RSM–GA Based Optimization of Bacterial PHA Production and In Silico Modulation of Citrate Synthase for Enhancing PHA Production


 ,
,  ,
,
Abstract
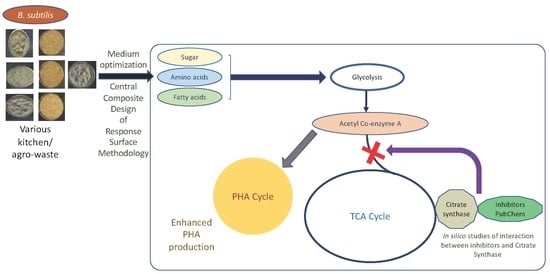
1. Introduction
2. Materials and Methods
2.1. Microorganism and Fermentation Condition
2.2. Selection of Production Medium
2.3. Extraction and Quantification of PHA
2.4. Selection of Carbon and Nitrogen Source
2.5. RSM-Based Optimization of PHA Production
2.6. GA Optimization
2.7. Characterization of PHAs
2.7.1. (i) FTIR Analysis of PHAs
2.7.2. Thin Layer Chromatography of PHAs
2.7.3. 1H-NMR
2.7.4. X-Ray Diffraction (XRD) Analysis
2.8. In Silico Modulation of Citrate Synthase for the Enhancement of PHA Production
2.8.1. Target Selection and Preparation
2.8.2. Library Preparation of Ligands
2.8.3. Molecular Docking
3. Results and Discussion
3.1. Effect of Various Media on PHA Production
3.2. Effect of Carbon and Nitrogen Sources Derived from Various Kitchen-/Agro-Waste on PHA Production
3.3. Statistical Optimization
3.4. GA-Based Optimization
3.5. PHA Characterization
3.6. Molecular Docking Studies of Citrate Synthase
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kumar, R.; Sharma, S.K. Biodegradation of Hibiscus mucilage and potato starch blended with LDPE: A comparative study. Int. J. Recent Sci. Res. 2018, 9, 26137–26139. [Google Scholar] [CrossRef]
- Reddy, C.S.K.; Ghai, R.; Kalia, V. Polyhydroxyalkanoates: An overview. Bioresour. Technol. 2003, 87, 137–146. [Google Scholar] [CrossRef]
- Kumar, A.; Srivastava, J.K.; Mallick, N.; Singh, A.K. Commercialization of bacterial cell factories for the sustainable production of polyhydroxyalkanoate thermoplastics: Progress and prospects. Recent Pat. Biotechnol. 2015, 9, 4–21. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yang, J.; Loh, X.J. Polyhydroxyalkanoates: Opening doors for a sustainable future. NPG Asia Mater. 2016, 8, e265. [Google Scholar] [CrossRef]
- Umesh, M.; Mani, V.M.; Thazeem, B.; Preethi, K. Statistical Optimization of Process Parameters for Bioplastic (PHA) Production by Bacillus subtilis NCDC0671 Using Orange Peel-Based Medium. Iran. J. Sci. Technol. Trans. A Sci. 2018, 42, 1947–1955. [Google Scholar] [CrossRef]
- Raza, Z.A.; Abid, S.; Banat, I.M. Polyhydroxyalkanoates: Characteristics, production, recent developments and applications. Int. Biodeterior. Biodegrad. 2018, 126, 45–56. [Google Scholar] [CrossRef]
- Myung, J.; Flanagan, J.C.; Waymouth, R.M.; Criddle, C.S. Expanding the range of polyhydroxyalkanoates synthesized by methanotrophic bacteria through the utilization of omega-hydroxyalkanoate co-substrates. AMB Express 2017, 7, 118. [Google Scholar] [CrossRef]
- Koller, M. Advances in Polyhydroxyalkanoate (PHA) Production. Bioengineering 2017, 4, 88. [Google Scholar] [CrossRef]
- Brigham, C.J.; Riedel, S.L. The Potential of Polyhydroxyalkanoate Production from Food Wastes. Appl. Food Biotechnol. 2018, 6, 7–18. [Google Scholar]
- Pratt, S.; Vandi, L.J.; Gapes, D.; Werker, A.; Oehmen, A.; Laycock, B. Polyhydroxyalkanoate (PHA) Bioplastics from Organic Waste. In Biorefinery; Springer: Cham, Switzerland, 2019; pp. 615–638. [Google Scholar]
- Chee, J.Y.; Yoga, S.S.; Lau, N.S.; Ling, S.C.; Abed, R.M.; Sudesh, K. Bacterially produced polyhydroxyalkanoate (PHA): Converting renewable resources into bioplastics. Curr. Res. Technol. Educ. Top. Appl. Microbiol. Microb. Biotechnol. 2010, 2, 1395–1404. [Google Scholar]
- Shivakumar, S. Polyhydroxybutyrate (PHB) production using agro-industrial residue as substrate by Bacillus thuringiensis IAM 12077. Int. J. Chem. Tech. Res. 2012, 4, 1158–1162. [Google Scholar]
- Gowda, V.; Shivakumar, S. Agrowaste-based Polyhydroxyalkanoate (PHA) production using hydrolytic potential of Bacillus thuringiensis IAM 12077. Braz. Arch. Biol. Technol. 2014, 57, 55–61. [Google Scholar] [CrossRef]
- Srivastava, A.; Singh, V.; Haque, S.; Pandey, S.; Mishra, M.; Jawed, A.; Shukla, P.K.; Singh, P.K. Tripathi CK. Response surface methodology-genetic algorithm based medium optimization, purification, and characterization of cholesterol oxidase from Streptomyces rimosus. Sci. Rep. 2018, 8, 10913. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.T.; Chen, W.C.; Chou, C.C. Medium improvement by orthogonal array designs for cholesterol oxidase production by Rhodococcus equi No. 23. Process Biochem. 1997, 32, 697–703. [Google Scholar] [CrossRef]
- El-Naggar, N.E.-A.; El-Shweihy, N.M.; El-Ewasy, S.M. Identification and statistical optimization of fermentation conditions for a newly isolated extracellular cholesterol oxidase-producing Streptomyces cavourensis strain NEAE-42. BMC Microbiol. 2016, 16, 217. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Tripathi, C.K.M. Production and statistical optimization of a novel olivanic acid by Streptomyces olivaceus MTCC 6820. Process Biochem. 2008, 43, 1313–1317. [Google Scholar] [CrossRef]
- Singh, V.; Haque, S.; Niwas, R.; Srivastava, A.; Pasupuleti, M.; Tripathi, C.K.M. Strategies for Fermentation Medium Optimization: An In-Depth Review. Front. Microbiol. 2017, 7, 2087. [Google Scholar] [CrossRef]
- Tripathi, C.K.M.; Khan, M.; Praveen, V.; Khan, S.; Srivastava, A. Enhanced antibiotic production by Streptomyces sindenensis using artificial neural networks coupled with genetic algorithm and Nelder-Mead Downhill Simplex. J. Microbiol. Biotechnol. 2012, 22, 939–946. [Google Scholar] [CrossRef]
- Heinrich, D.; Madkour, M.H.; Al-Ghamdi, M.A.; Shabbaj, I.I.; Steinbüchel, A. Large scale extraction of poly (3-hydroxybutyrate) from Ralstonia eutropha H16 using sodium hypochlorite. AMB Express 2012, 2, 59. [Google Scholar] [CrossRef]
- Law, J.H.; Slepecky, R.A. Assay of poly-β-hydroxybutyric acid. J. Bacteriol. 1961, 82, 33–36. [Google Scholar]
- Salgaonkar, B.B.; Mani, K.; Braganca, J.M. Characterization of polyhydroxyalkanoates accumulated by a moderately halophilic salt pan isolate Bacillus megaterium strain H 16. J. Appl. Microbiol. 2013, 114, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.K.; Ongpipattanakul, B. Au-Yeung, J. Aggregation of rhDNase occurred during the compression of KBr pellets used for FTIR spectroscopy. Pharm. Res. 1996, 13, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Panda, B.; Sharma, L.; Singh, A.K.; Mallick, N. Thin layer chromatographic detection of poly-β-hydroxybutyrate (PHB) and poly-β-hydroxyvalerate (PHV) in cyanobacteria. Indian J. Biotechnol. 2008, 7, 230–234. [Google Scholar]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Production of polyhydroxyalkanoates (PHAs) by Bacillus subtilis and Escherichia coli grown on cane molasses fortified with ethanol. Braz. Arch. Biol. Technol. 2014, 57, 145–154. [Google Scholar] [CrossRef]
- Porras, M.A.; Villar, M.A.; Cubitto, M.A. Improved intracellular PHA determinations with novel spectrophotometric quantification methodologies based on Sudan black dye. J. Microbiol. Methods 2018, 148, 1–11. [Google Scholar] [CrossRef]
- Bhuwal, A.K.; Singh, G.; Aggarwal, N.K.; Goyal, V.; Yadav, A. Isolation and screening of polyhydroxyalkanoates producing bacteria from pulp, paper, and cardboard industry wastes. Int. J. Biomater. 2013, 2013, 752821. [Google Scholar] [CrossRef]
- Kulkarni, S.O.; Kanekar, P.P.; Jog, J.P.; Sarnaik, S.S.; Nilegaonkar, S.S. Production of copolymer, poly (hydroxybutyrate-co-hydroxyvalerate) by Halomonas campisalis MCM B-1027 using agro-wastes. Int. J. Biol. Macromol. 2015, 72, 784–789. [Google Scholar] [CrossRef]
- Shah, K.R. Optimization and production of Polyhydroxybutarate (PHB) by Bacillus subtilis G1S1 from soil. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 377–387. [Google Scholar]
- Singh, V.; Khan, M.; Khan, S.; Tripathi, C.K. Optimization of actinomycin V production by Streptomyces triostinicus using artificial neural network and genetic algorithm. Appl. Microbiol. Biotechnol. 2009, 82, 379–385. [Google Scholar] [CrossRef]
- Hong, K.; Sun, S.; Tian, W.; Chen, G.Q.; Huang, W. A rapid method for detecting bacterial polyhydroxyalkanoates in intact cells by Fourier transform infrared spectroscopy. Appl. Microbiol. Biotechnol. 1999, 51, 523–526. [Google Scholar] [CrossRef]
- Nair, A.M.; Annamalai, K.; Kannan, S.K.; Kuppusamy, S. Utilization of sugarcane molasses for the production ofpolyhydroxyalkanoates using Bacillus subtilis 2014. Malaya J. Biosci. 2013, 1, 24–30. [Google Scholar]
- Vega-Castro, O.; Contreras-Calderon, J.; León, E.; Segura, A.; Arias, M.; Pérez, L.; Sobral, P.J. Characterization of a polyhydroxyalkanoate obtained from pineapple peel waste using Ralsthonia eutropha. J. Biotechnol. 2016, 231, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Tan, G.Y.; Chen, C.L.; Li, L.; Ge, L.; Wang, L.; Razaad, I.M.; Li, Y.; Zhao, L.; Mo, Y.; Wang, J.Y. Start a research on biopolymer polyhydroxyalkanoate (PHA): A review. Polymers 2014, 6, 706–754. [Google Scholar] [CrossRef]
- Pillai, A.B.; Kumar, A.J.; Kumarapillai, H. Enhanced production of poly (3-hydroxybutyrate) in recombinant Escherichia coli and EDTA–microwave-assisted cell lysis for polymer recovery. AMB Express 2018, 8, 142. [Google Scholar] [CrossRef] [PubMed]
- Sedlacek, P.; Slaninova, E.; Enev, V.; Koller, M.; Nebesarova, J.; Marova, I.; Hrubanova, K.; Krzyzanek, V.; Samek, O.; Obruca, S. What keeps polyhydroxyalkanoates in bacterial cells amorphous? A derivation from stress exposure experiments. Appl. Microbiol. Biotechnol. 2019, 103, 1905–1917. [Google Scholar] [CrossRef] [PubMed]
- Alashwal, B.Y.; Bala, M.S.; Gupta, A.; Sharma, S.; Mishra, P. Improved properties of keratin-based bioplastic film blended with microcrystalline cellulose: A comparative analysis. J. King Saud Univ. Sci. 2019. [Google Scholar] [CrossRef]
- Mohapatra, S.; Sarkar, B.; Samantaray, D.P.; Daware, A.; Maity, S.; Pattnaik, S.; Bhattacharjee, S. Bioconversion of fish solid waste into PHB using Bacillus subtilis based submerged fermentation process. Environ. Technol. 2017, 38, 3201–3208. [Google Scholar] [CrossRef]
- Man, M.Q.; Yang, B.; Elias, P.M. Benefits of hesperidin for cutaneous functions. Evid. Based Complement. Altern. Med. 2019, 2019, 2676307. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No. | Cell Dry Weight (g/L) | PHAs (g) | % Conversion of PHAs | |
|---|---|---|---|---|
| Carbon Source | ||||
| 1 | Mixed fruit peels | 8.5 | 4.9 | 57.64 |
| 2 | Mixed vegetable peels | 9.8 | 6.2 | 63.27 |
| 3 | Green pea shells | 11.26 | 8.77 | 77.89 |
| 4 | Muskmelon peels | 9.98 | 7.84 | 78.56 |
| 5 | Watermelon rind | 16.5 | 12.97 | 78.61 |
| 6 | Papaya peels | 15.0 | 11.65 | 77.67 |
| 7 | Orange peels | 19.39 | 9.68 | 49.93 |
| Nitrogen Source | ||||
| 1 | Peptone | 16.5 | 12.97 | 78.61 |
| 2 | Pulse peel | 19.51 | 13.5 | 69.20 |
| 3 | Beef extract | 18.85 | 11.5 | 61.01 |
| 4 | Yeast extract | 12.02 | 9.45 | 78.62 |
| Runs | Carbon Concentration (g/100 mL) | Nitrogen Concentration (g/100 mL) | PHA Content (g) | ||
|---|---|---|---|---|---|
| Observed | Predicted | Residual | |||
| 1 | 2 | 0.1 | 27.221 | 23.095 | 4.125 |
| 2 | 2 | 0.3 | 27.928 | 25.881 | 2.046 |
| 3 | 6 | 0.1 | 37.996 | 35.613 | 2.382 |
| 4 | 6 | 0.3 | 36.052 | 35.748 | 0.303 |
| 5 | 4 | 0.2 | 33.124 | 35.966 | −2.842 |
| 6 | 0 | 0.2 | 2.25 | 4.228 | −1.978 |
| 7 | 8 | 0.2 | 26.377 | 26.612 | −0.235 |
| 8 | 4 | 0 | 29.38 | 31.526 | −2.146 |
| 9 | 4 | 0.4 | 34.38 | 34.447 | −0.067 |
| 10 | 4 | 0.2 | 34.38 | 35.966 | −1.586 |
| Source | SS | df | MS | F-value | Prob (p) |
|---|---|---|---|---|---|
| Whole model | 896.0238 | 5 | 179.2048 | 19.72049 | 0.006392 |
| Residual | 36.34894 | 4 | 9.087235 |
| Effect | Var3 Param. | Var3 Std. Err | Var3 t | Var3 p |
|---|---|---|---|---|
| Intercept | −2.8622 | 8.43462 | −0.33934 | 0.751425 |
| Var1 | 13.7336 | 2.43016 | 5.65131 | 0.004830 |
| Var1^2 | −1.2841 | 0.20846 | −6.15993 | 0.003525 |
| Var2 | 50.3486 | 48.60319 | 1.03591 | 0.358759 |
| Var2^2 | −74.4777 | 83.38407 | −0.89319 | 0.422238 |
| Var1*Var2 | −3.3137 | 8.49142 | −0.39025 | 0.716244 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rao, A.; Haque, S.; El-Enshasy, H.A.; Singh, V.; Mishra, B.N. RSM–GA Based Optimization of Bacterial PHA Production and In Silico Modulation of Citrate Synthase for Enhancing PHA Production. Biomolecules 2019, 9, 872. https://doi.org/10.3390/biom9120872
Rao A, Haque S, El-Enshasy HA, Singh V, Mishra BN. RSM–GA Based Optimization of Bacterial PHA Production and In Silico Modulation of Citrate Synthase for Enhancing PHA Production. Biomolecules. 2019; 9(12):872. https://doi.org/10.3390/biom9120872
Chicago/Turabian StyleRao, Apoorva, Shafiul Haque, Hesham A. El-Enshasy, Vineeta Singh, and Bhartendu Nath Mishra. 2019. "RSM–GA Based Optimization of Bacterial PHA Production and In Silico Modulation of Citrate Synthase for Enhancing PHA Production" Biomolecules 9, no. 12: 872. https://doi.org/10.3390/biom9120872
APA StyleRao, A., Haque, S., El-Enshasy, H. A., Singh, V., & Mishra, B. N. (2019). RSM–GA Based Optimization of Bacterial PHA Production and In Silico Modulation of Citrate Synthase for Enhancing PHA Production. Biomolecules, 9(12), 872. https://doi.org/10.3390/biom9120872