
Soil Amendment Using Biochar and Application of K-Humate Enhance the Growth, Productivity, and Nutritional Value of Onion (Allium cepa L.) under Deficit Irrigation Conditions

 , ,
, ,  ,
,  ,
,  , , ,
, , ,
Abstract
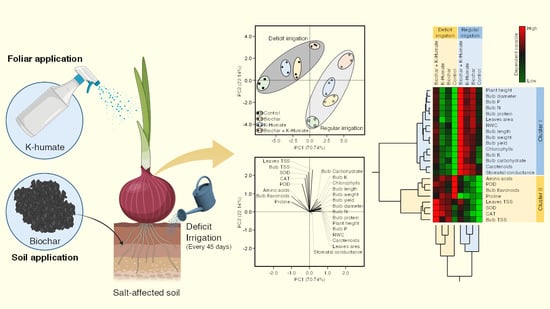
:
1. Introduction
2. Results
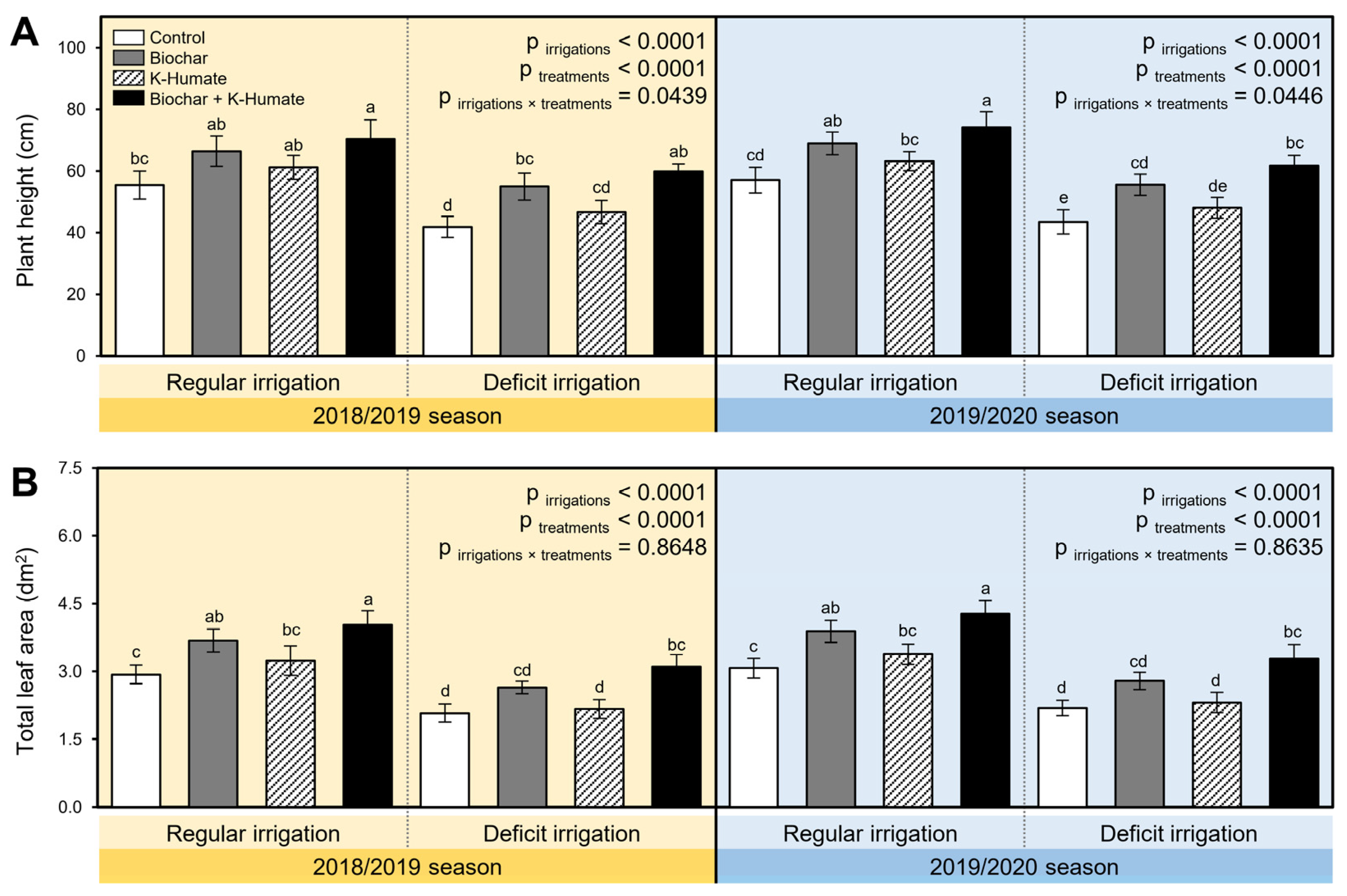
2.1. Soil Amendment Using Biochar and K-Humate Foliar Application Enhanced Onion Growth
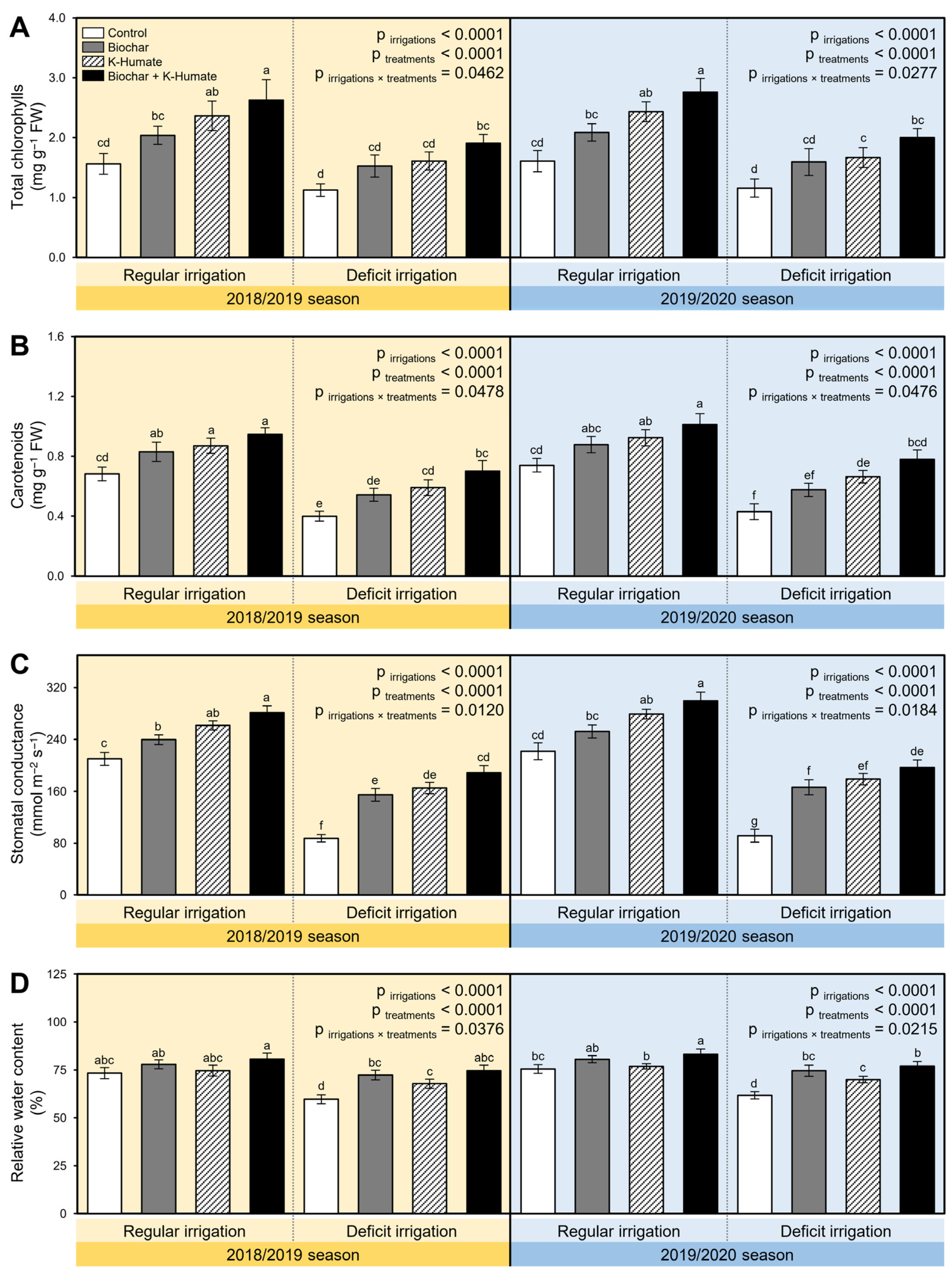
2.2. Biochar and K-Humate Application Boosted the Content of the Photosynthetic Pigments of Onion Plants
2.3. Exogenous Application of Biochar and K-Humate Enhanced Water Relations of Deficiently Irrigated Onion Plants
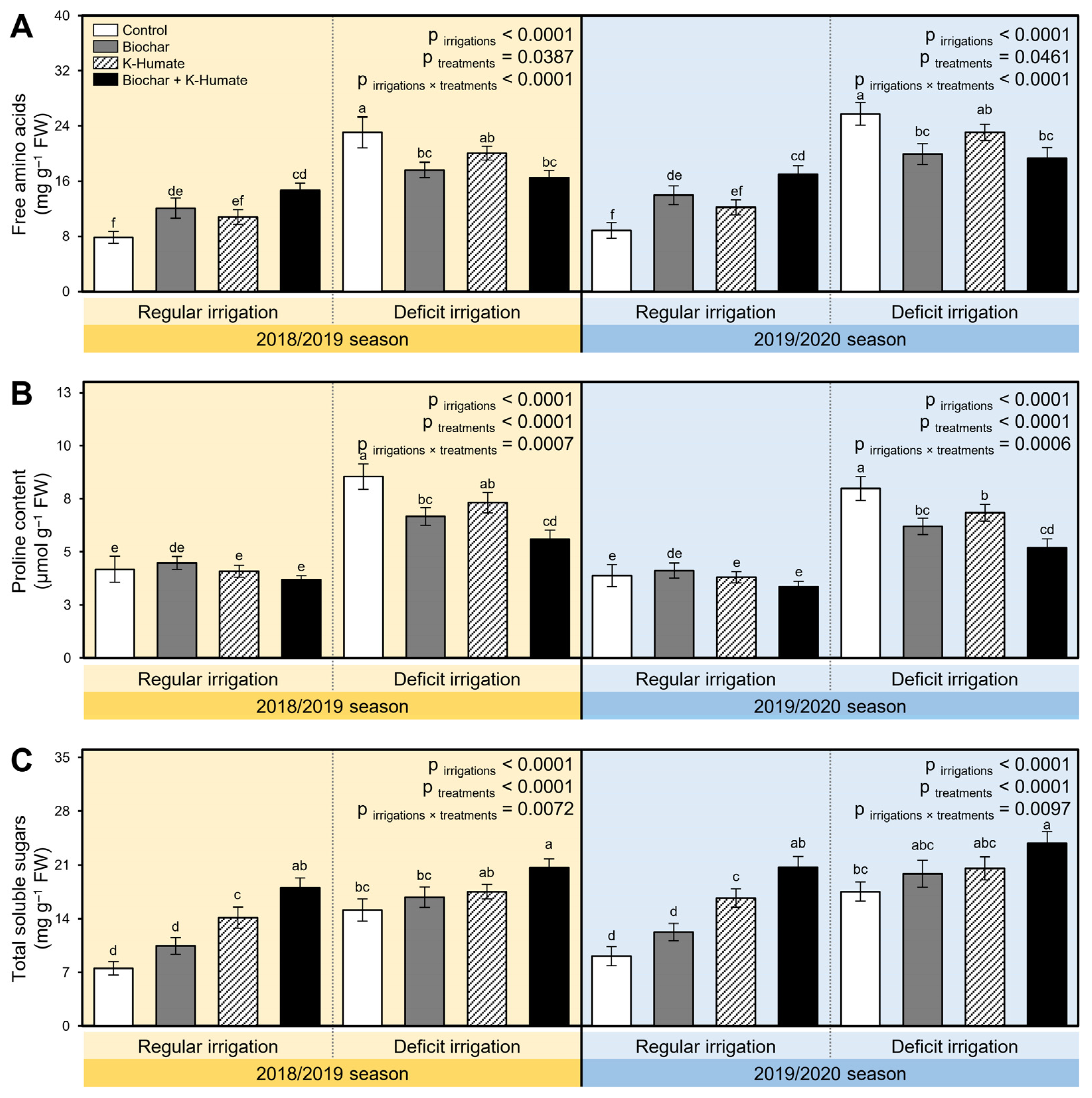
2.4. Soil Amendment Using Biochar and K-Humate Foliar Application Improved the Biochemical Response of Deficiently Irrigated Onion Plants
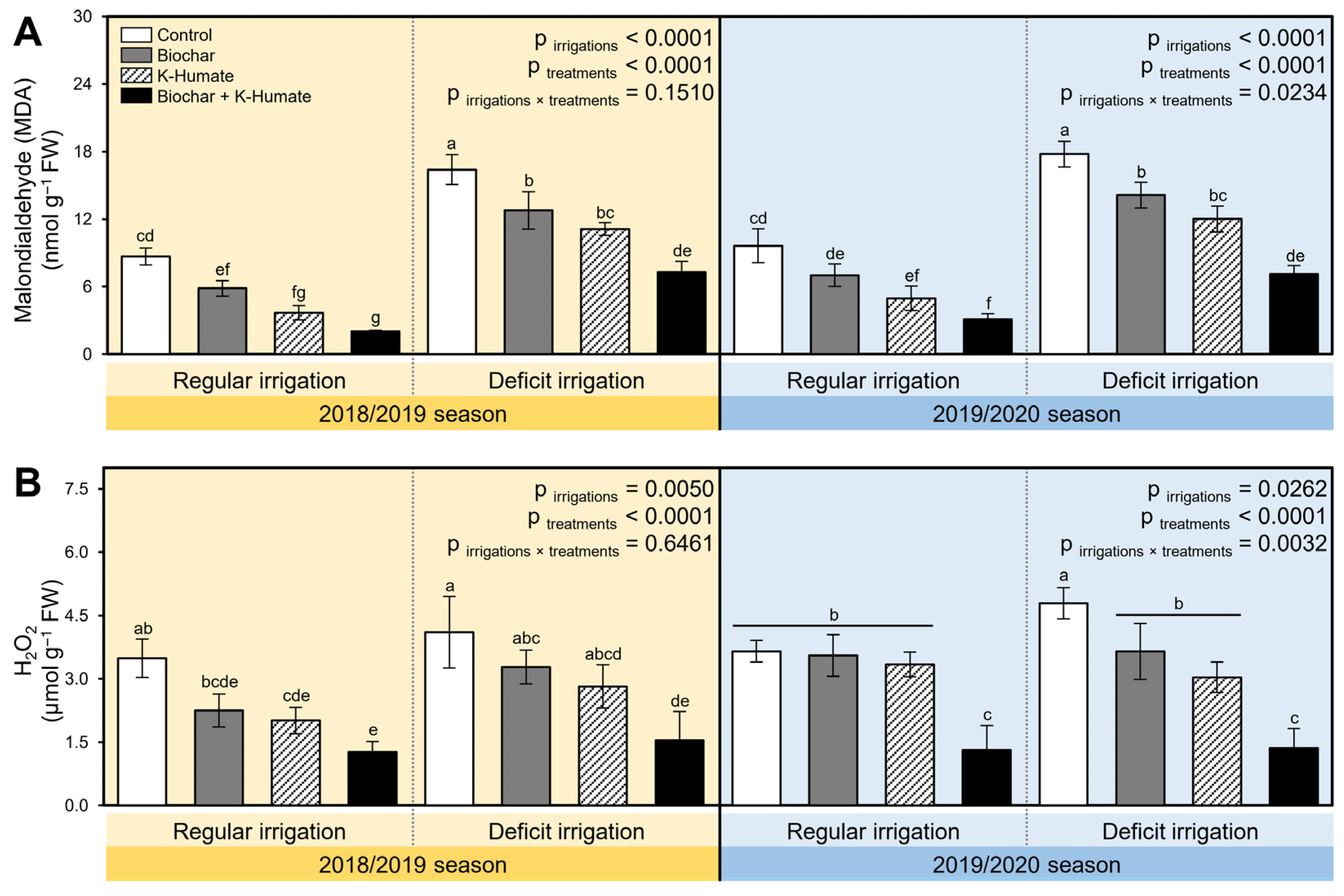
2.5. Integrated Biochar and K-Humate Application Diminish the Stress Biochemical Indicators in Stressed Onion Plants
2.6. Integrated Biochar and K-Humate Application Enhanced the Antioxidant-Related Enzymatic Activity in Deficiently Irrigated Onion Plants
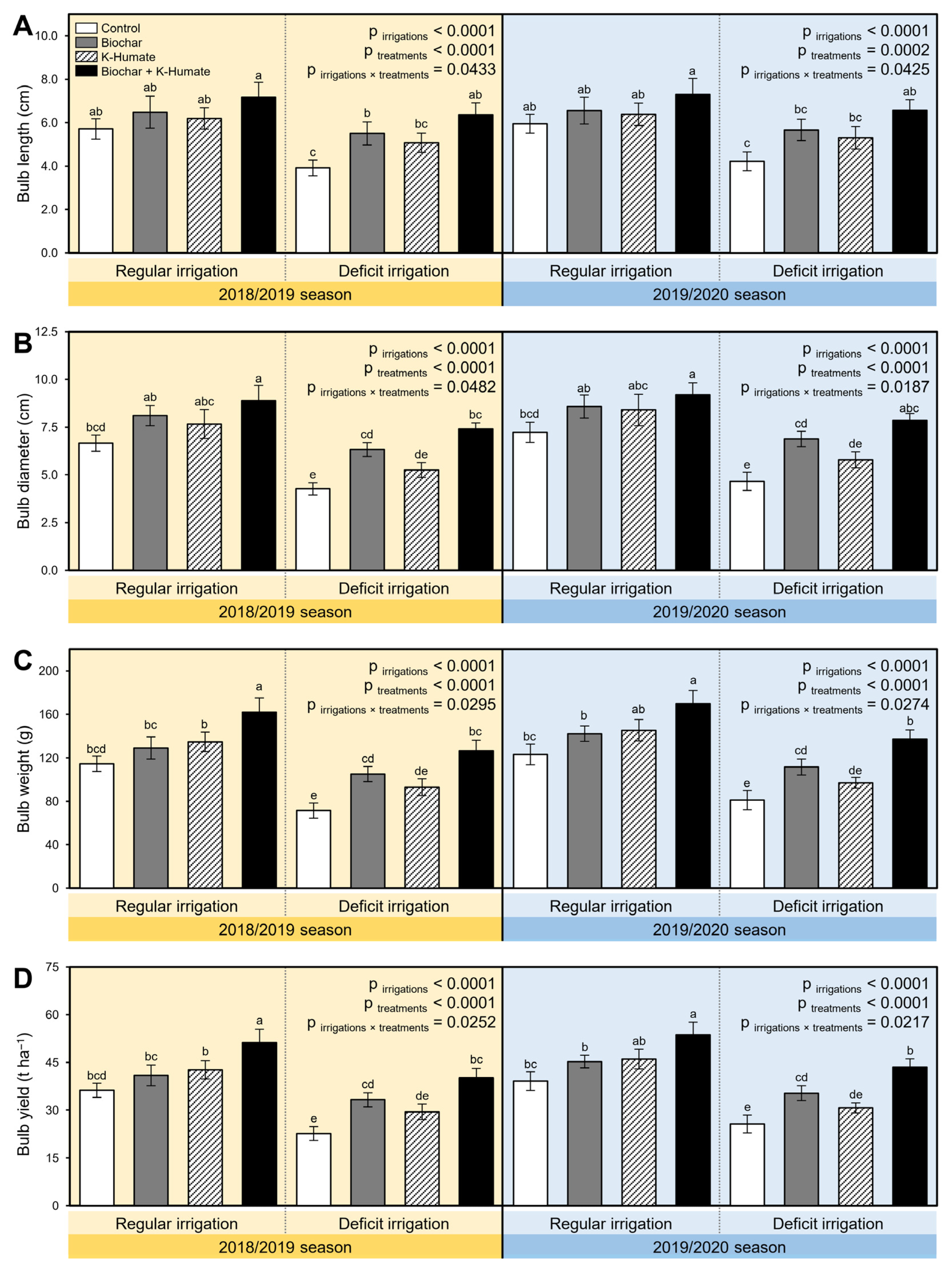
2.7. Soil Amendment Using Biochar and K-Humate Foliar Application Enhanced the Yield Traits of Deficient Irrigated Onion Plants
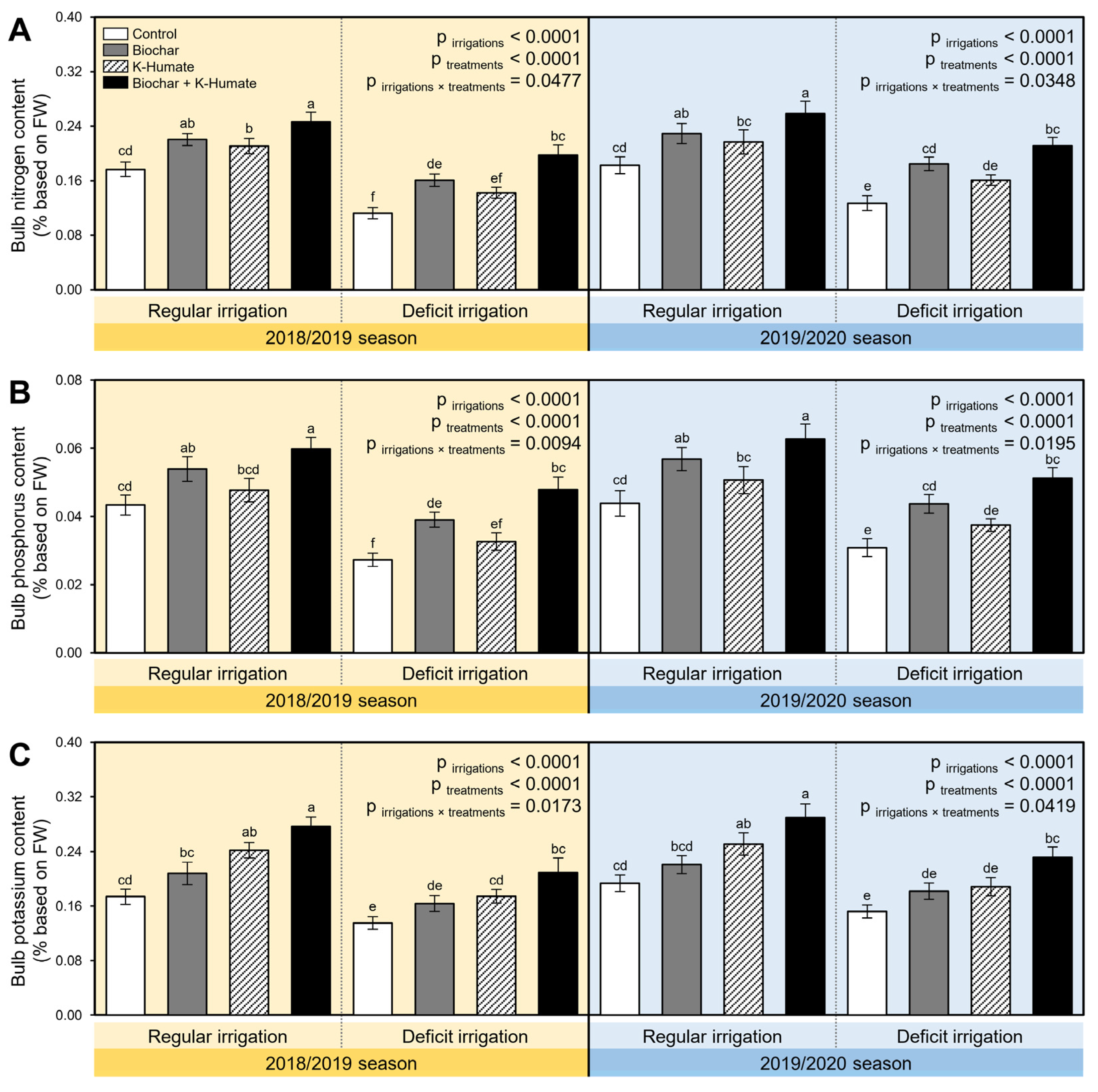
2.8. Biochar and K-Humate Supplementation Heightened the N, P, and K Content of Deficient Irrigated Onion Bulbs
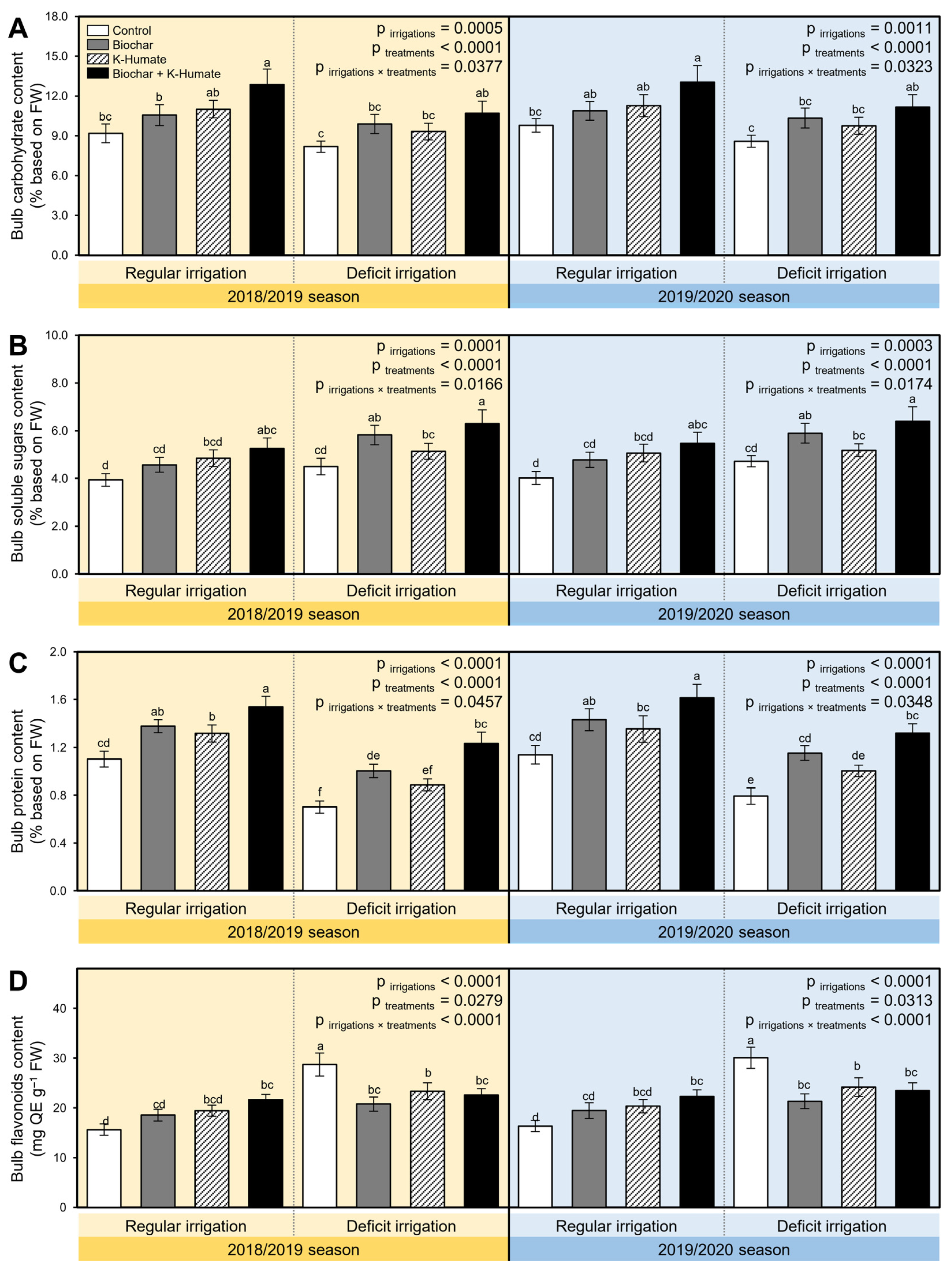
2.9. Combined Biochar and K-Humate Foliar Application Enriched the Nutritional Value of Deficient Irrigated Onion Bulbs
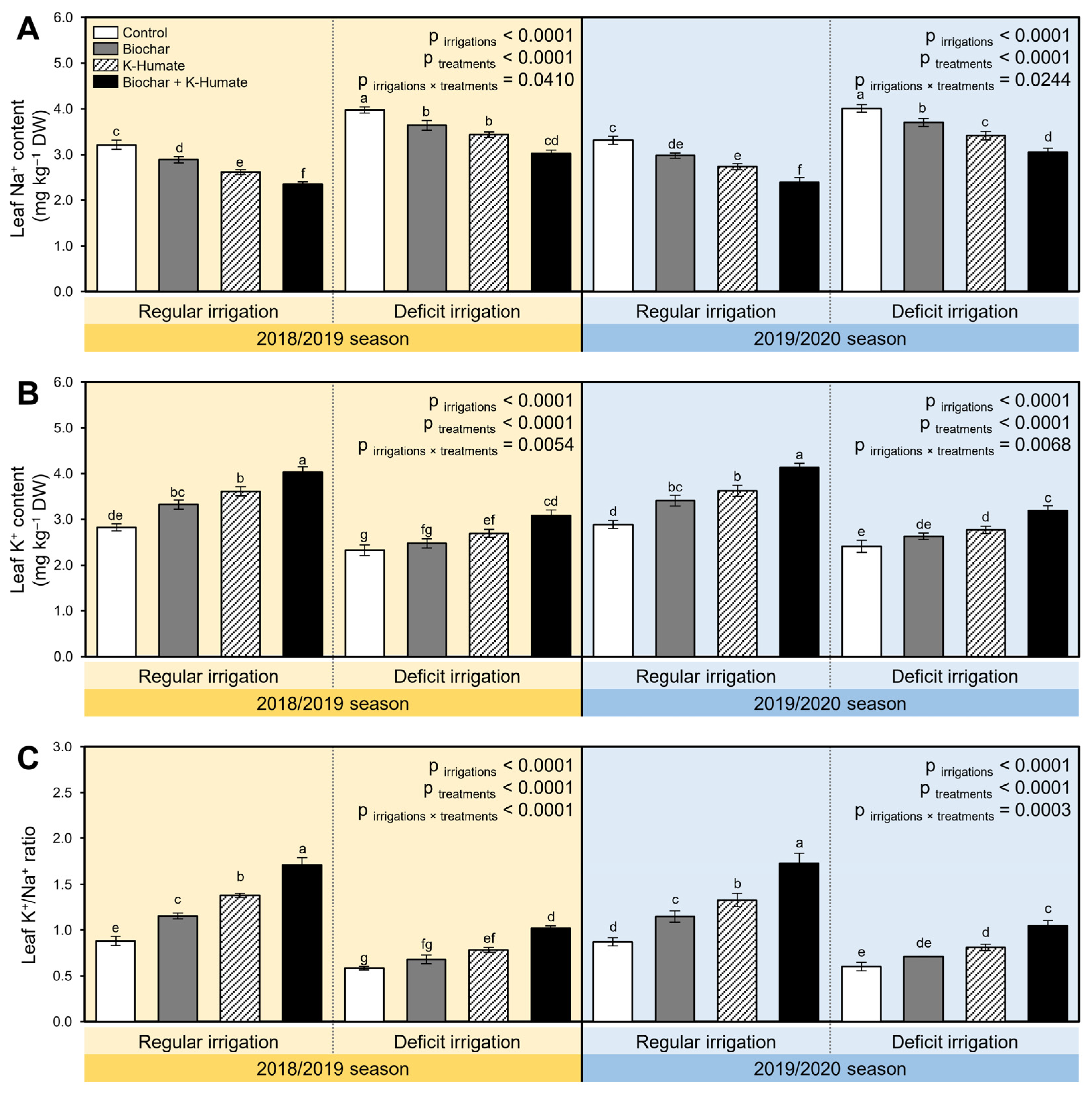
2.10. Combined Biochar and K-Humate Application Reduced the Na+ Accumulation in Onion Leaves under Deficient Irrigation Conditions
2.11. Principal Component Analysis (PCA) and Two-Way Hierarchical Cluster Analysis (HCA) Revealed the Differences among Treatments
3. Discussion
4. Materials and Methods
4.1. Experimental Site, Soil, and Meteorological Conditions
4.2. Plant Materials and Growth Conditions
4.3. Biochar Application
4.4. Potassium-Humate Application
4.5. Growth and Morphological Traits
4.5.1. Plant Height
4.5.2. Total Leaf Area
4.6. Photosynthetic Pigment Analysis
4.7. Water Relation Analysis
4.7.1. Stomatal Conductance (gs; mmol H2O m−2 s−1)
4.7.2. Relative Water Content
4.8. Biochemical Analysis of Onion Leaves
4.8.1. Free Amino Acids
4.8.2. Proline Content
4.8.3. Total Soluble Sugars (TSS)
4.9. Antioxidant Enzyme Activity and Stress Indicators in Onion Leaves
4.9.1. Antioxidant Enzyme Activity
4.9.2. Stress Indicators (Lipid Peroxidation (MDA) and H2O2)
4.10. Yield Traits
4.11. Nutritional Value Analyses of Onion Bulbs
4.11.1. N, P, and K Content
4.11.2. Total Carbohydrate, TSS, Protein, and Flavonoid Content
4.12. Na+ and K+ Accumulation in Onion Leaves
4.13. Experimental Design and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sidhu, J.S.; Ali, M.; Al-Rashdan, A.; Ahmed, N. Onion (Allium cepa L.) is potentially a good source of important antioxidants. J. Food Sci. Technol. 2019, 56, 1811–1819. [Google Scholar] [CrossRef]
- Metrani, R.; Singh, J.; Acharya, P.; Jayaprakasha, G.K.; Patil, B.S. Comparative Metabolomics Profiling of Polyphenols, Nutrients and Antioxidant Activities of Two Red Onion (Allium cepa L.) Cultivars. Plants 2020, 9, 1077. [Google Scholar] [CrossRef] [PubMed]
- Wakchaure, G.; Minhas, P.; Meena, K.K.; Singh, N.P.; Hegade, P.M.; Sorty, A.M. Growth, bulb yield, water productivity and quality of onion (Allium cepa L.) as affected by deficit irrigation regimes and exogenous application of plant bio–regulators. Agric. Water Manag. 2018, 199, 1–10. [Google Scholar] [CrossRef]
- Statista. Agriculture in Egypt—Statistics & Facts. Available online: https://www.statista.com/topics/5674/agriculture-in-egypt/ (accessed on 2 November 2021).
- Wakchaure, G.; Minhas, P.; Kumar, S.; Khapte, P.; Meena, K.; Rane, J.; Pathak, H. Quantification of water stress impacts on canopy traits, yield, quality and water productivity of onion (Allium cepa L.) cultivars in a shallow basaltic soil of water scarce zone. Agric. Water Manag. 2021, 249, 106824. [Google Scholar] [CrossRef]
- Bhatt, R.M.; Srinivasa, R.N.K.; Veere, G.R. Response of bulb onion (Allium cepa L.) to water stress: Photosynthesis, stomatal conductance and osmotic adjustment. Indian J. Hortic. 2006, 63, 276–280. [Google Scholar]
- El–Metwally, I.; Geries, L.; Saudy, H. Interactive effect of soil mulching and irrigation regime on yield, irrigation water use efficiency and weeds of trickle–irrigated onion. Arch. Agron. Soil Sci. 2021, 1–13. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Nahar, K.; Hossain, M.S.; Mahmud, J.A.; Hossen, M.S.; Masud, A.A.C.; Moumita; Fujita, M. Potassium: A Vital Regulator of Plant Responses and Tolerance to Abiotic Stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. Proc. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Chakraborty, K.; Bhaduri, D.; Meena, H.N.; Kalariya, K. External potassium (K+) application improves salinity tolerance by promoting Na+-exclusion, K+-accumulation and osmotic adjustment in contrasting peanut cultivars. Plant Physiol. Biochem. 2016, 103, 143–153. [Google Scholar] [CrossRef]
- Przygocka-Cyna, K.; Barłóg, P.; Grzebisz, W.; Spiżewski, T. Onion (Allium cepa L.) Yield and Growth Dynamics Response to In-Season Patterns of Nitrogen and Sulfur Uptake. Agronomy 2020, 10, 1146. [Google Scholar] [CrossRef]
- Kuroda, K.; Kurashita, H.; Arata, T.; Miyata, A.; Kawazoe, M.; Nobu, M.K.; Narihiro, T.; Ohike, T.; Hatamoto, M.; Maki, S.; et al. Influence of Green Tuff Fertilizer Application on Soil Microorganisms, Plant Growth, and Soil Chemical Parameters in Green Onion (Allium fistulosum L.) Cultivation. Agronomy 2020, 10, 929. [Google Scholar] [CrossRef]
- Hafez, E.M.; Alsohim, A.S.; Farig, M.; Omara, A.E.-D.; Rashwan, E.; Kamara, M.M. Synergistic Effect of Biochar and Plant Growth Promoting Rhizobacteria on Alleviation of Water Deficit in Rice Plants under Salt-Affected Soil. Agronomy 2019, 9, 847. [Google Scholar] [CrossRef] [Green Version]
- Yaghubi, K.; Ghaderi, N.; Vafaee, Y.; Javadi, T. Potassium silicate alleviates deleterious effects of salinity on two strawberry cultivars grown under soilless pot culture. Sci. Hortic. 2016, 213, 87–95. [Google Scholar] [CrossRef]
- Mehregan, B.; Mousavi Fard, S.; Rezaei Nezhad, A. Effect of foliar application of potassium silicate on some morphological, physiological and biochemical characteristics of Alternanthera repens L. under drought stress. J. Crops Improv. 2018, 20, 299–314. [Google Scholar] [CrossRef]
- Kheir, A.M.S.; Abouelsoud, H.M.; Hafez, E.M.; Ali, O.A.M. Integrated effect of nano-Zn, nano-Si, and drainage using crop straw–filled ditches on saline sodic soil properties and rice productivity. Arab. J. Geosci. 2019, 12, 1–8. [Google Scholar] [CrossRef]
- Ismail, E.E.M.; Halmy, M.M. Effect of Proline and Potassium Humate on Growth, Yield and Quality of Broad Bean under Saline Soil Conditions. J. Plant Prod. 2018, 9, 1141–1145. [Google Scholar] [CrossRef]
- Mahdi, A.; Badawy, S.; Latef, A.A.; El Hosary, A.; El Razek, U.A.; Taha, R. Integrated Effects of Potassium Humate and Planting Density on Growth, Physiological Traits and Yield of Vicia faba L. Grown in Newly Reclaimed Soil. Agronomy 2021, 11, 461. [Google Scholar] [CrossRef]
- Taha, R.S.; Seleiman, M.F.; Alotaibi, M.; Alhammad, B.A.; Rady, M.M.; Mahdi, A.H.A. Exogenous potassium treatments elevate salt tolerance and performances of Glycine max L. by boosting antioxidant defense system under actual saline field conditions. Agronomy 2020, 10, 1741. [Google Scholar] [CrossRef]
- Osman, M.E.; Mohsen, A.A.; Elfeky, S.S.; Mohamed, W.A.E. Response of Salt-Stressed Wheat (Triticum aestivum L.) to Potassium Humate Treatment and Potassium Silicate Foliar Application. Egypt. J. Bot. 2017, 57, 85–102. [Google Scholar] [CrossRef]
- Nehela, Y.; Mazrou, Y.S.A.; Alshaal, T.; Rady, A.M.S.; El-Sherif, A.M.A.; Omara, A.E.-D.; El-Monem, A.M.A.; Hafez, E.M. The Integrated Amendment of Sodic-Saline Soils Using Biochar and Plant Growth-Promoting Rhizobacteria Enhances Maize (Zea mays L.) Resilience to Water Salinity. Plants 2021, 10, 1960. [Google Scholar] [CrossRef]
- Singh, M.; Saini, R.K.; Singh, S.; Sharma, S.P. Potential of Integrating Biochar and Deficit Irrigation Strategies for Sustaining Vegetable Production in Water-limited Regions: A Review. HortScience 2019, 54, 1872–1878. [Google Scholar] [CrossRef] [Green Version]
- Abbasi, G.H.; Akhtar, J.; Anwar-Ul-Haq, M.; Ali, S.; Chen, Z.H.; Malik, W. Exogenous potassium differentially mitigates salt stress in tolerant and sensitive maize hybrids. Pak. J. Bot. 2014, 46, 135–146. [Google Scholar]
- Zaki, N.M.; Hassanein, M.S.; Ahmed, A.G.; El-Housini, E.A.; Tawfik, M.M. Foliar application of potassium to mitigate the adverse impact of salinity on some sugar beet varieties. 2: Effect on yield and quality. Middle East J. Agric. Res. 2014, 3, 448–460. [Google Scholar]
- Hafez, E.; Osman, H.; El-Razek, U.; Elbagory, M.; Omara, A.; Eid, M.; Gowayed, S. Foliar-Applied Potassium Silicate Coupled with Plant Growth-Promoting Rhizobacteria Improves Growth, Physiology, Nutrient Uptake and Productivity of Faba Bean (Vicia faba L.) Irrigated with Saline Water in Salt-Affected Soil. Plants 2021, 10, 894. [Google Scholar] [CrossRef] [PubMed]
- Hafez, E.M.; Omara, A.E.D.; Alhumaydhi, F.A.; El-Esawi, M.A. Minimizing hazard impacts of soil salinity and water stress on wheat plants by soil application of vermicompost and biochar. Physiol. Plant. 2020, 172, 587–602. [Google Scholar] [CrossRef]
- Hafez, E.M.; Kheir, A.M.S.; Badawy, S.A.; Rashwan, E.; Farig, M.; Osman, H.S. Differences in Physiological and Biochemical Attributes of Wheat in Response to Single and Combined Salicylic Acid and Biochar Subjected to Limited Water Irrigation in Saline Sodic Soil. Plants 2020, 9, 1346. [Google Scholar] [CrossRef]
- Mesbah, E.A.E. Effect of irrigation regimes and foliar spraying of potassium on yield, yield components and water use efficiency of wheat (Triticum aestivum L.) in Sandy Soils. World J. Agric. Sci. 2009, 5, 662–669. [Google Scholar]
- Ullah, A.; Ali, M.; Shahzad, K.; Ahmad, F.; Iqbal, S.; Rahman, M.H.U.; Ahmad, S.; Iqbal, M.M.; Danish, S.; Fahad, S.; et al. Impact of Seed Dressing and Soil Application of Potassium Humate on Cotton Plants Productivity and Fiber Quality. Plants 2020, 9, 1444. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, S. ROS and Oxidative Stress: Origin and Implication. In Reactive Oxygen Species in Plant Biology; Springer: New Delhi, India, 2019; pp. 1–31. [Google Scholar] [CrossRef]
- Nafees, M.; Fahad, S.; Shah, A.N.; Bukhari, M.A.; Maryam, I.; Ahmad, S.; Hussain, S. Reactive Oxygen Species Signaling in Plants. In Plant Abiotic Stress Tolerance; Springer: Cham, Switzerland, 2019; pp. 259–272. [Google Scholar] [CrossRef]
- Chung, W.-H. Unraveling new functions of superoxide dismutase using yeast model system: Beyond its conventional role in superoxide radical scavenging. J. Microbiol. 2017, 55, 409–416. [Google Scholar] [CrossRef]
- Ahmad, R.; Hussain, S.; Anjum, M.A.; Khalid, M.F.; Saqib, M.; Zakir, I.; Hassan, A.; Fahad, S.; Ahmad, S. Oxidative Stress and Antioxidant Defense Mechanisms in Plants under Salt Stress. In Plant Abiotic Stress Tolerance; Springer: Cham, Switzerland, 2019; pp. 191–205. [Google Scholar] [CrossRef]
- Alam, M.Z.; Carpenter-Boggs, L.; Hoque, A.; Ahammed, G.J. Effect of soil amendments on antioxidant activity and photosynthetic pigments in pea crops grown in arsenic contaminated soil. Heliyon 2020, 6, e05475. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Parvin, K.; Bardhan, K.; Nahar, K.; Anee, T.I.; Masud, A.A.C.; Fotopoulos, V. Biostimulants for the Regulation of Reactive Oxygen Species Metabolism in Plants under Abiotic Stress. Cells 2021, 10, 2537. [Google Scholar] [CrossRef]
- Wang, Y.; Pan, F.; Wang, G.; Zhang, G.; Wang, Y.; Chen, X.; Mao, Z. Effects of biochar on photosynthesis and antioxidative system of Malus hupehensis Rehd. seedlings under replant conditions. Sci. Hortic. 2014, 175, 9–15. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, Z.; Zhai, Y.; Lu, P.; Zhu, C. Effect of Straw Biochar on Soil Properties and Wheat Production under Saline Water Irrigation. Agronomy 2019, 9, 457. [Google Scholar] [CrossRef] [Green Version]
- Thomas, S.C.; Frye, S.; Gale, N.; Garmon, M.; Launchbury, R.; Machado, N.; Melamed, S.; Murray, J.; Petroff, A.; Winsborough, C. Biochar mitigates negative effects of salt additions on two herbaceous plant species. J. Environ. Manag. 2013, 129, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Kamara, M.M.; Rehan, M.; Ibrahim, K.M.; Alsohim, A.S.; Elsharkawy, M.M.; Kheir, A.M.S.; Hafez, E.M.; El-Esawi, M.A. Genetic Diversity and Combining Ability of White Maize Inbred Lines under Different Plant Densities. Plants 2020, 9, 1140. [Google Scholar] [CrossRef]
- Hemida, K.A.; Eloufey, A.Z.A.; El-Yazal, M.A.S.; Rady, M.M. Integrated effect of potassium humate and α-tocopherol applications on soil characteristics and performance of Phaseolus vulgaris plants grown on a saline soil. Arch. Agron. Soil Sci. 2017, 63, 1556–1571. [Google Scholar] [CrossRef]
- Klute, A.; Page, A.L. Methods of Soil Analysis. Part 2—Chemical and Microbiological Analysis, 2nd ed.; Klute, A., Page, A.L., Eds.; American Society of Agronomy: Madison, WI, USA, 1982; ISBN 0891180729. [Google Scholar]
- Campbell, D. Determination and Use of Soil Bulk Density in Relation to Soil Compaction. In Developments in Agricultural Engineering; Elsevier: Amsterdam, The Netherlands, 1994; pp. 113–139. [Google Scholar] [CrossRef]
- Trout, T.; Garcia-Castillas, I.; Hart, W. Soil-Water Engineering: Field and Laboratory Manual; Department of Agricultural and Chemical Engineering, Colordo State University: Fort Collins, CO, USA, 2012; ISBN 9788172337230. [Google Scholar]
- Hesse, P.R. A Textbook of Soil Chemical Analysis; Juan Murry Ltd.: London, UK, 1971. [Google Scholar]
- Gee, G.W.; Bauder, J.W. Particle-size analysis. In Methods of Soil Analysis. Part 1. Physical and Mineralogical Methods; Klute, A., Ed.; American Society of Agronomy, Inc.: Madison, WI, USA, 1986; pp. 383–411. [Google Scholar]
- Hafez, E.M.; Gowayed, S.M.; Nehela, Y.; Sakran, R.M.; Rady, A.M.S.; Awadalla, A.; Omara, A.E.-D.; Alowaiesh, B.F. Incorporated Biochar-Based Soil Amendment and Exogenous Glycine Betaine Foliar Application Ameliorate Rice (Oryza sativa L.) Tolerance and Resilience to Osmotic Stress. Plants 2021, 10, 1930. [Google Scholar] [CrossRef]
- Rafique, M.; Ortas, I.; Ahmed, I.A.; Rizwan, M.; Afridi, M.S.; Sultan, T.; Chaudhary, H.J. Potential impact of biochar types and microbial inoculants on growth of onion plant in differently textured and phosphorus limited soils. J. Environ. Manag. 2019, 247, 672–680. [Google Scholar] [CrossRef]
- Shafeek, M.R.; Helmy, Y.I.; Omar, N.M. Use of some bio-stimulants for improving the growth, yield and bulb quality of onion plants (Allium cepa L.) under sandy soil conditions. Middle East J. Appl. Sci. 2015, 5, 68–75. [Google Scholar]
- Mohsen, A.A.M.; Ibraheim, S.K.A.; Abdel-Fattah, M.K. Effect of potassium humate, nitrogen bio fertilizer and molybdenum on growth and productivity of garlic (Allium sativum L.). Curr. Sci. Int. 2017, 6, 75–85. [Google Scholar]
- Wallace, D.H.; Munger, H.M. Studies of the Physiological Basis for Yield Differences. I. Growth Analysis of Six Dry Bean Varieties 1. Crops Sci. 1965, 5, 343–348. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 48, 350–382. [Google Scholar]
- Mullan, D.; Pietragalla, J. Leaf relative water content. In Physiological Breeding II: A Field Guide to Wheat Phenotyping; Cimmyt: México-Veracruz, Mexico, 2012; pp. 25–27. [Google Scholar]
- Hansen, S.L. Content of Free Amino Acids in Onion (Allium cepa L.) as Influenced by the Stage of Development at Harvest and Long-term Storage. Acta Agric. Scand. Sect. B—Plant Soil Sci. 2001, 51, 77–83. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Sadasivam, S.; Manickam, A. Biochemical Methods, 3rd ed.; Sadasivam, S., Manickam, A., Eds.; New Age International Publishers: New Delhi, India, 2008. [Google Scholar]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Aebi, H.B. Isolation, purification, characterization, and assay of antioxygenic enzymes: Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Vetter, J.L.; Steinberg, M.P.; Nelson, A.I. Enzyme Assay, Quantitative Determination of Peroxidase in Sweet Corn. J. Agric. Food Chem. 1958, 6, 39–41. [Google Scholar] [CrossRef]
- Du, Z.; Bramlage, W.J. Modified thiobarbituric acid assay for measuring lipid oxidation in sugar-rich plant tissue extracts. J. Agric. Food Chem. 1992, 40, 1566–1570. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. AOAC Official Methods of Analysis; Association of Official Analytical Chemists: Washington, DC, USA, 1975; p. 1018. [Google Scholar]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H. Methods of Soil Analysis. Part 3, Chemical Methods; John Wiley & Sons: Hoboken, NJ, USA, 2020; ISBN 978-0-891-18825-4. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis; Prentice Hall Inc.: Englewood Cliffs, NJ, USA, 1958. [Google Scholar]
- Dubois, M.; Gilles, K.; Hamilton, J.; DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. AOAC Official Methods, 17th ed.; 2nd Version; Analysis of Association of Official Agriculture Chemists: Washington, DC, USA, 2000; p. 2200. [Google Scholar]
- Zou, Y.; Lu, Y.; Wei, D. Antioxidant Activity of a Flavonoid-Rich Extract of Hypericum perforatum L. in Vitro. J. Agric. Food Chem. 2004, 52, 5032–5039. [Google Scholar] [CrossRef] [PubMed]
- SAS JMP®. Data Analysis Software; Version 15; SAS Institute Inc.: Cary, NC, USA, 2013; Available online: http://www.jmp.com/ (accessed on 15 February 2013).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Attribute | Season | |
|---|---|---|
| 2018/2019 | 2019/2020 | |
| Soil texture | Clay loam | Clay loam |
| Organic matter (%) | 1.51 | 1.48 |
| Electric conductivity (EC; dS m−1) | 11.14 | 12.09 |
| Field capacity (%) | 29.4 | 31.1 |
| pH | 8.35 | 8.58 |
| Soil bulk density (g cm−3) | 1.42 | 1.38 |
| Total porosity (%) | 45.25 | 47.65 |
| Cations (meq L−1) | ||
| Na+ | 17.78 | 18.65 |
| K+ | 10.73 | 11.86 |
| Mg+2 | 14.76 | 16.23 |
| Ca+2 | 16.54 | 18.29 |
| Anions (meq L−1) | ||
| Cl− | 23.56 | 24.21 |
| HCO3− | 17.22 | 19.67 |
| SO4−2 | 19.03 | 21.15 |
| Year Month | 2018/2019 | 2019/2020 | ||||||
|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | Rainfall (mm) | Relative Humidity (%) | Temperature (°C) | Rainfall (mm) | Relative Humidity (%) | |||
| Max | Min | Max | Min | |||||
| Oct. | 26.3 | 17.2 | 0.98 | 32.6 | 25.3 | 16.2 | 0.94 | 31.6 |
| Dec. | 25.9 | 15.3 | 0.85 | 34.2 | 24.9 | 14.3 | 0.82 | 33.2 |
| Jan. | 24.5 | 13.2 | 1.1 | 35.1 | 23.2 | 12.4 | 0.54 | 32.7 |
| Feb. | 22.3 | 10.3 | 3.1 | 46.2 | 20.3 | 11.1 | 3.32 | 42.4 |
| Mar. | 21.4 | 9.7 | 6.4 | 44.3 | 20.6 | 10.7 | 6.85 | 43.1 |
| April | 23.7 | 13.8 | 0.5 | 43.8 | 22.5 | 12.5 | 0.63 | 44.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelrasheed, K.G.; Mazrou, Y.; Omara, A.E.-D.; Osman, H.S.; Nehela, Y.; Hafez, E.M.; Rady, A.M.S.; El-Moneim, D.A.; Alowaiesh, B.F.; Gowayed, S.M. Soil Amendment Using Biochar and Application of K-Humate Enhance the Growth, Productivity, and Nutritional Value of Onion (Allium cepa L.) under Deficit Irrigation Conditions. Plants 2021, 10, 2598. https://doi.org/10.3390/plants10122598
Abdelrasheed KG, Mazrou Y, Omara AE-D, Osman HS, Nehela Y, Hafez EM, Rady AMS, El-Moneim DA, Alowaiesh BF, Gowayed SM. Soil Amendment Using Biochar and Application of K-Humate Enhance the Growth, Productivity, and Nutritional Value of Onion (Allium cepa L.) under Deficit Irrigation Conditions. Plants. 2021; 10(12):2598. https://doi.org/10.3390/plants10122598
Chicago/Turabian StyleAbdelrasheed, Khaled G., Yasser Mazrou, Alaa El-Dein Omara, Hany S. Osman, Yasser Nehela, Emad M. Hafez, Asmaa M. S. Rady, Diaa Abd El-Moneim, Bassam F. Alowaiesh, and Salah M. Gowayed. 2021. "Soil Amendment Using Biochar and Application of K-Humate Enhance the Growth, Productivity, and Nutritional Value of Onion (Allium cepa L.) under Deficit Irrigation Conditions" Plants 10, no. 12: 2598. https://doi.org/10.3390/plants10122598
APA StyleAbdelrasheed, K. G., Mazrou, Y., Omara, A. E.-D., Osman, H. S., Nehela, Y., Hafez, E. M., Rady, A. M. S., El-Moneim, D. A., Alowaiesh, B. F., & Gowayed, S. M. (2021). Soil Amendment Using Biochar and Application of K-Humate Enhance the Growth, Productivity, and Nutritional Value of Onion (Allium cepa L.) under Deficit Irrigation Conditions. Plants, 10(12), 2598. https://doi.org/10.3390/plants10122598