A Multispecies Assessment to Identify the Functional Connectivity of Amphibians in a Human-Dominated Landscape


Abstract
1. Introduction
2. Material and Methods
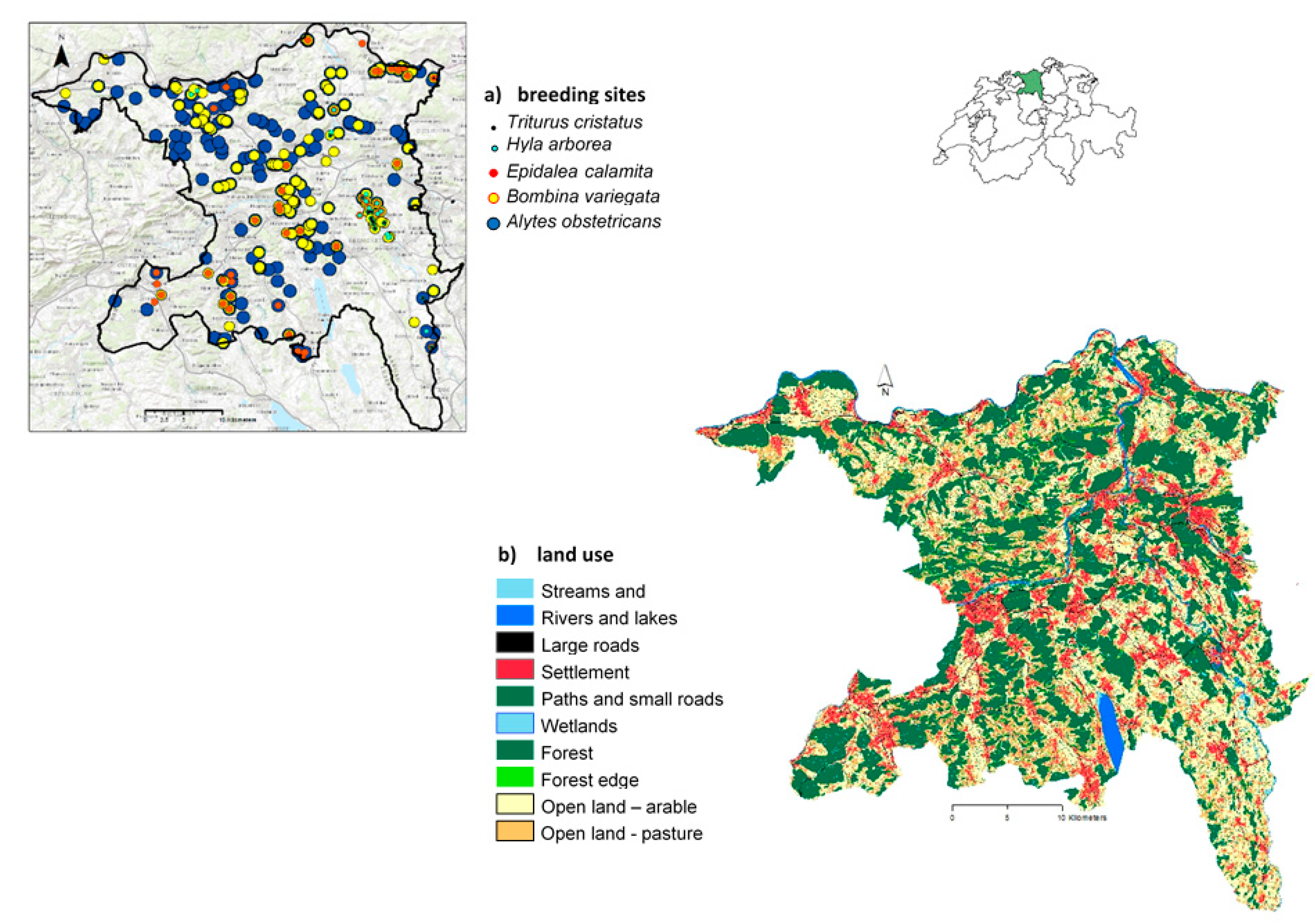
2.1. Study Area
2.2. Study Species
2.3. Landscape Map
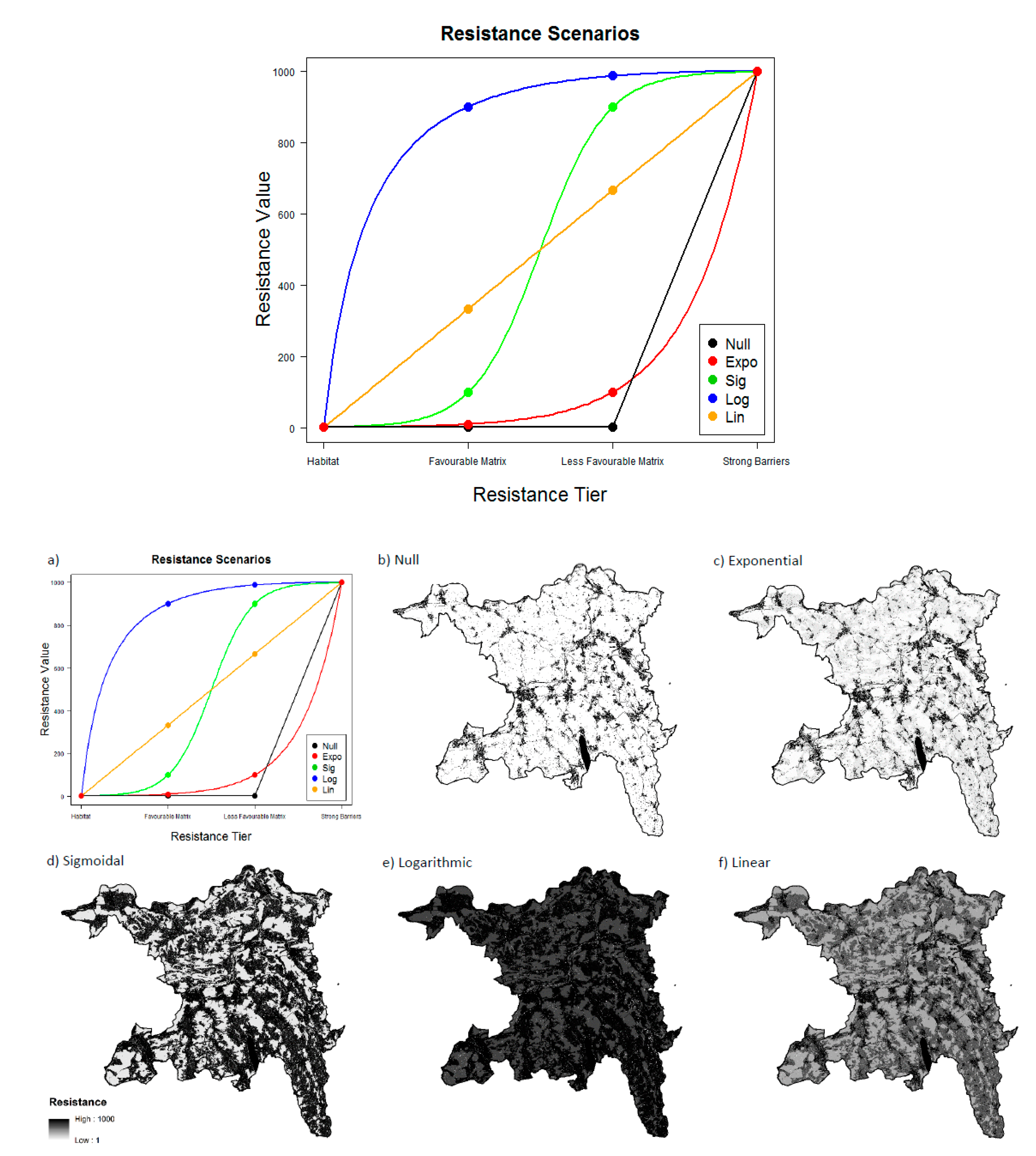
2.4. Landscape Resistance Maps
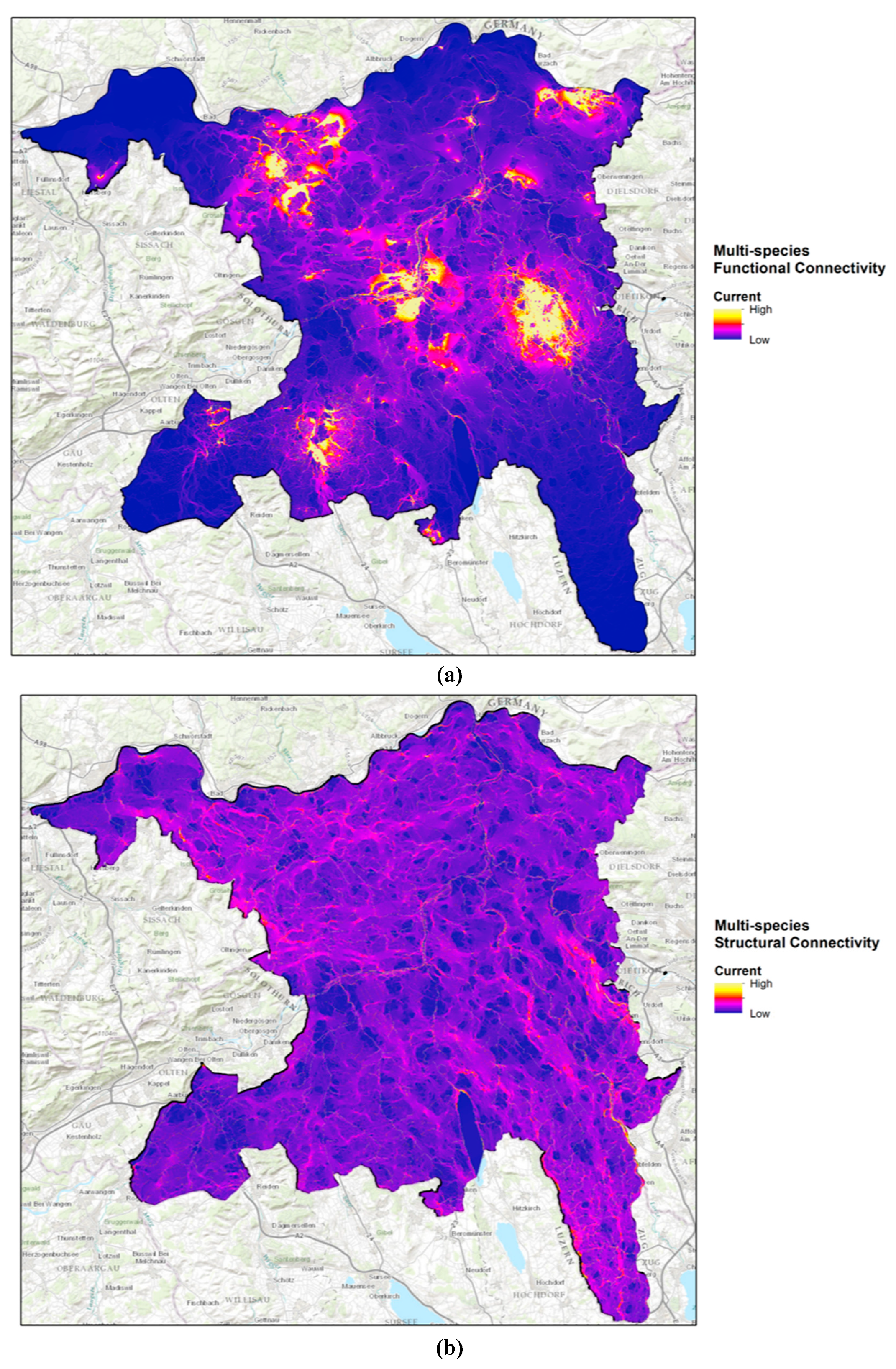
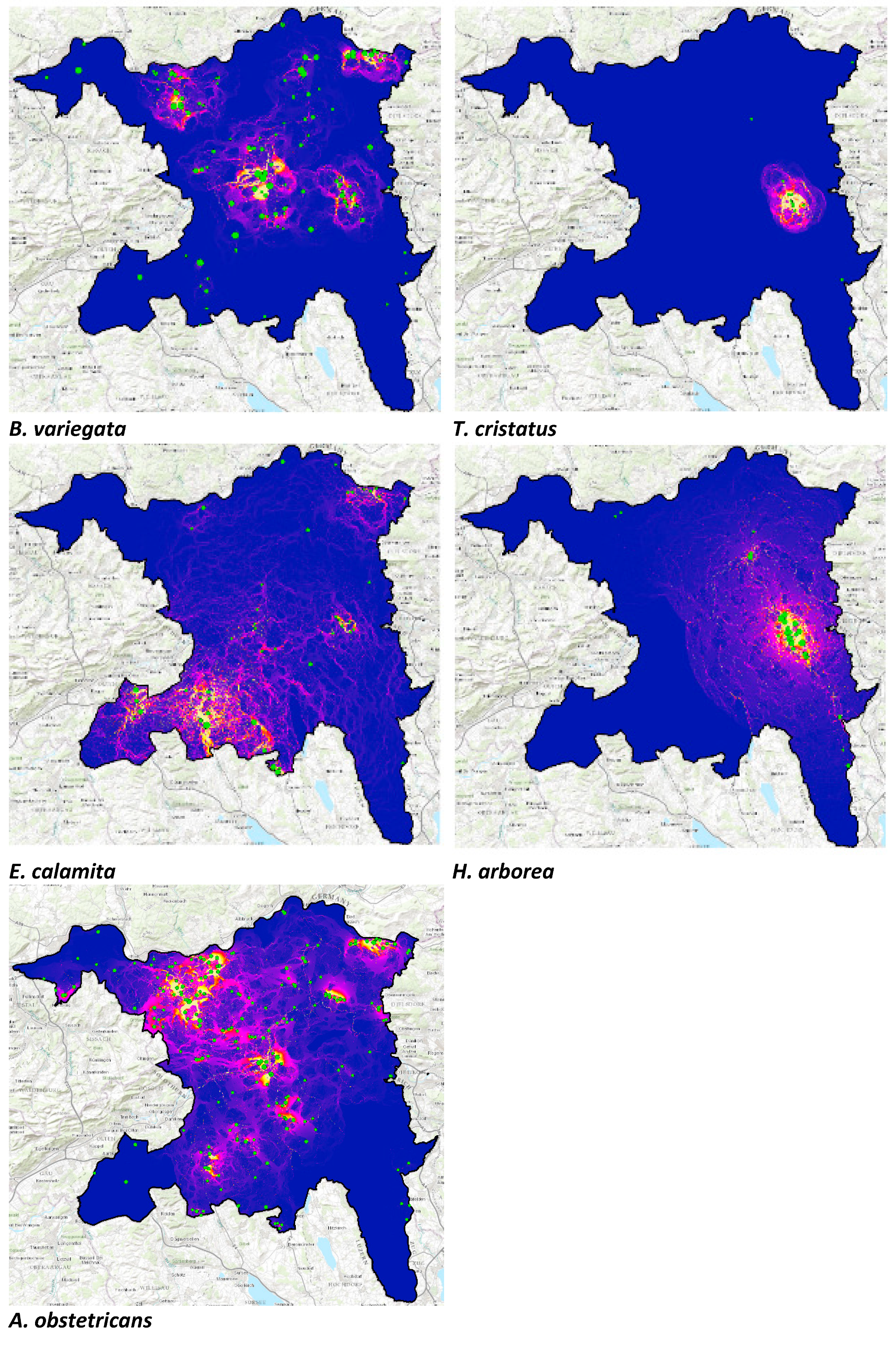
2.5. Modelling Connectivity
2.6. Sensitivity Analysis
3. Results
3.1. Connectivity Analysis
3.2. Resistance Scenarios
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Generating the Landscape Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drawing Order | Land Cover/Land Use | Source | Selection | Type | Buffer | Notes |
|---|---|---|---|---|---|---|
| 1 | Streams Ponds Lake and river margins | TLM_FLIESSGEWAESSER_2015 TLM_BODENBEDECKUNG_2015 TLM_BODENBEDECKUNG_2015 | Objektart = Fliessgewaesser (4) + Verlauf = Oberirdisch (100) Objektart = Stehende Gewaesser (10) + ShapeArea < 600,000 Objektart = Fliessgewaesser (5) + ShapeArea/ShapeLength > 10 Objektart = Stehende Gewaesser (10) + ShapeArea > 600,000 | Polyline Polygon Polygon | 5 m full 10 m full −10 m outside_only | There is an overlap between the two data sources for streams and rivers => all stream polylines that intersected river polygons were considered rivers |
| 2 | Rivers and lakes | TLM_BODENBEDECKUNG_2015 | Objektart = Fliessgewaesser (5) + ShapeArea/ShapeLength > 10 Objektart = Stehende Gewaesser (10) + ShapeArea > 600,000 | Polygon | None | Wetlands and lakes overlap in some locations => these were erased from the lake polygons |
| 3 | Major roads | TLM_STRASSE_2015 | Objektart = Autobahn, Autostrassen, 6 m, 8 m, 10m Strasse (2, 21, 9, 20, 8) | Polyline | 15 m full | |
| 4 | Settlements | TLM_GEBAUDE_FOOTPRINT_2015 | Objektart = Gebaude, etc (1-5) | Polygon | 10 m full | Overlapping features dissolved together |
| 5 | Paths and minor roads | TLM_STRASSE_2015 | Objektart = 3m Strasse, 2 m Wed (11,15) | Polyline | 5 m full | |
| 6 | Wetlands | TLM_BODENBEDECKUNG_2015 | Objektart = Feuchtgebiet (11) | Polygon | 10 m full | |
| 7 | Forest | TLM_BODENBEDECKUNG_2015 | Objektart = Wald, Wald offen (12, 13) | Polygon | None | |
| 8 | Forest margin | TLM_BODENBEDECKUNG_2015 | Objektart = Wald, Wald offen (12, 13) | Polygon | −10 m outside only | |
| 9 | Arable land | Areal Statistik 2004/2009 | Objektart = Ackerland (41) | Raster (100 m cell size) | None | 100 m raster cells resampled down to 10 m |
| 10 | Pasture | Areal Statistik 2004/2009 | Objektart = Naturwiesen (42) and Heimwieden (43), + No Data | Raster (100 m cell size) | None | 100 m raster cells resampled down to 10 m |


References
- Baguette, M.; Blanchet, S.; Legrand, D.; Stevens, V.M.; Turlure, C. Individual dispersal, landscape connectivity and ecological networks. Biol. Rev. 2013, 88, 310–326. [Google Scholar] [CrossRef] [PubMed]
- Cayuela, H.; Lambrey, J.; Vacher, J.P.; Miaud, C. Highlighting the effecs of land-use change on a threatened amphibian in a human-dominated landscape. Popul. Ecol. 2015, 57, 433–443. [Google Scholar] [CrossRef]
- Eterovick, P.C.; Sloss, B.L.; Scalzo, J.A.M.; Alford, R.A. Isolated frogs in a crowded world: Effects of human-caused habitat loss on frog heterozygosity and fluctuating asymmetry. Biol. Conserv. 2016, 195, 52–59. [Google Scholar] [CrossRef]
- Bowne, D.R.; Bowers, M.A. Interpatch movements in spatially structured populations: A literature review. Landsc. Ecol. 2004, 19, 1–20. [Google Scholar] [CrossRef]
- Fahrig, L. Effect of habitat fragmentation on the extinction threshold: A synthesis. Ecol. Appl. 2002, 12, 346–353. [Google Scholar]
- Allen, C.; Gonzales, R.; Parrott, L. Modelling the contribution of ephemeral wetlands to landscape connectivity. Ecol. Model. 2020, 419. [Google Scholar] [CrossRef]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctinos worldwide. Science 2004, 306, 183–186. [Google Scholar] [CrossRef]
- Cushman, S.A.; Shirk, A.J.; Landguth, E.L. Landscape genetics and limiting factors. Conserv. Genet. 2013, 14, 263–274. [Google Scholar] [CrossRef]
- Bolliger, J.; Silbernagel, J. Contribution of connectivity assessments to Green Infrastructure (GI). ISPRS Int. J. Geo Inf. 2020, 9, 212. [Google Scholar] [CrossRef]
- Naidoo, R.; Kilian, J.W.; Du Preez, P.; Beytell, P.; Aschenborn, O.; Taylor, R.D.; Stuart-Hill, G. Evaluating the effectiveness of local- and regional-scale wildlife corridors using quantitative metrics of functional connectivity. Biol. Conserv. 2018, 217, 96–103. [Google Scholar] [CrossRef]
- Milanesi, P.; Holderegger, R.; Caniglia, R.; Fabbri, E.; Galaverni, M.; Randi, E. Expert-based versus habitat-suitability models to develop resistance surfaces in landscape genetics. Oecologia 2017, 183, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Zeller, K.A.; Jennings, M.K.; Vickers, T.W.; Ernest, H.B.; Cushman, S.A.; Boyce, W.M. Are all data types and connectivity models created equal? Validating common connectivity approaches with dispersal data. Divers. Distrib. 2018, 24, 868–879. [Google Scholar] [CrossRef]
- Parks, L.C.; Wallin, D.O.; Cushman, S.A.; McRae, B.H. Landscape-level analysis of mountain goat population connectivity in Washington and southern British Columbia. Conserv. Genet. 2015, 16, 1195–1207. [Google Scholar] [CrossRef]
- Squires, J.R.; DeCesare, N.J.; Olson, L.E.; Kolbe, J.A.; Hebblewhite, M.; Parks, S.A. Combining resource selection and movement behavior to predict corridors for Canada lynx at their southern range periphery. Biol. Conserv. 2013, 157, 187–195. [Google Scholar] [CrossRef]
- Wasserman, T.N.; Cushman, S.A.; Schwartz, M.K.; Wallin, D.O. Spatial scaling and multi-model inference in landscape genetics: Martes americana in northern Idaho. Landsc. Ecol. 2010, 25, 1601–1612. [Google Scholar] [CrossRef]
- Fattebert, J.; Robinson, H.S.; Balme, G.; Slotow, R.; Hunter, L. Structural habitat predicts functional dispersal habitat of a large carnivore: How leopards change spots. Ecol. Appl. 2015, 25, 1911–1921. [Google Scholar] [CrossRef]
- Reed, G.C.; Litvaitis, J.A.; Callahan, C.; Carroll, R.P.; Litvaitis, M.K.; Broman, D.J.A. Modeling landscape connectivity for bobcats using expert-opinion and empirically derived models: How well do they work? Anim. Conserv. 2017, 20, 308–320. [Google Scholar] [CrossRef]
- Marrotte, R.R.; Bowman, J.; Brown, M.G.C.; Cordes, C.; Morris, K.Y.; Prentice, M.B.; Wilson, P.J. Multi-species genetic connectivity in a terrestrial habitat network. Mov. Ecol. 2017, 5, 21. [Google Scholar] [CrossRef]
- Fleishman, E.; Anderson, J.; Dickson, B.G. Single-species and multiple-species connectivity models for large mammals on the Navajo nation. West. N. Am. Nat. 2017, 77, 237–251. [Google Scholar] [CrossRef]
- Bleyhl, B.; Baumann, M.; Griffiths, P.; Heidelberg, A.; Manvelyan, K.; Radeloff, V.C.; Zazanashvili, N.; Kuemmerle, T. Assessing landscape connectivity for large mammals in the Caucasus using Landsat 8 seasonal image composites. Remote Sens. Environ. 2017, 193, 193–203. [Google Scholar] [CrossRef]
- Brodie, J.F.; Giordano, A.J.; Dickson, B.G.; Hebblewhite, M.; Bernard, H.; Mohd-Azlan, J.; Anderson, J.; Ambu, L. Evaluating multispecies landscape connectivity in a threatened tropical mammal community. Conserv. Biol. 2015, 29, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Keller, D.; Holderegger, R.; Van Strien, M.J.; Bolliger, J. How to make landscape genetics beneficial for conservation management? Conserv. Genet. 2015, 16, 503–512. [Google Scholar] [CrossRef]
- Koen, E.L.; Ellington, E.H.; Bowman, J. Mapping landscape connectivity for large spatial extents. Landsc. Ecol. 2019, 34, 2421–2433. [Google Scholar] [CrossRef]
- Clauzel, C.; Bannwarth, C.; Foltete, J.C. Integrating regional-scale connectivity in habitat restoration: An application for amphibian conservation in eastern France. J. Nat. Conserv. 2015, 23, 98–107. [Google Scholar] [CrossRef]
- McRae, B.H.; Dickson, B.G.; Keitt, T.H.; Shah, V.B. Using circuit theory to model connectivity in ecology and conservation. Ecology 2008, 10, 2712–2724. [Google Scholar] [CrossRef]
- McRae, B.H.; Beier, P. Circuit theory predicts gene flow in plant and animal populations. Proc. Natl. Acad. Sci. USA 2007, 104, 19885–19890. [Google Scholar] [CrossRef]
- Dickson, B.G.; Albano, C.M.; Anantharaman, R.; Beier, P.; Fargione, J.; Graves, T.A.; Gray, M.E.; Hall, K.R.; Lawler, J.J.; Leonard, P.B.; et al. Circuit-theory applications to connectivity science and conservation. Conserv. Biol. 2019, 33, 239–249. [Google Scholar] [CrossRef]
- Schmidt, B.R.; Zumbach, S. Rote Liste der Gefährdeten Amphibien der Schweiz, 1st ed.; Bundesamt für Umwelt, Wald und Landschaft (BUWAL); Koordinationsstelle für Amphibien- und Reptilienschutz in der Schweiz (KARCH): Bern, Switzerland, 2005; p. 48. [Google Scholar]
- Meier, C.; Schelbert, B. Amphibienschutzkonzept Kanton Aargau. Mitt. Aargauer Nat. Ges. 1999, 35, 41–69. [Google Scholar]
- Le Lay, G.; Angelone, S.; Flory, C.; Holderegger, R.; Bolliger, J. Increasing pond density to maintain a patchy habitat network of the European tree frog (Hyla arborea). J. Herpetol. 2015, 49, 217–221. [Google Scholar] [CrossRef]
- Frei, M.; Csencsics, C.; Brodbeck, S.; Schweizer, E.; Bühler, C.; Gugerli, F.; Bolliger, J. Combining landscape genetics, radio-tracking and long-term monitoring to derive management implications for Natterjack toads (Epidalea calamita) in agricultural landscapes. J. Nat. Conserv. 2016, 32, 22–34. [Google Scholar] [CrossRef]
- Smith, M.A.; Green, D.M. Dispersal and the metapopulation paradigm in amphibian ecology and conservation: Are all amphibian populations metapopulations? Ecography 2005, 28, 110–128. [Google Scholar] [CrossRef]
- Primus, J. Disperal and Migration in Yellow-Bellied Toads, Bombina Variegata; University of Vienna: Vienna, Austria, 2013. [Google Scholar]
- Baker, J.; Beebee, T.; Buckley, J.; Gent, A.; Orchard, D. Amphibian Habitat Management Handbook; Amphibian and Reptile Conservation: Bournemouth, UK, 2011. [Google Scholar]
- swissTLM 3D. Vectorized National Map 1:25’000, Bundesamt für Landestopographie; Swisstopo: Wabern, Switzerland, 2015. [Google Scholar]
- BfS, S. Land Use in Switzerland: Results of the Swiss Land Use Statistics; Federal Statistical Office: Neuchatel, Switzerland, 2013.
- Koen, E.L.; Bowman, J.; Sadowski, C.; Walpole, A.A. Landscape connectivity for wildlife: Development and validation of multispecies linkage maps. Methods Ecol. Evol. 2014, 5, 626–633. [Google Scholar] [CrossRef]
- Rayfield, B.; Fortin, M.J.; Fall, A. The sensitivity of least-cost habitat graphs to relative cost surface values. Landsc. Ecol. 2010, 25, 519–532. [Google Scholar] [CrossRef]
- Braaker, S.; Moretti, M.; Boesch, R.; Ghazoul, J.; Obrist, M.K.; Bontadina, F. Assessing habitat connectivity for ground-dwelling animals in an urban environment. Ecol. Appl. 2014, 24, 1583–1595. [Google Scholar] [CrossRef] [PubMed]
- Nowakowski, A.J.; Veiman-Echeverria, M.; Kurz, D.J.; Donnelly, M.A. Evaluating connectivity for tropical amphibians using empirically derived resistance surfaces. Ecol. Appl. 2015, 25, 928–942. [Google Scholar] [CrossRef]
- McRae, B.H.; Schumaker, N.; McKane, R.; Busing, R.; Solomon, A.; Burdick, C. A multi-model framework for simulating wildlife population response to land use and climate change. Ecol. Model. 2008, 219, 77–79. [Google Scholar] [CrossRef]
- Walpole, A.A.; Bowman, J.; Murray, D.L.; Wilson, P.J. Functional connectivity of lynx at their southern range periphery in Ontario, Canada. Landsc. Ecol. 2012, 27, 761–773. [Google Scholar] [CrossRef]
- Pelletier, D.; Clark, M.; Anderson, M.G.; Rayfield, B.; Wulder, M.A.; Cardille, J.A. Applying Circuit Theory for Corridor Expansion and Management at Regional Scales: Tiling, Pinch Points, and Omnidirectional Connectivity. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Beier, C.M. Influence of Political Opposition and Compromise on Conservation Outcomes in the Tongass National Forest, Alaska. Conserv. Biol. 2008, 22, 1485–1496. [Google Scholar] [CrossRef]
- Breckheimer, I.; Haddad, N.M.; Morris, W.F.; Trainor, A.M.; Fields, W.R.; Jobe, R.T.; Hudgens, B.R.; Moody, A.; Walters, J.R. Defining and evaluating the umbrella species concept for conserving and restoring landscape connectivity. Conserv. Biol. 2014, 28, 1584–1593. [Google Scholar] [CrossRef]
- Mikolas, M.; Tejkal, M.; Kuemmerle, T.; Griffiths, P.; Svoboda, M.; Hlasny, T.; Leitao, P.J.; Morrissey, R.C. Forest management impacts on capercaillie (Tetrao urogallus) habitat distribution and connectivity in the Carpathians. Landsc. Ecol. 2017, 32, 163–179. [Google Scholar] [CrossRef]
- Beier, P.; Majka, D.R.; Newell, S.L. Uncertainty analysis of least-cost modeling for designing wildlife linkages. Ecol. Appl. 2009, 19, 2067–2077. [Google Scholar] [CrossRef] [PubMed]
- Dondina, O.; Orioli, V.; Chiatante, G.; Bani, L. Practical insights to select focal species and design priority areas for conservation. Ecol. Indic. 2020, 108. [Google Scholar] [CrossRef]
- Meurant, M.; Gonzalez, A.; Doxa, A.; Albert, C.H. Selecting surrogate species for connectivity conservation. Biol. Conserv. 2018, 227, 326–334. [Google Scholar] [CrossRef]
- Peterman, W.E.; Connette, G.M.; Semlitsch, R.D.; Eggert, L.S. Ecological resistance surfaces predict fine-scale genetic differentiation in a terrestrial woodland salamander. Mol. Ecol. 2014, 23, 2402–2413. [Google Scholar] [CrossRef] [PubMed]
- Frei, M.; Csencsics, D.; Bühler, C.; Gugerli, F.; Bolliger, J. Wie gut vernetzt sind Vorkommen der Kreuzkröte in einer landwirtschaftlich geprägten Landschaft? N. L. Inside 2014, 4, 16–20. [Google Scholar]
- Abrahms, B.; DiPietro, D.; Graffis, A.; Hollander, A. Managing biodiversity under climate change: Challenges, frameworks, and tools for adaptation. Biodivers. Conserv. 2017, 26, 2277–2293. [Google Scholar] [CrossRef]
- Bolliger, J.; Keller, D.; Holderegger, R. When landscape variables do not explain migration rates: An example from an endangered dragonfly (Leucorrhinia caudalis). Eur. J. Entomol. 2011, 108, 327–330. [Google Scholar] [CrossRef]
- Keller, D.; Van Strien, M.J.; Ghazoul, J.; Holderegger, R. Landscape Genetics of Insects in Intensive Agriculture: New Ecological Insights; Swiss Federal Research Institute WSL: Birmensdorf, Switzerland, 2012.
- Keller, D.; Van Strien, M.J.; Holderegger, R. Do landscape barriers affect functional connectivity of populations of an endangered damselfly? Freshw. Biol. 2012, 57, 1373–1384. [Google Scholar] [CrossRef]
- Bolliger, J.; Lander, T.; Balkenhol, N. Landscape genetics since 2003: Status, challenges and future directions. Landsc. Ecol. 2014, 29, 361–366. [Google Scholar] [CrossRef]
- Holderegger, R.; Balkenhol, N.; Bolliger, J.; Engler, J.O.; Gugerli, F.; Hochkirch, A.; Nowak, C.; Segelbacher, G.; Widmer, A.; Zachos, F.E. Conservation genetics: Linking science with practice. Mol. Ecol. 2019, 28, 3848–3856. [Google Scholar] [CrossRef] [PubMed]
- Sinsch, U.; Oromi, N.; Miaud, C.; Denton, J.; Sanuy, D. Connectivity of local amphibian populations: Modelling the migratory capacity of radio-tracked Natterjack toads. Anim. Conserv. 2012, 15, 388–396. [Google Scholar] [CrossRef]
- Wang, J.X.; Banzhaf, E. Towards a better understanding of Green Infrastructure: A critical review. Ecol. Indic. 2018, 85, 758–772. [Google Scholar] [CrossRef]
- Snäll, T.; Lehtomäki, J.; Arponen, A.; Elith, J.; Moilanen, A. Green infrastructure design based on spatial conservation prioritization and modeling of biodiversity features and ecosystem services. Environ. Manag. 2016, 57, 251–256. [Google Scholar]
- European Commission. Green Infrastructure (GI)—Enhancing Europe’s Natural Capital; European Commission: Brussels, Belgium, 2013; p. 149. [Google Scholar]



| Estimated Breeding Population Sizes (# Individuals) | A. obstetricans | B. variegata | B. calamita | H. arborea | T. cristatus |
|---|---|---|---|---|---|
| 1-5 | 146 | 58 | 30 | 6 | 12 |
| 6-20 | 58 | 38 | 9 | 8 | 0 |
| 21-50 | 7 | 19 | 5 | 9 | 0 |
| 50+ | 0 | 11 | 1 | 3 | 0 |
| Total | 211 | 126 | 45 | 26 | 12 |
| Maximum Dispersal Distance | 2 km | 1.5 km | 5 km | 12 km | 1.5 km |
| Resistance Tier | A. obstetricans | B. variegata | B. calamita | H. arborea | T. cristatus |
|---|---|---|---|---|---|
| Habitat | 1 | 1, 7 | 5, 9 | 1, 6 | 1, 6 |
| Favourable Matrix | 5, 7, 8 | 5, 6, 8 | 1, 6, 8, 10 | 8, 10 | 7, 8 |
| Less Favourable Matrix | 6, 9, 10 | 9, 10 | 7 | 5, 7, 9 | 5, 9, 10 |
| Strong Barrier | 2, 3, 4 | 2, 3, 4 | 2, 3, 4 | 2, 3, 4 | 2, 3, 4 |
| Resistance Tier | Null | Exponential | Sigmoidal | Logarithmic | Linear |
|---|---|---|---|---|---|
| Habitat | 1 | 1 | 1 | 1 | 1 |
| Favourable Matrix | 1 | 10 | 100 | 900 | 333 |
| Less Favourable Matrix | 1 | 100 | 900 | 990 | 666 |
| Strong Barrier | 1000 | 1000 | 1000 | 1000 | 1000 |
| Null | Exponential | Sigmoidal | Logarithmic | Linear | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Null | 100 | 0 | 50 | 8 | 41 | 10 | 42 | 14 | 44 | 14 |
| Exponential | - | 100 | 0 | 77 | 5 | 60 | 9 | 68 | 7 | |
| Sigmoidal | - | - | 100 | 0 | 74 | 12 | 82 | 8 | ||
| Logarithmic | - | - | - | 100 | 0 | 90 | 4 | |||
| Linear | - | - | - | - | 100 | 0 | ||||
| Null | Exponential | Sigmoidal | Logarithmic | Linear | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Null | 1.00 | 0.00 | 0.27 | 0.23 | 0.23 | 0.13 | 0.15 | 0.13 | 0.17 | 0.15 |
| Exponential | - | 1.00 | 0.00 | 0.56 | 0.33 | 0.34 | 0.29 | 0.41 | 0.26 | |
| Sigmoidal | - | - | 1.00 | 0.00 | 0.43 | 0.35 | 0.46 | 0.36 | ||
| Logarithmic | - | - | - | 1.00 | 0.00 | 0.46 | 0.31 | |||
| Linear | - | - | - | - | 1.00 | 0.00 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Churko, G.; Kienast, F.; Bolliger, J. A Multispecies Assessment to Identify the Functional Connectivity of Amphibians in a Human-Dominated Landscape. ISPRS Int. J. Geo-Inf. 2020, 9, 287. https://doi.org/10.3390/ijgi9050287
Churko G, Kienast F, Bolliger J. A Multispecies Assessment to Identify the Functional Connectivity of Amphibians in a Human-Dominated Landscape. ISPRS International Journal of Geo-Information. 2020; 9(5):287. https://doi.org/10.3390/ijgi9050287
Chicago/Turabian StyleChurko, Gregory, Felix Kienast, and Janine Bolliger. 2020. "A Multispecies Assessment to Identify the Functional Connectivity of Amphibians in a Human-Dominated Landscape" ISPRS International Journal of Geo-Information 9, no. 5: 287. https://doi.org/10.3390/ijgi9050287
APA StyleChurko, G., Kienast, F., & Bolliger, J. (2020). A Multispecies Assessment to Identify the Functional Connectivity of Amphibians in a Human-Dominated Landscape. ISPRS International Journal of Geo-Information, 9(5), 287. https://doi.org/10.3390/ijgi9050287