
A Natural Alkaloid, 6-Hydroxymethyldihydronitidine, Suppresses Tumor Progression by Co-Regulating Apoptosis, Ferroptosis, and FAK Pathways

Abstract

1. Introduction
2. Materials and Methods
2.1. Materials and Cell Culture
2.2. Extraction and Purification of 6-HMDN
2.3. MTT Assay
2.4. Zebrafish Husbandry
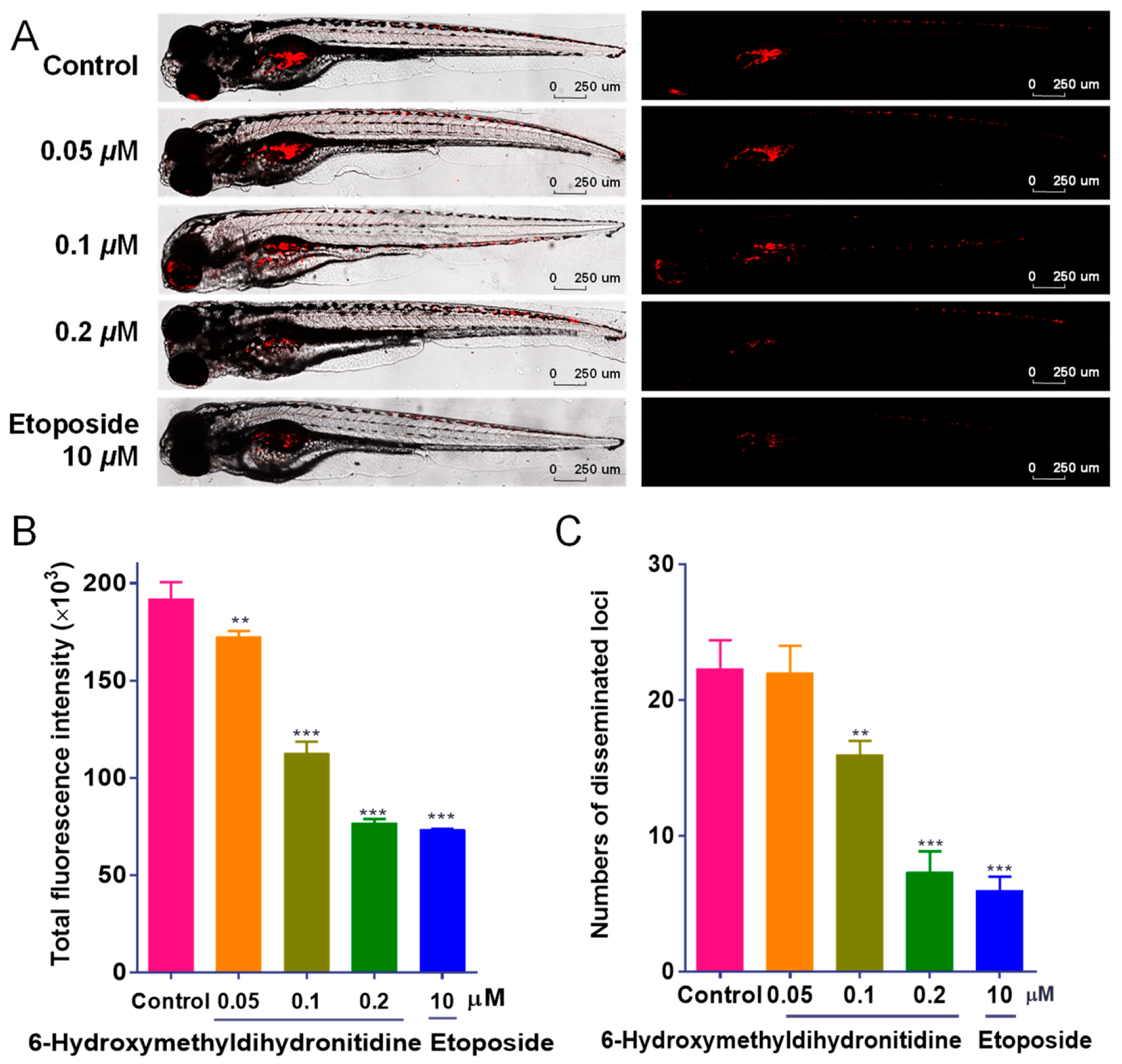
2.5. In Vivo Anti-Tumor Assay Using a Zebrafish Xenograft Model
2.6. Apoptosis Assay
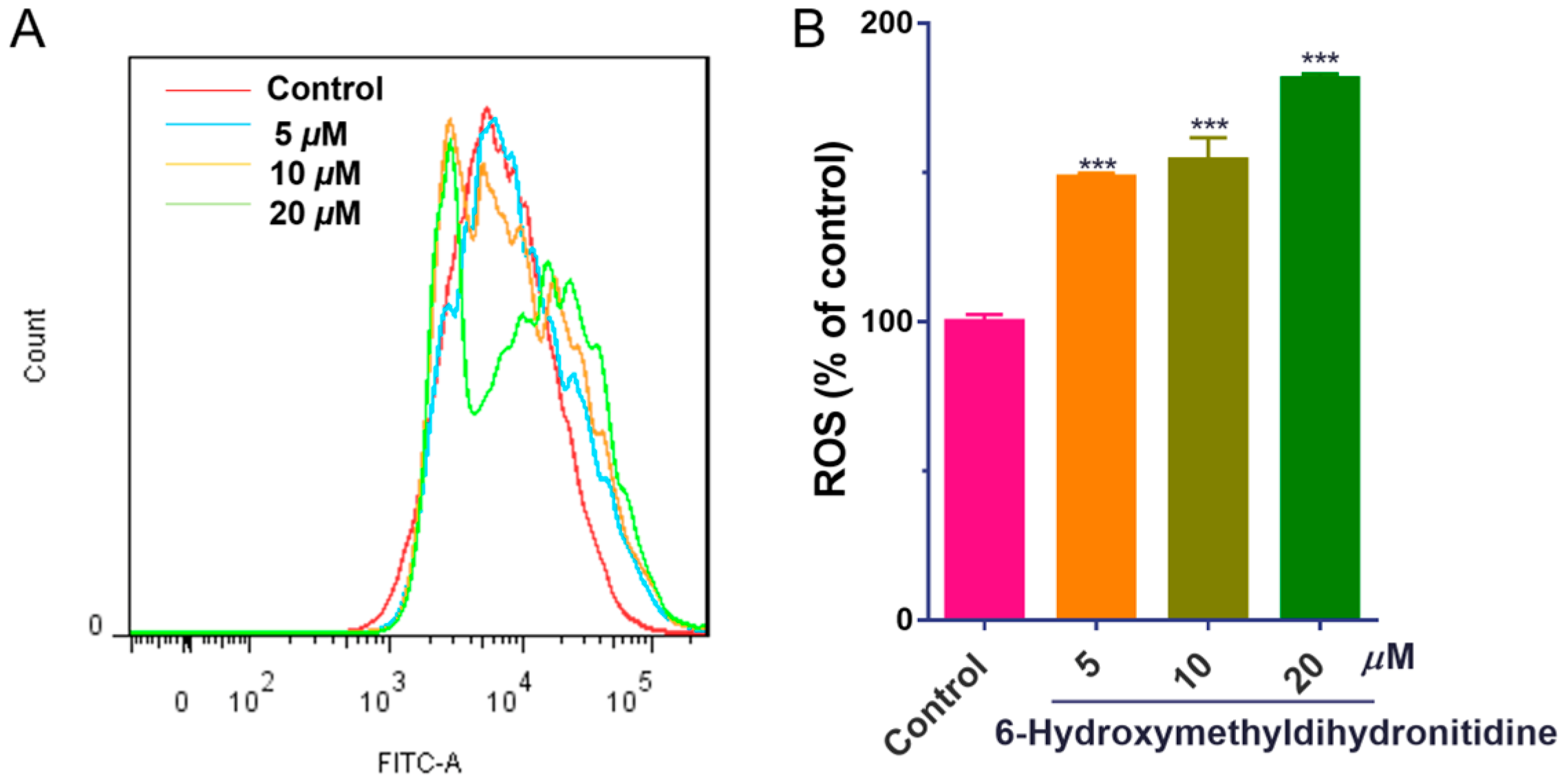
2.7. Measurement of ROS Production
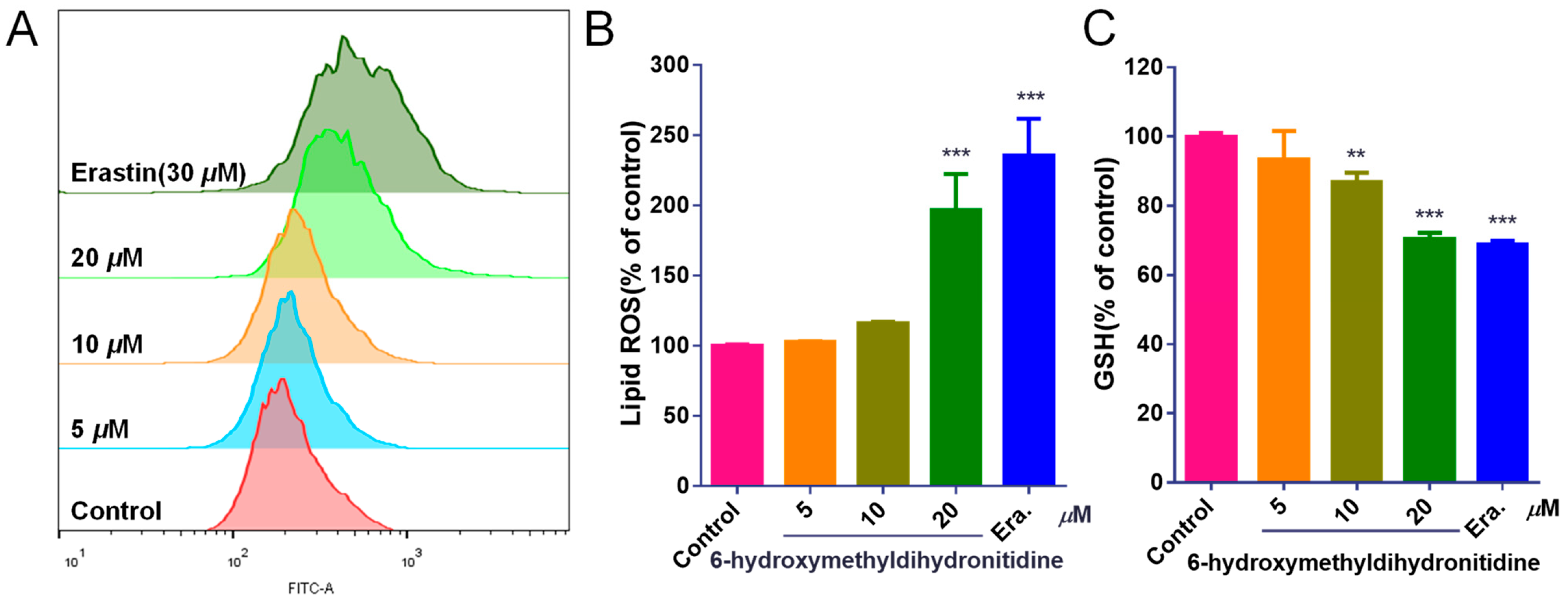
2.8. Measurement of Glutathione Content
2.9. Measurement of Lipid ROS Levels
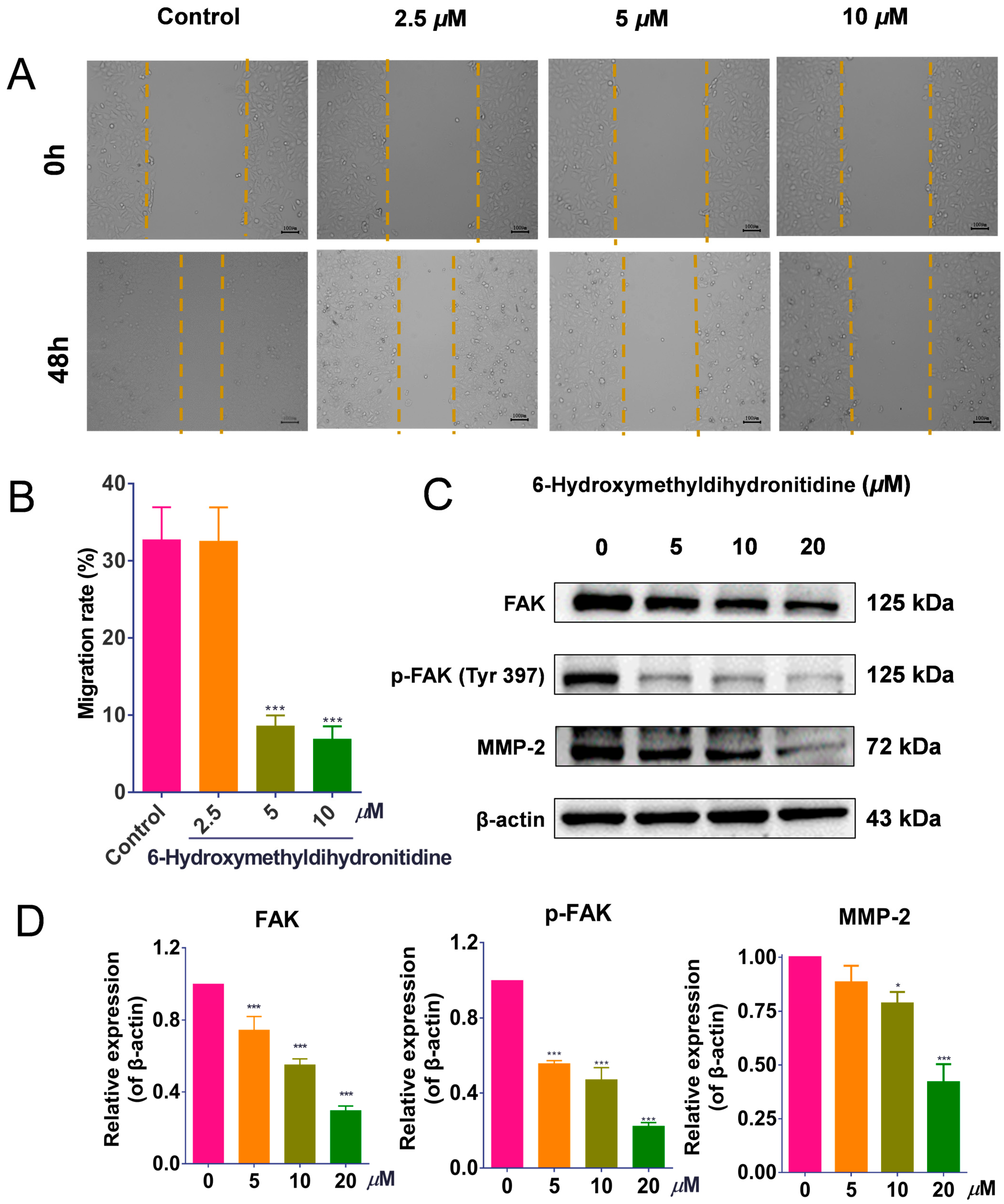
2.10. Wound-Scratch Assay
2.11. Western Blotting Analysis
2.12. Statistical Analysis
3. Results
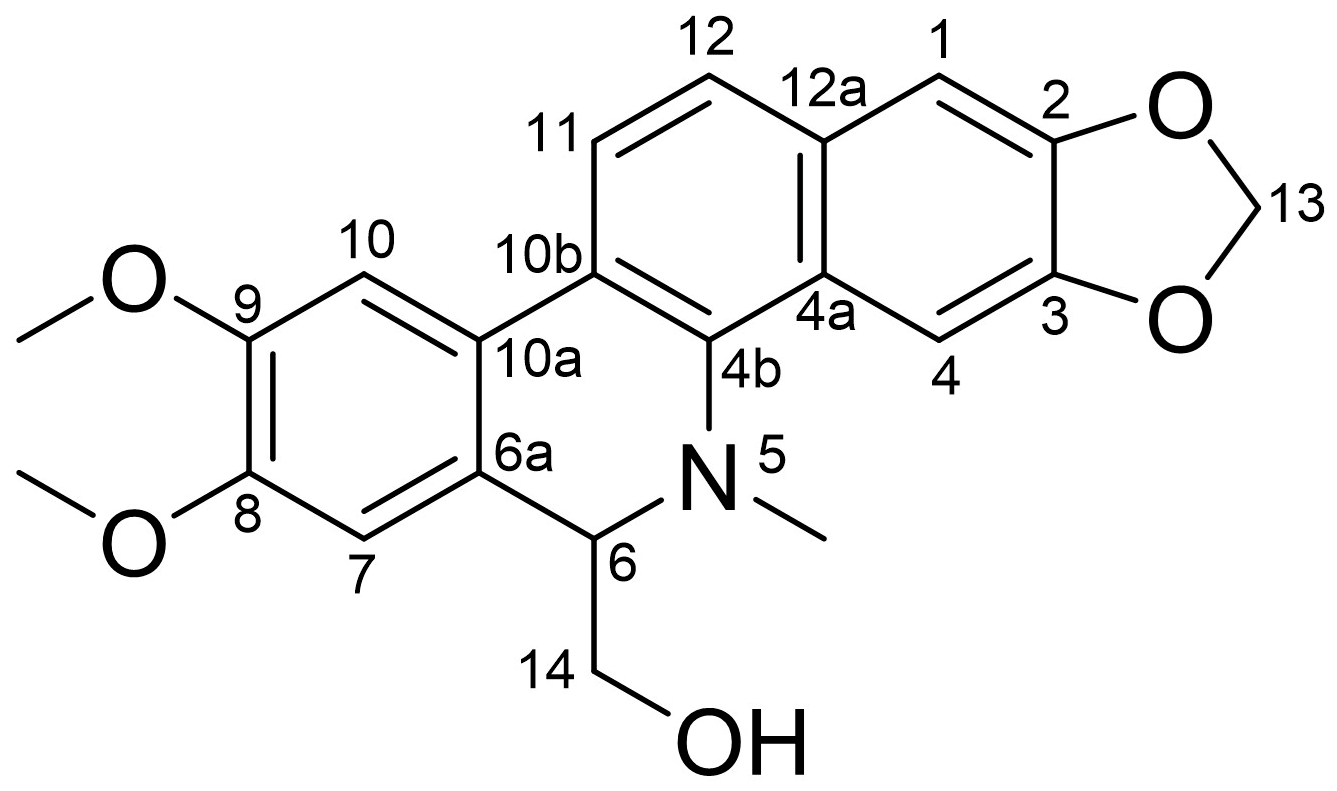
3.1. Identification of 6-HMDN
3.2. 6-HMDN Inhibited Tumor Cell Proliferation In Vitro
3.3. 6-HMDN Inhibited Tumor Proliferation and Metastasis In Vivo
3.4. 6-HMDN Induced Apoptosis in HepG2 Cells
3.5. 6-HMDN Increased Intracellular ROS Levels in HepG2 Cells
3.6. 6-HMDN Activated Ferroptosis
3.7. 6-HMDN Reduced HepG2 Cell Migration
4. Discussion
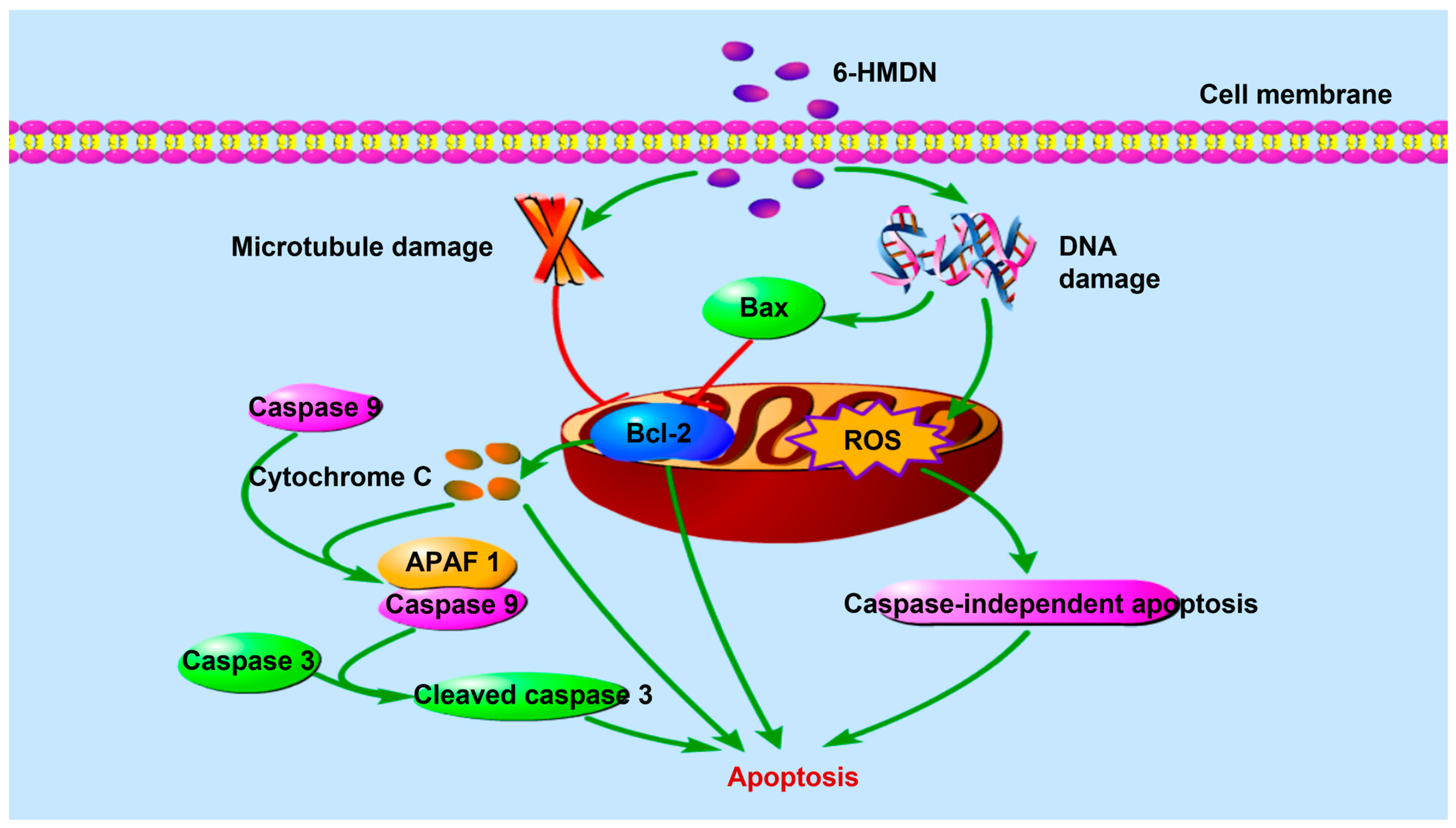
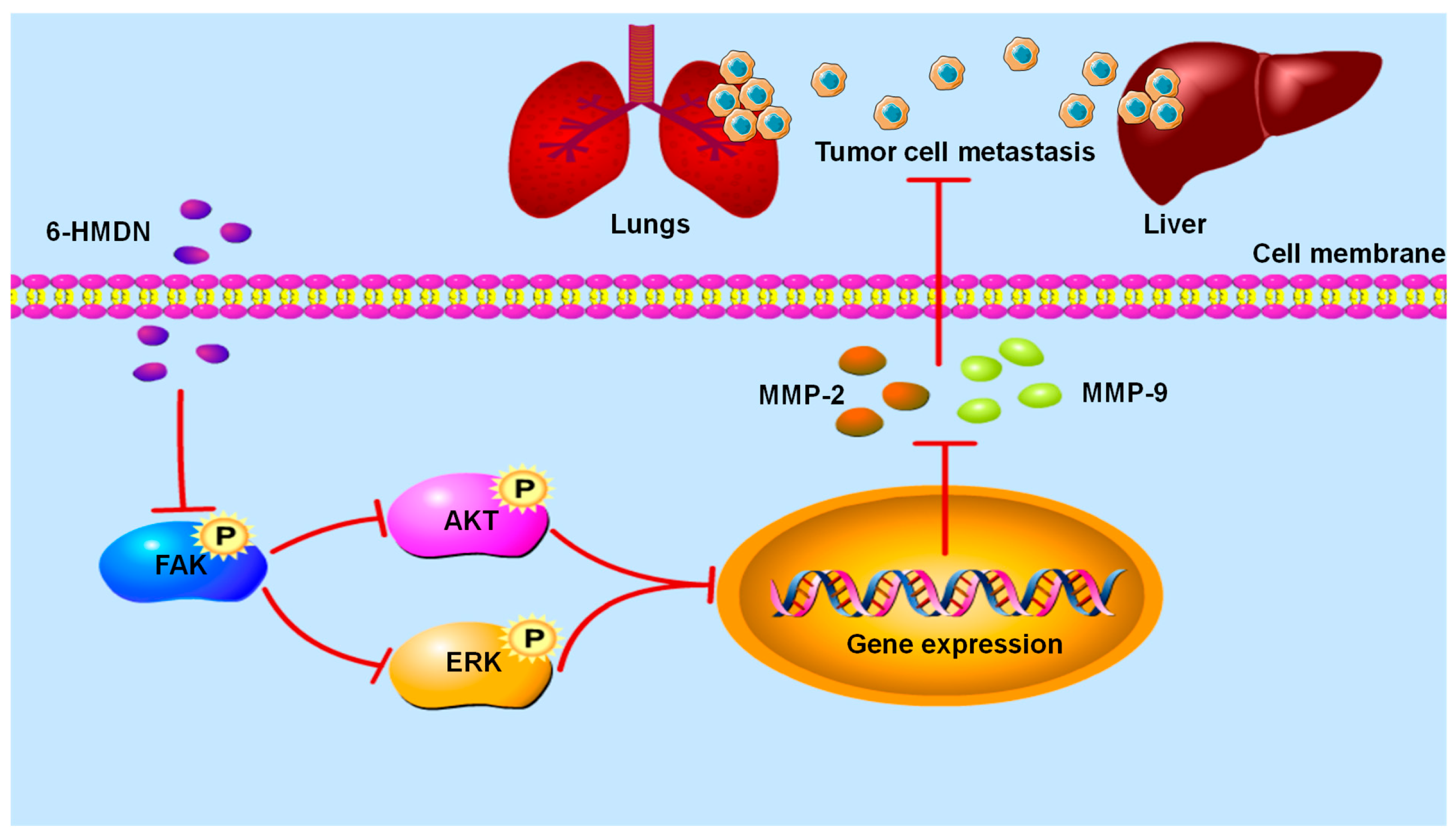
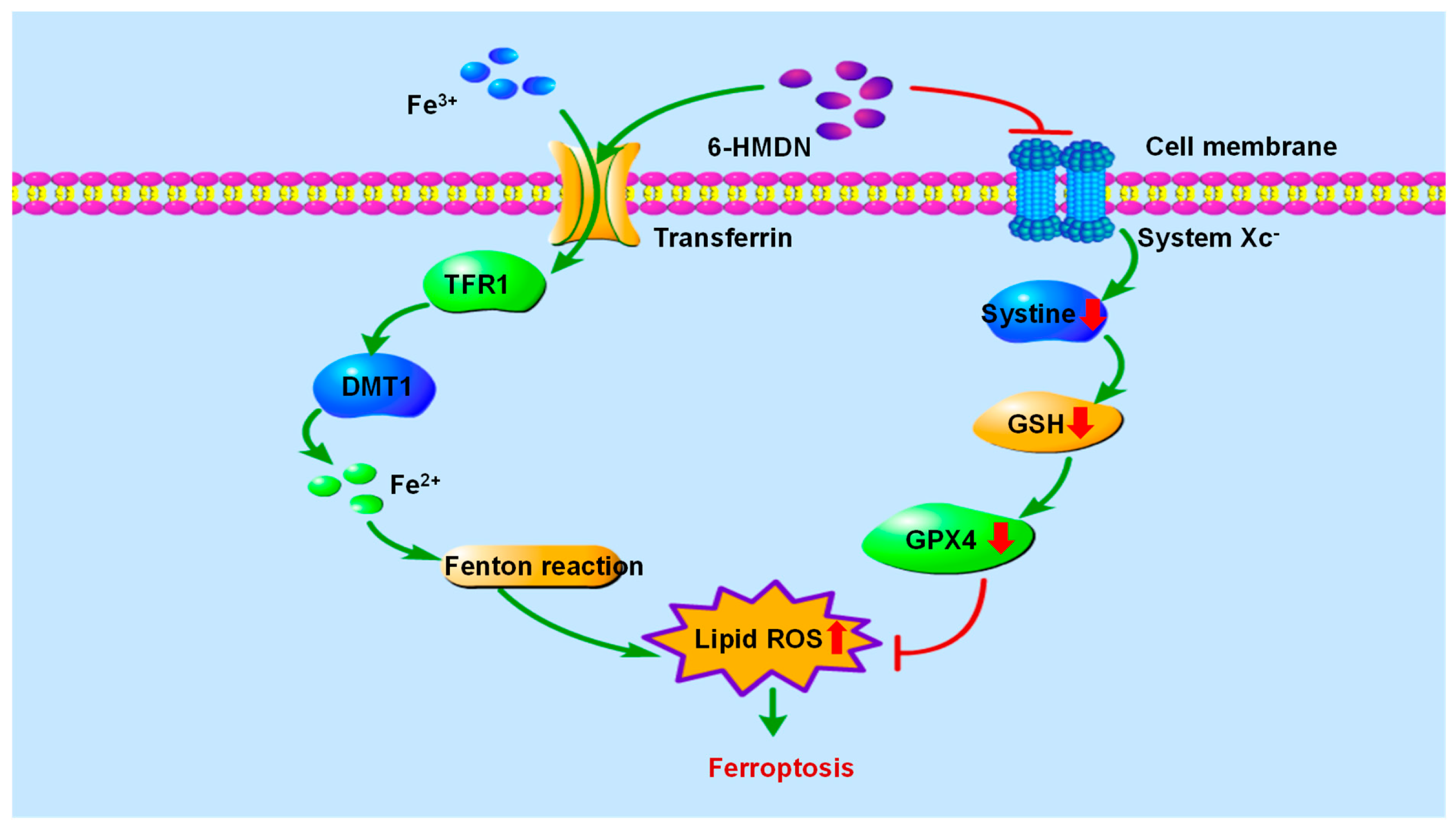
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luzzati, T.; Parenti, A.; Rughi, T. Economic Growth and Cancer Incidence. Ecol. Econ. 2018, 146, 381–396. [Google Scholar] [CrossRef]
- Saha, S.; Tandon, R.; Sanku, J.; Kumari, A.; Shukla, R.; Srivastava, N. siRNA-based Therapeutics in Hormone-driven Cancers: Advancements and benefits over conventional treatments. Int. J. Pharm. 2025, 674, 125463. [Google Scholar] [CrossRef] [PubMed]
- Vasan, N.; Baselga, J.; Hyman, D.M. A view on drug resistance in cancer. Nature 2019, 575, 299–309. [Google Scholar] [CrossRef]
- Ghosh, S.; Das, S.K.; Sinha, K.; Ghosh, B.; Sen, K.; Ghosh, N.; Sil, P.C. The Emerging Role of Natural Products in Cancer Treatment. Arch. Toxicol. 2024, 98, 2353–2391. [Google Scholar] [CrossRef]
- Hockenbery, D.M. bcl-2 in cancer, development and apoptosis. J. Cell Sci. Suppl. 1994, 18, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Skoupilová, H.; Michalová, E.; Hrstka, R. Ferroptosis as a New Type of Cell Death and its Role in Cancer Treatment. Klin Onkol. 2018, 31 (Suppl. 2), 21–26. [Google Scholar] [CrossRef]
- Olofinsan, K.; Abrahamse, H.; George, B.P. Therapeutic Role of Alkaloids and Alkaloid Derivatives in Cancer Management. Molecules 2023, 28, 5578. [Google Scholar] [CrossRef]
- Qin, R.; You, F.M.; Zhao, Q.; Xie, X.; Peng, C.; Zhan, G.; Han, B. Naturally derived indole alkaloids targeting regulated cell death (RCD) for cancer therapy: From molecular mechanisms to potential therapeutic targets. J. Hematol. Oncol. 2022, 15, 133. [Google Scholar] [CrossRef]
- Li, B.C.; Chen, J.; He, J.; Peng, J.; Wang, Y.X.; Liu, S.; Jiang, Y.P. Total alkaloids in Stephania tetrandra induce apoptosis by regulating BBC3 in human non-small cell lung cancer cells. Biomed. Pharmacother. 2023, 162, 114635. [Google Scholar] [CrossRef]
- Liao, W.L.; Zhang, R.; Chen, G.; Zhu, X.Y.; Wu, W.Y.; Chen, Z.Y.; Jiang, C.Y.; Lin, Z.C.; Ma, L.J.; Yu, H.J. Berberine synergises with ferroptosis inducer sensitizing NSCLC to ferroptosis in p53-dependent SLC7A11-GPX4 pathway. Biomed. Pharmacother. 2024, 176, 116832. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.H.; Chien, C.T.; Teng, K.Y.; Chou, S.T. Antioxidative and mutagenic properties of Zanthoxylum ailanthoides Sieb & zucc. Food Chem. 2006, 97, 418–425. [Google Scholar]
- Filippi-Chiela, E.C.; Vargas, J.E.; Bueno e Silva, M.M.; Thomé, M.P.; Lenz, G. Vincristine promotes differential levels of apoptosis, mitotic catastrophe, and senescence depending on the genetic background of glioblastoma cells. Toxicol. Vitr. 2022, 85, 105472. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.J.; Lee, K.H.; Tsai, I.L.; Chen, I.S. Two new sesquiterpenoids and anti-HIV principles from the root bark of Zanthoxylum ailanthoides. Bioorg. Med. Chem. 2005, 13, 5915–5920. [Google Scholar] [CrossRef]
- Chen, J.J.; Chung, C.Y.; Hwang, T.L.; Chen, J.F.; Chang, W.H.; Ho, Y.H.; Lin, C.W. New alkaloid, benzenoid, and anti-inflammatory constituents from Zanthoxylum ailanthoides. Planta Med. 2008, 74, PB51. [Google Scholar] [CrossRef]
- Teshima, S.; Yamashita-Higuchi, Y.; Sugimoto, S.; Matsunami, K.; Otsuka, H.; Shinzato, T. Zanthosides A–D: Four Aromatic Glucosides from the Leaves of Zanthoxylum ailanthoides. Chem. Pharm. Bull. 2020, 68, 814–817. [Google Scholar] [CrossRef]
- Sheen, W.S.; Tsai, I.L.; Teng, C.M.; Chen, I.S. Nor-neolignan and phenyl propanoid from Zanthoxylum ailanthoides. Phytochemistry 1994, 36, 213–215. [Google Scholar]
- Liu, W.; Zhang, H.; Wan, H.; Hou, J.; Lee, D.; Xu, J.; Guo, Y. Anti-inflammatory withanolides from the aerial parts of Physalis minima. Phytochemistry 2022, 202, 113301. [Google Scholar] [CrossRef]
- Liu, W.H.; Li, N.; Hou, J.T.; Cao, R.Y.; Jia, L.Y.; Guo, Y.Q.; Xu, J. Structure and antitumor activity of a polysaccharide from Rosa roxburghii. Int. J. Biol. Macromol. 2024, 273, 132807. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, R.; Zhong, K.; Guo, W.; Tong, A. An Anti-CD7 Antibody–Drug Conjugate Target Showing Potent Antitumor Activity for T-Lymphoblastic Leukemia (T-ALL). Biomolecules 2024, 14, 106. [Google Scholar] [CrossRef]
- Cocchi, V.; Jávega, B.; Gasperini, S.; O’Connor, J.-E.; Lenzi, M.; Hrelia, P. 6-(Methylsulfonyl) Hexyl Isothiocyanate: A Chemopreventive Agent Inducing Autophagy in Leukemia Cell Lines. Biomolecules 2022, 12, 1485. [Google Scholar] [CrossRef] [PubMed]
- Cao, R.Y.; Liu, Y.H.; Bao, J.H.; Rong, M.M.; Xu, J.; Liao, H.B.; Guo, Y.Q. Faberidilactone A, a Sesquiterpene Dimer, Inhibits Hepatocellular Carcinoma Progression Through Apoptosis, Ferroptosis, and Anti-Metastatic Mechanisms. Molecules 2025, 30, 1095. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, C.; Cai, Y.; Xu, A.; Han, C.; Tong, Y.; Cheng, S.; Liu, M. Revealing the Mechanism of Esculin in Treating Renal Cell Carcinoma Based on Network Pharmacology and Experimental Validation. Biomolecules 2024, 14, 1043. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Hua, S.X.; Deng, L.L.; Hao, X.J.; Mu, S.Z. Study on the Antitumor Active Alkaloid Components in Zanthoxylum nitidum Produced in Yunnan Province. Nat. Prod. Res. Dev. 2020, 32, 1370–1378. [Google Scholar]
- Yang, T.H.; Lee, C.I.; Huang, W.H.; Lee, A.R. Structural optimization and evaluation of novel 2-pyrrolidone-fused (2-oxoindolin-3-ylidene)methylpyrrole derivatives as potential VEGFR-2/PDGFRβ inhibitors. Chem. Cent. J. 2017, 11, 72. [Google Scholar] [CrossRef]
- Zhang, X.H.; Ma, H.J.; Gao, Y.; Liang, Y.B.; Du, Y.T.; Hao, S.L.; Ni, T. The Tumor Microenvironment: Signal Transduction. Biomolecules 2024, 14, 438. [Google Scholar] [CrossRef]
- Zhang, P.; Huang, C.R.; Wang, W.; Zhang, X.K.; Chen, J.J.; Wang, J.J.; Lin, C.; Jiang, J.W. Harmine Hydrochloride Triggers G2 Phase Arrest and Apoptosis in MGC-803 Cells and SMMC-7721 Cells by Upregulating p21, Activating Caspase-8/Bid, and Downregulating ERK/Bad Pathway. Phytother. Res. 2016, 30, 31–40. [Google Scholar] [CrossRef]
- Ola, M.S.; Nawaz, M.; Ahsan, H. Role of Bcl-2 family proteins and caspases in the regulation of apoptosis. Mol. Cell. Biochem. 2011, 351, 41–58. [Google Scholar] [CrossRef]
- Moradipour, A.; Dariushnejad, H.; Ahmadizadeh, C.; Lashgarian, H.E. Dietary flavonoid carvacrol triggers the apoptosis of human breast cancer MCF-7 cells via the p53/Bax/Bcl-2 axis. Med. Oncol. 2022, 40, 46. [Google Scholar] [CrossRef]
- Hou, D.; Liu, Z.J.; Xu, X.H.; Liu, Q.; Zhang, X.Y.; Kong, B.H.; Wei, J.J.; Gong, Y.Q.; Shao, C.S. Increased oxidative stress mediates the antitumor effect of PARP inhibition in ovarian cancer. Redox Biol. 2018, 17, 99–111. [Google Scholar] [CrossRef]
- Kojima, Y.; Tanaka, M.; Sasaki, M.; Ozeki, K.; Shimura, T.; Kubota, E.; Kataoka, H. Induction of ferroptosis by photodynamic therapy and enhancement of antitumor effect with ferroptosis inducers. J. Gastroenterol. 2024, 59, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shen, T.; Lian, J.; Deng, K.; Qu, C.; Li, E.; Li, G.; Ren, Y.; Wang, Z.; Jiang, Z.; et al. Resveratrol reduces ROS-induced ferroptosis by activating SIRT3 and compensating the GSH/GPX4 pathway. Mol. Med. 2023, 29, 137. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Xie, M.X.; Jia, Z.Y.; Tao, Z.W.; Zhao, P.; Ying, M.Y. FOXF1 inhibits invasion and metastasis of lung adenocarcinoma cells and enhances anti-tumor immunity via MFAP4/FAK signal axis. Sci. Rep. 2024, 14, 21451. [Google Scholar] [CrossRef]
- Sitarek, P.; Toma, M.; Ntungwe, E.; Kowalczyk, T.; Skała, E.; Wieczfinska, J.; Śliwiński, T.; Rijo, P. Insight the Biological Activities of Selected Abietane Diterpenes Isolated from Plectranthus spp. Biomolecules 2020, 10, 194. [Google Scholar] [CrossRef]
- Dhyani, P.; Quispe, C.; Sharma, E.; Bahukhandi, A.; Sati, P.; Attri, D.C.; Szopa, A.; Sharifi-Rad, J.; Docea, A.O.; Mardare, I.; et al. Anticancer potential of alkaloids: A key emphasis to colchicine, vinblastine, vincristine, vindesine, vinorelbine and vincamine. Cancer Cell Int. 2022, 22, 206. [Google Scholar] [CrossRef] [PubMed]
- Walton, M.I.; Whysong, D.; O’Connor, P.M.; Hockenbery, D.; Korsmeyer, S.J.; Kohn, K.W. Constitutive expression of human Bcl-2 modulates nitrogen mustard and camptothecin induced apoptosis. Cancer Res. 1993, 53, 1853–1861. [Google Scholar]
- Ho, Y.T.; Yang, J.S.; Li, T.C.; Lin, J.J.; Lin, J.G.; Lai, K.C.; Ma, C.Y.; Wood, W.G.; Chung, J.G. Berberine suppresses in vitro migration and invasion of human SCC-4 tongue squamous cancer cells through the inhibitions of FAK, IKK, NF-κB, u-PA and MMP-2 and -9. Cancer Lett. 2009, 279, 155–162. [Google Scholar] [CrossRef]
- Yang, L.; Li, X.; Huang, W.; Rao, X.; Lai, Y. Pharmacological properties of indirubin and its derivatives. Biomed. Pharmacother. 2022, 151, 113112. [Google Scholar] [CrossRef]
- Jing, Y.M.; Liang, W.Q.; Liu, J.; Zhang, L.; Wei, J.G.; Zhu, Y.F.; Yang, J.H.; Ji, K.W.; Zhang, Y.; Huang, Z.L. Stress-induced phosphoprotein 1 promotes pancreatic cancer progression through activation of the FAK/AKT/MMP signaling axis. Pathol. Res. Pract. 2019, 215, 152564. [Google Scholar] [CrossRef]
- Ru, Q.; Li, Y.S.; Chen, L.; Wu, Y.X.; Min, J.X.; Wang, F.D. Iron homeostasis and ferroptosis in human diseases: Mechanisms and therapeutic prospects. Signal Transduct. Targeted Ther. 2024, 9, 271. [Google Scholar]
- Fan, B.; Guo, Q.; Wang, S. The application of alkaloids in ferroptosis: A review. Biomed. Pharmacother. 2024, 178, 117232. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Su, Q.; Pan, H.; You, Y.L.; Ruan, Z.Y.; Wu, Y.H.; Tang, Z.X.; Hu, L.M. Arsenic-Induced Ferroptosis in Chicken Hepatocytes via the Mitochondrial ROS Pathway. Biol. Trace Elem. Res. 2024, 202, 4180–4190. [Google Scholar] [CrossRef] [PubMed]
- Affranchi, F.; Di Liberto, D.; Lauricella, M.; D’Anneo, A.; Calvaruso, G.; Pratelli, G.; Carlisi, D.; De Blasio, A.; Tesoriere, L.; Giuliano, M.; et al. The Antitumor Potential of Sicilian Grape Pomace Extract: A Balance between ROS-Mediated Autophagy and Apoptosis. Biomolecules 2024, 14, 1111. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δC | δH |
|---|---|---|
| 1 | 104.7 | 7.13, s |
| 2 | 147.6 | |
| 3 | 148.5 | |
| 4 | 99.8 | 7.65, s |
| 4a | 127.0 | |
| 4b | 137.9 | |
| 6 | 65.4 | 4.15, dd (10.7, 3.8) |
| 6a | 124.0 | |
| 7 | 110.7 | 6.80, s |
| 8 | 149.1 | |
| 9 | 149.1 | |
| 10 | 106.4 | 7.34, s |
| 10a | 124.3 | |
| 10b | 123.5 | |
| 11 | 119.6 | 7.69, d (8.5) |
| 12 | 124.3 | 7.52, d (8.5) |
| 12a | 131.0 | |
| 13 | 101.2 | 6.06, s |
| 14 | 62.9 | 3.15, t (10.7) 3.47, dd (10.7, 3.8) |
| 8-OMe | 56.1 | 4.00, s |
| 9-OMe | 56.1 | 3.96, s |
| N-Me | 42.8 | 2.74, s |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, H.; Hou, J.; Wang, J.; Xu, J.; Guo, Y. A Natural Alkaloid, 6-Hydroxymethyldihydronitidine, Suppresses Tumor Progression by Co-Regulating Apoptosis, Ferroptosis, and FAK Pathways. Biomolecules 2025, 15, 814. https://doi.org/10.3390/biom15060814
Jiang H, Hou J, Wang J, Xu J, Guo Y. A Natural Alkaloid, 6-Hydroxymethyldihydronitidine, Suppresses Tumor Progression by Co-Regulating Apoptosis, Ferroptosis, and FAK Pathways. Biomolecules. 2025; 15(6):814. https://doi.org/10.3390/biom15060814
Chicago/Turabian StyleJiang, Haojing, Jiantong Hou, Jianliang Wang, Jing Xu, and Yuanqiang Guo. 2025. "A Natural Alkaloid, 6-Hydroxymethyldihydronitidine, Suppresses Tumor Progression by Co-Regulating Apoptosis, Ferroptosis, and FAK Pathways" Biomolecules 15, no. 6: 814. https://doi.org/10.3390/biom15060814
APA StyleJiang, H., Hou, J., Wang, J., Xu, J., & Guo, Y. (2025). A Natural Alkaloid, 6-Hydroxymethyldihydronitidine, Suppresses Tumor Progression by Co-Regulating Apoptosis, Ferroptosis, and FAK Pathways. Biomolecules, 15(6), 814. https://doi.org/10.3390/biom15060814