélieu, S.; Mougin, C. Soil irrigation with water and toxic cyanobacterial microcystins accelerates tomato development. Environ. Chem. Lett. 2015, 13, 447–452. [Google Scholar] [CrossRef]
- Pereira, A.L.; Azevedo, J.; Vasconcelos, V. Assessment of uptake and phytotoxicity of cyanobacterial extracts containing microcystins or cylindrospermopsin on parsley (Petroselinum crispum L.) and coriander (Coriandrum sativum L.). Environ. Sci. Pollut. Res. 2017, 24, 1999–2009. [Google Scholar] [CrossRef] [PubMed]
- Pflugmacher, S. Reduction in germination rate and elevation of peroxidase activity in Zea mays seedlings due to exposure to different microcystin analogues and toxic cell free cyanobacterial crude extract. J. Appl. Bot. Food Qual. 2007, 81, 45–48. [Google Scholar]
- Pflugmacher, S.; Hofmann, J.; Hübner, B. Effects on growth and physiological parameters in wheat (Triticum aestivum L.) grown in soil and irrigated with cyanobacterial toxin contaminated water. Environ. Toxicol. Chem. 2007, 26, 2710–2716. [Google Scholar] [CrossRef] [PubMed]
- Gehringer, M.M.; Kewada, V.; Coates, N.; Downing, T.G. The use of Lepidium sativum in a plant bioassay system for the detection of microcystin-LR. Toxicon 2003, 41, 871–876. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Jung, K.; Lundvall, L.; Neumann, S.; Peuthert, A. Effects of cyanobacterial toxins and cyanobacterial cell-free crude extract on germination of alfalfa (Medicago sativa) and induction of oxidative stress. Environ. Toxicol. Chem. 2006, 25, 2381–2387. [Google Scholar] [CrossRef] [PubMed]
- Gehringer, M.M. Microcystin-LR and okadaic acid-induced cellular effects: A dualistic response. FEBS Lett. 2004, 557, 1–8. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Aulhorn, M.; Grimm, B. Influence of a cyanobacterial crude extract containing microcystin-LR on the physiology and antioxidative defence systems of different spinach variants. New Phytol. 2007, 175, 482–489. [Google Scholar] [CrossRef]
- Corbel, S.; Mougin, C.; Martin-Laurent, F.; Crouzet, O.; Bru, D.; Nélieu, S.; Bouaïcha, N. Evaluation of phytotoxicity and ecotoxicity potentials of a cyanobacterial extract containing microcystins under realistic environmental concentrations and in a soil-plant system. Chemosphere 2015, 128, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Petrou, M.; Karas, P.A.; Vasileiadis, S.; Zafiriadis, I.; Papadimitriou, T.; Levizou, E.; Kormas, K.; Karpouzas, D.G. Irrigation of radish (Raphanus sativus L.) with microcystin-enriched water holds low risk for plants and their associated rhizopheric and epiphytic microbiome. Environ. Pollut. 2020, 266, 115208. [Google Scholar] [CrossRef]
- Levizou, E.; Papadimitriou, T.; Papavasileiou, E.; Papadimitriou, N.; Kormas, K.A. Root vegetables bioaccumulate microcystins-LR in a developmental stage-dependent manner under realistic exposure scenario: The case of carrot and radish. Agric. Water Manag. 2020, 240, 106274. [Google Scholar] [CrossRef]
- Lee, S.; Jiang, X.; Manubolu, M.; Riedl, K.; Ludsin, S.A.; Martin, J.F.; Lee, J. Fresh produce and their soils accumulate cyanotoxins from irrigation water: Implications for public health and food security. Food Res. Int. 2017, 102, 234–245. [Google Scholar] [CrossRef]
- Gu, Y.; Liang, C. Responses of antioxidative enzymes and gene expression in Oryza sativa L. and Cucumis sativus L. seedlings to microcystins stress. Ecotoxicol. Environ. Saf. 2020, 193, 110351. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Ren, X.; Liu, H.; Liang, C. Effect of irrigation with microcystins-contaminated water on growth and fruit quality of Cucumis sativus L. and the health risk. Agric. Water Manag. 2018, 204, 91–99. [Google Scholar] [CrossRef]
- El Khalloufi, F.; Oufdou, K.; Lahrouni, M.; Faghire, M.; Peix, A.; Ramírez-Bahena, M.H.; Vasconcelos, V.; Oudra, B. Physiological and antioxidant responses of Medicago sativa-rhizobia symbiosis to cyanobacterial toxins (Microcystins) exposure. Toxicon 2013, 76, 167–177. [Google Scholar] [CrossRef]
- Cao, Q.; Steinman, A.D.; Wan, X.; Xie, L. Combined toxicity of microcystin-LR and copper on lettuce (Lactuca sativa L.). Chemosphere 2018, 206, 474–482. [Google Scholar] [CrossRef]
- Levizou, E.; Statiris, G.; Papadimitriou, T.; Laspidou, C.S.; Kormas, K.A. Lettuce facing microcystins-rich irrigation water at different developmental stages: Effects on plant performance and microcystins bioaccumulation. Ecotoxicol. Environ. Saf. 2017, 143, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Steinman, A.D.; Wan, X.; Xie, L. Bioaccumulation of microcystin congeners in soil-plant system and human health risk assessment: A field study from Lake Taihu region of China. Environ. Pollut. 2018, 240, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Lahrouni, M.; Oufdou, K.; Faghire, M.; Peix, A.; El Khalloufi, F.; Vasconcelos, V.; Oudra, B. Cyanobacterial extracts containing microcystins affect the growth, nodulation process and nitrogen uptake of faba bean (Vicia faba L., Fabaceae). Ecotoxicology 2012, 21, 681–687. [Google Scholar] [CrossRef]
- Lahrouni, M.; Oufdou, K.; El Khalloufi, F.; Benidire, L.; Albert, S.; Göttfert, M.; Caviedes, M.A.; Rodriguez-Llorente, I.D.; Oudra, B.; Pajuelo, E. Microcystin-tolerant Rhizobium protects plants and improves nitrogen assimilation in Vicia faba irrigated with microcystin-containing waters. Environ. Sci. Pollut. Res. 2016, 23, 10037–10049. [Google Scholar] [CrossRef]
- Xiang, L.; Li, Y.-W.; Wang, Z.-R.; Liu, B.-L.; Zhao, H.-M.; Li, H.; Cai, Q.-Y.; Mo, C.-H.; Li, Q.X. Bioaccumulation and Phytotoxicity and Human Health Risk from Microcystin-LR under Various Treatments: A Pot Study. Toxins 2020, 12, 523. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Song, L.; Dai, J.; Gan, N.; Liu, Z. Effects of microcystins on the growth and the activity of superoxide dismutase and peroxidase of rape (Brassica napus L.) and rice (Oryza sativa L.). Toxicon 2004, 43, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Kuang, X.; Gu, J.D.; Tie, B.Q.; Yao, B.; Shao, J. Interactive effects of cadmium and Microcystis aeruginosa (cyanobacterium) on the growth, antioxidative responses and accumulation of cadmium and microcystins in rice seedlings. Ecotoxicology 2016, 25, 1588–1599. [Google Scholar] [CrossRef]
- Zhu, X.; Shen, Y.; Chen, X.; Hu, Y.O.O.; Xiang, H.; Tao, J.; Ling, Y. Biodegradation mechanism of microcystin-LR by a novel isolate of Rhizobium sp. TH and the evolutionary origin of the mlrA gene. Int. Biodeterior. Biodegrad. 2016, 115, 17–25. [Google Scholar] [CrossRef]
- Ramani, A.; Rein, K.; Shetty, K.G.; Jayachandran, K. Microbial degradation of microcystin in Florida’s freshwaters. Biodegradation 2012, 23, 35–45. [Google Scholar] [CrossRef]
- Chen, W.; Song, L.; Gan, N.; Li, L. Sorption, degradation and mobility of microcystins in Chinese agriculture soils: Risk assessment for groundwater protection. Environ. Pollut. 2006, 144, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Steinman, A.D.; Yao, L.; Xie, L. Effects of light, microorganisms, farming chemicals and water content on the degradation of microcystin-LR in agricultural soils. Ecotoxicol. Environ. Saf. 2018, 156, 141–147. [Google Scholar] [CrossRef]
- Cao, Q.; Steinman, A.D.; Su, X.; Xie, L. Effects of microcystins contamination on soil enzyme activities and microbial community in two typical lakeside soils. Environ. Pollut. 2017, 231, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Li, Y.-W.; Liu, B.-L.; Zhao, H.-M.; Li, H.; Cai, Q.-Y.; Mo, C.-H.; Wong, M.-H.; Li, Q.X. High ecological and human health risks from microcystins in vegetable fields in southern China. Environ. Int. 2019, 133, 105142. [Google Scholar] [CrossRef]
- Wu, X.; Xiao, B.; Li, R.; Wang, C.; Huang, J.; Wang, Z. Mechanisms and factors affecting sorption of microcystins onto natural sediments. Environ. Sci. Technol. 2011, 45, 2641–2647. [Google Scholar] [CrossRef]
- Miller, M.J.; Critchley, M.M.; Hutson, J.; Fallowfield, H.J. The adsorption of cyanobacterial hepatotoxins from water onto soil during batch experiments. Water Res. 2001, 35, 1461–1468. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Al Shehri, A.M. Microcystins in groundwater wells and their accumulation in vegetable plants irrigated with contaminated waters in Saudi Arabia. J. Hazard. Mater. 2009, 172, 310–315. [Google Scholar] [CrossRef]
- Cevallos-Casals, B.A.; Cisneros-Zevallos, L. Impact of germination on phenolic content and antioxidant activity of 13 edible seed species. Food Chem. 2010, 119, 1485–1490. [Google Scholar] [CrossRef]
- Tarasevičienė, Ž.; Viršilė, A.; Danilčenko, H.; Duchovskis, P.; Paulauskienė, A.; Gajewski, M. Effects of germination time on the antioxidant properties of edible seeds. CyTA J. Food 2019, 17, 447–454. [Google Scholar] [CrossRef]
- Pereira, S.; Saker, M.L.; Vale, M.; Vasconcelos, V.M. Comparison of sensitivity of grasses (Lolium perenne L. and Festuca rubra L.) and lettuce (Lactuca sativa L.) exposed to water contaminated with microcystins. Bull. Environ. Contam. Toxicol. 2009, 83, 81–84. [Google Scholar] [CrossRef] [PubMed]


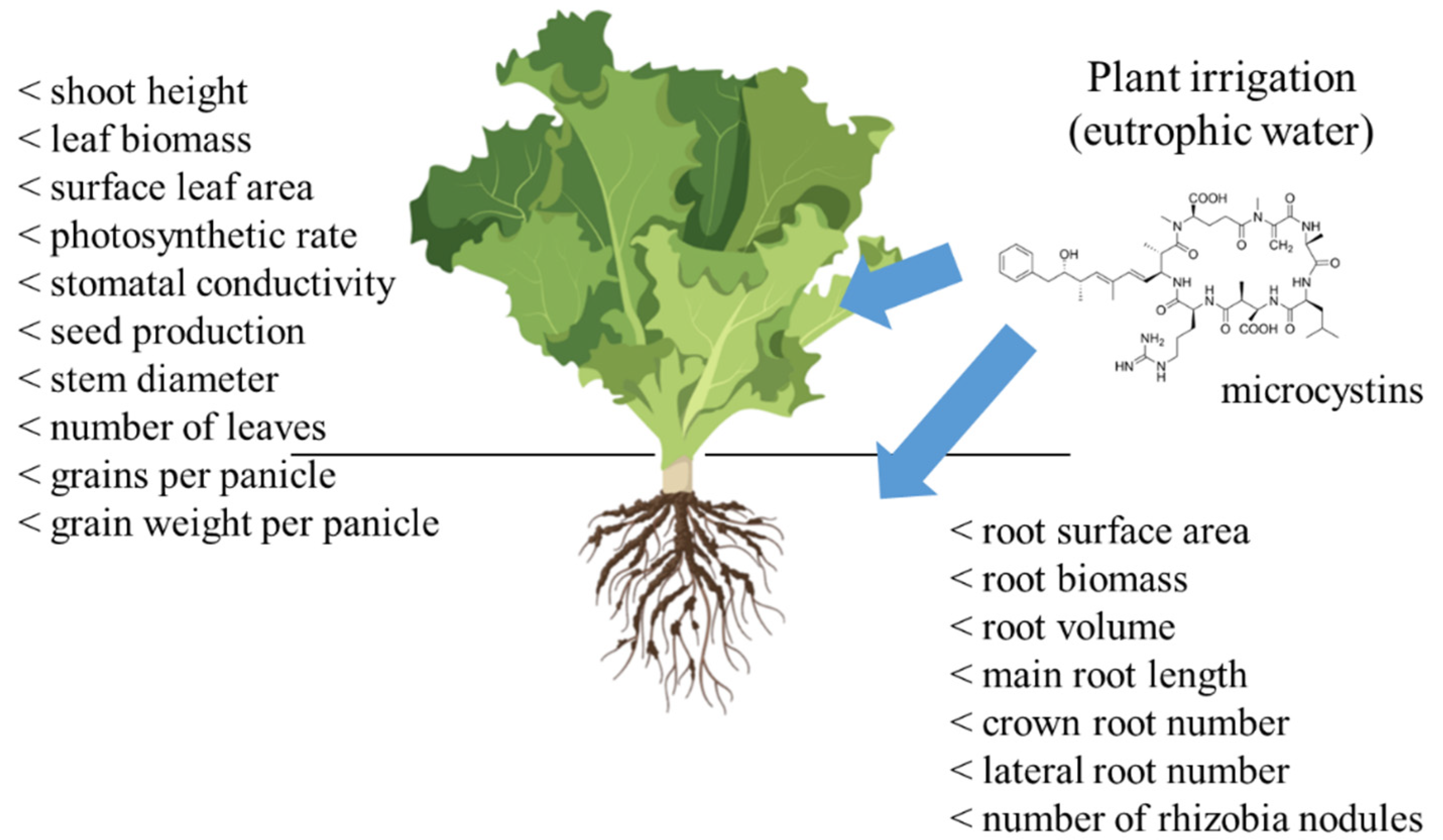{kind=link}
| Plant Species | Growth Stage (1) | Type Experiment/Exposure Time (days) (2) | Toxin (3) | Toxin Conc. (µg/L) (4) | Physiological and Morphological Parameters (5) | Reference |
|---|---|---|---|---|---|---|
| EUDICOTS | ||||||
| Spinacia oleracea (6 cultivars) | Seedlings | Soil culture/42 | MC-LR # | 0.5 | < photosynthetic oxygen production | [44] |
| Spinacia oleracea | Developed plants | Hydroponics/21 | MC-LR # | 50 | < leaf fresh weight | [23] |
| MC-LR, CYN # | 5 and 25 | |||||
| Lepidium sativum | Seeds | Germination/6 | MC-LR #* | 10 | < total fresh weight | [41] |
| 1–10 | < root length; leaf length | |||||
| Solanum lycopersicum | Seeds | Germination/7 | MCs | 50 and 100 | > radicle length | [45] |
| 500–20,000 | < radicle length | |||||
| Soil culture/14 | 5–100 | > shoot dry weight | ||||
| Seeds | Soil culture/90 | MCs | 5 | Anticipation of bloom of first flower; anticipation of first inflorescence emergence | [37] | |
| Developed plants | Soil culture/24 | MCs | 3–6 | < inhibition of root length; carbohydrate content | [35] | |
| 6 | < chlorophyll content; inhibition of stem length; decrease in surface leaf area | |||||
| Raphanus sativus | Seedlings | Soil culture/64 | MC-LR *# | raw water + MC-LR | < taproot fresh weight | [46] |
| 1-seeds | Soil culture 45 | MC-LR + MC-RR # | 3.76 | < leaf biomass; taproot biomass; taproot volume | [47] | |
| 2-seedlings | < taproot biomass; taproot volume | |||||
| Daucus carota | Seeds | Soil culture/75 | MC-LR + MC-RR # | 3.76 | < leaf biomass; taproot biomass; taproot volume | [47] |
| Developed plants | Soil culture/28 | MC-LR * | 1–100 | < taproot mass and taproot diameter; taproot diameter | [48] | |
| Developed plants | Soil experiment/32 | MC-LR # | 50 | < root fresh weight; < ascorbic acid (vitamin c) | [25] | |
| 10 | > max. fluorescence yield | |||||
| Cucumis sativus | Seedlings | Hydroponics/7 | MCs | 5 | < RGR; > H2O2, O2−, MDA | [49] |
| 1-seedlings | Hydroponics/7 | MCs | 1 | < plant height | [50] | |
| 10–1000 | < plant height; stem diameter; number of leaves; leaf area; root dry weight; shoot dry weight; yield | |||||
| 2-flowering | < plant height; stem diameter; leaf area | |||||
| 100–1000 | < number of leaves; shoot dry weight; root dry weight; yield | |||||
| 3-fruiting | 10–1000 | < plant height; leaf area; root dry weight | ||||
| 100–1000 | < stem diameter; number of leaves; shoot dry weight; yield | |||||
| Medicago sativa | Seeds | Germination/7 | MC-LR, MC-LW *# | 5 | < length of primary root; > lipid peroxidation | [42] |
| Seedlings | Irrigation/30 | MCs # | 5–20 | < shoot dry weight; root dry weight; nodules dry weight | [51] | |
| 10–20 | < root nodule number | |||||
| Lactuca sativa | 1-seeds | Germination/3 | MC-LR *, Cu | 50 | < germination | [52] |
| 2-seedlings | Hydroponics/14 | 5 | < total fresh weight | |||
| 50 | < root length | |||||
| 1000 | < shoot length | |||||
| 1-seeds | Soil experiment/60 | MCs | 5.11 | < root biomass; root/shoot biomass; > chlorophyll content | [53] | |
| 2-seedling -cotyledon stage | < root biomass; root/shoot biomass | |||||
| 3-seedling with 2 and 4 leaves | nc | |||||
| Seeds | Germination/7 | 50–5000 | > radicle length | [45] | ||
| Seedlings | Soil culture/60 | 2.61, 5.22 | nc | [54] | ||
| Developed plants | Soil culture/15 | MC-LR + MC-RR # | 0.65–13 | > net photosynthetic rate; leaf tissue transpiration; intercellular CO2 concentration | [36] | |
| 0.65 and 2.5 | > stomatal conductance | |||||
| Developed plants | Hydroponics/10 | MC-LR * | 1 and 10 | > leaf biomass | [24] | |
| 1 and 100 | > root biomass | |||||
| MC-LR + CYN * | 100 | < leaf biomass | ||||
| Developed plants | Soil culture/28 | MC-LR * | 5–100 | < leaf length | [48] | |
| 1–100 | < total leaf mass; number of leaves | |||||
| Vicia faba | 1-seeds | Germination/7 | MCs | 50 and 100 | < germination | [55] |
| 2-seedlings | Hydroponics/48 | < shoot dry weight; root dry weight; rhizobia nodules dry weight | ||||
| 2-seedlings | < shoot and root dry weight; rhizobia nodules dry weight | |||||
| 10–100 | < number of rhizobia nodules | |||||
| Seedlings | Soil culture/28 | MCs | 100 | < shoot and root dry weight; root nodule number | [56] | |
| Ipomoea batatas | Developed plants | Soil culture/10 | MC-LR #*, ACM | 150 | < main root length | [57] |
| MC-LR #*, IPW, CPS, ACM | < total weight; aerial part weight | |||||
| Brassica juncea | Developed plants | Soil culture/10 | MC-LR #*, IPW, ACM | 150 | < plant height; total weight | [57] |
| MC-LR #*, IPW, CPS, ACM | < main root length; aerial part weight | |||||
| Brassica alboglabra | Developed plants | Soil culture/10 | MC-LR #*, IPW | 150 | < main root length | [57] |
| MC-LR #*, IPW, CPS, ACM | < plant height; total weight; aerial part weight | |||||
| Petroselinum crispum | Developed plants | Soil culture/7 | MCs | 100–1000 | nc | [38] |
| Coriandrum sativum | Developed plants | Soil culture/7 | MCs | 100–1000 | nc | [38] |
| Brassica napus | Seeds | Germination/10d | MC-RR, MC-LR, MC-YR # | 600–3000 | < germination | [58] |
| 120–3000 | < plant height | |||||
| Phaseolus vulgaris | Seeds | Germination/1 | (D-Leu1) MC-LR * | 3500 | < germination; > stomatal density and conductivity | [27] |
| (D-Leu1) MC-LR and MC-LR * | delay seedling development; > morphological anomalies; delay phototropic response | |||||
| Seeds | Germination/1 | (D-Leu1) MC-LR and MC-LR * | 3500 | < chlorophyll content; delay seedling development | [28] | |
| 15000 | > lipid peroxidation | |||||
| (D-Leu1) MC-LRR * | 3500 | < germination | ||||
| Developed plants | Soil culture/28 | MC-LR * | 1 and 5 | > bean length | [48] | |
| 10-100 | < bean mass; number of beans | |||||
| MONOCOTS | ||||||
| Triticum aestivum | 1-seeds | Germination/3 | MC-RR, MC-LR # | 0.5 | < germination | [40] |
| 2-seedlings | Soil culture/15 | < shoot length; root length; photosynthetic oxygen production | ||||
| Triticum aestivum | Seeds | Germination/7 | MCs | 5000–20000 | < germination | [45] |
| 50 and 100 | > radicle length | |||||
| 20000 | < radicle length | |||||
| Zea mays | Seeds | Germination/-- | MCs (6 analogues) #* | 5 | < germination; shoot length; root length | [39] |
| Oryza sativa | Seeds | Germination/10 | MC-RR, MC-LR, MC-YR # | 600–3000 | < plant height | [58] |
| 120–3000 | < root length; root fresh weight | |||||
| Seedlings | Soil culture/60 | MCs | 2.61, 5.22 | nc | [54] | |
| Seedlings | Hydroponics/30 | MCs | 5–500 | < root dry weight | [30] | |
| 50–500 | < root length, surface area and volume; root surface area; root volume; lateral root number | |||||
| 500 | < crown root number | |||||
| Seedlings | Hydroponics/30 | MCs | 50–500 | < plant height; root length; shoot dry weight; >membrane permeability | [31] | |
| 5–500 | < root dry weight | |||||
| Seedlings | Hydroponics/7 | MCs | 1 | > root biomass | [32] | |
| 1–3000 | < stem biomass | |||||
| 100–3000 | < leaves biomass | |||||
| 1000–3000 | < root biomass; grains per panicle; grain weight per panicle; setting percentage | |||||
| Seedlings | Hydroponics/7 | MCs | 1 | > root surface area, shoot height | [21] | |
| 10 | < root surface area | |||||
| 100 | < shoot height | |||||
| Seedlings | Hydroponics/7 | MCs | 1 | > leaves dry weight; stem dry weight; roots dry weight | [34] | |
| 100–3000 | < leaves dry weight; stem dry weight; roots dry weight; net photosynthetic rate (Pn) | |||||
| Seedlings | Hydroponics/7 | MCs | 5 | > H2O2, O2-, MDA | [49] | |
| 10 | < RGR | |||||
| Seedlings | Hydroponics/21 | MCs | 10 | > shoot and root dry weight; P content in shoots and roots | [59] | |
| MCs, Cd | 10 | < root dry weight | ||||
| 1-seedlings | Hydroponics/7 | MCs | 10–1000 | < root surface area | [33] | |
| 100–1000 | < plant height; Net assimilation rate; filled grains per panicle; seed setting rate; panicle weight; soluble protein, sugar and starch in grain | |||||
| 2-booting | 100–1000 | < plant height; root surface area; Net assimilation rate; filled grains per panicle; seed setting rate; panicle weight; soluble protein, sugar and starch in grain | ||||
| 3-filling | 10–1000 | < seed setting rate | ||||
| 100–1000 | < root surface area; Net assimilation rate; filled grains per panicle; panicle weight; soluble protein, sugar and starch in grain | |||||
| 1000 | < plant height | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campos, A.; Redouane, E.M.; Freitas, M.; Amaral, S.; Azevedo, T.; Loss, L.; Máthé, C.; Mohamed, Z.A.; Oudra, B.; Vasconcelos, V. Impacts of Microcystins on Morphological and Physiological Parameters of Agricultural Plants: A Review. Plants 2021, 10, 639. https://doi.org/10.3390/plants10040639
Campos A, Redouane EM, Freitas M, Amaral S, Azevedo T, Loss L, Máthé C, Mohamed ZA, Oudra B, Vasconcelos V. Impacts of Microcystins on Morphological and Physiological Parameters of Agricultural Plants: A Review. Plants. 2021; 10(4):639. https://doi.org/10.3390/plants10040639
Chicago/Turabian StyleCampos, Alexandre, El Mahdi Redouane, Marisa Freitas, Samuel Amaral, Tomé Azevedo, Leticia Loss, Csaba Máthé, Zakaria A. Mohamed, Brahim Oudra, and Vitor Vasconcelos. 2021. "Impacts of Microcystins on Morphological and Physiological Parameters of Agricultural Plants: A Review" Plants 10, no. 4: 639. https://doi.org/10.3390/plants10040639
APA StyleCampos, A., Redouane, E. M., Freitas, M., Amaral, S., Azevedo, T., Loss, L., Máthé, C., Mohamed, Z. A., Oudra, B., & Vasconcelos, V. (2021). Impacts of Microcystins on Morphological and Physiological Parameters of Agricultural Plants: A Review. Plants, 10(4), 639. https://doi.org/10.3390/plants10040639