
Methodological Pitfalls of Investigating Lipid Rafts in the Brain: What Are We Still Missing?




Abstract
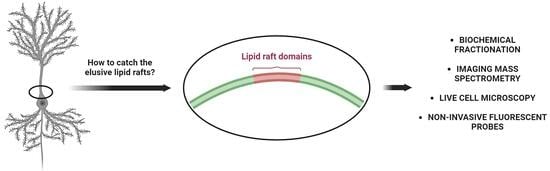
1. Introduction
2. Structure and Function of Lipid Rafts

{kind=link}
{kind=link}
{kind=link}
| Raft Component | Functional Characteristic of a Raft Component |
|---|---|
| Lipids | |
| Phospholipids | A form fluid phospholipid bilayer made of saturated fatty-acid side chains of the phospholipids. This allows close packing with the saturated acyl chains of sphingolipids. Phospholipids provide flexibility and fluidity, allowing lateral movement of proteins and other molecules within the raft [21] and interact with cholesterol [22]. |
| Phosphatidic acids (PA) | It contributes to the overall structural organization of lipid rafts affecting the fluidity and packing of lipid molecules within the raft [23]. In addition to that, PA is involved in regulation of membrane curvature [24], protein–lipid interactions within lipid rafts [23], and signaling modulation [25]. |
| Phosphatidylcholines (PC) | Present in small amounts in lipid rafts [9]. Each of these phospholipids (PC, PE, PS, PI) contributes differently to the overall structure and function of lipid rafts, due to variability of both polar groups and saturation and length of fatty acyl residues in the hydrophobic backbone [26,27,28,29,30]. |
| Phosphatidylethanolamines (PE) | |
| Phosphatidylserines (PS) | |
| Phosphatidylinositols (PI) | |
| Sphingolipids (mostly sphingomyelin) | Mostly present in the outer leaflet of plasma membrane [31]. Sphingolipids are integral to the structure and function of lipid rafts, contributing to their stability [32], organization, and involvement in signaling pathways [31]. The variations in the hydrophobic part of the molecule crucially contribute to lipid raft organization. |
| Cholesterol and oxysterols | Cholesterol plays a fundamental and multifaceted role in the structure and function of lipid rafts affecting structural integrity [21], modulating fluidity, organization and segregation of lipids and proteins within lipid rafts, modulation of cellular signaling pathways, endocytosis, and intracellular trafficking while cholesterol rich lipid rafts are often targeted by pathogens for cellular entry [21,22,32,33,34,35]. Oxysterols, derivatives of cholesterol that can either promote or inhibit the formation of lipid rafts, play a crucial role in signaling processes [36,37]. |
| Glycosphingolipids | Gangliosides are sialylated glycosphingolipids, highly diverse and abundant in the mammalian brain. Gangliosides play important roles in various cellular processes, and their presence in lipid rafts has implications for the structure and function of these membrane microdomains [38,39,40]. |
| Proteins | |
| Raftophilic proteins | |
| GPI-anchored proteins | Proteins anchored to the cell membrane via a GPI moiety, such as CD55, CD59 and Thy 1 are raftophilic and are involved in cell adhesion, signal transduction, immune regulation immune regulation, or neuronal connectivity [41]. The prion protein (PrPC) is also an important GPI-anchored protein, highly expressed in neurons that contain a sialic acid in their GPI-anchor structures [41]. Recent studies indicate PrPCas a key factor in cell fate regulation [42] and neuronal differentiation [43]. |
| Receptor Tyrosine Kinases (RTKs) | Several RTKs, including the epidermal growth factor receptor (EGFR) and insulin receptor, preferentially associate with lipid rafts, influencing downstream signaling pathways [44]. |
| G-Protein-Coupled Receptors (GPCRs) | Certain GPCRs, such as the serotonin receptor, are raftophilic. Their localization in lipid rafts impacts receptor signaling and cellular responses [45]. |
| Src-Family Kinases | Src-family kinases, including Src and Lyn, are raftophilic proteins involved in signaling cascades and cellular processes such as proliferation and migration [46]. |
| Cholesterol-Binding Proteins | Cholesterol-binding proteins contribute to the stability and functionality of lipid rafts [10]. |
| Intracellular Trafficking Proteins | Proteins involved in vesicle trafficking and membrane transport are associated with lipid rafts. Their presence contributes to the regulation of endocytosis, exocytosis, and intracellular membrane dynamics [47]. |
| Immunoreceptors | Immunoreceptors, including those on T and B cells, are known to be localized in lipid rafts. This localization is critical for efficient immune cell activation and response [48]. |
| Autophagy-Related Proteins | The association of autophagy-related proteins (LC3, Beclin-1, ATG9, ULK1, WIPI) with lipid rafts depends on the cellular context, experimental conditions, and the specific phase of autophagy [49,50]. Additionally, the precise mechanisms through which lipid rafts influence autophagy and the functional significance of these associations are still areas of ongoing research. |
| Apoptosis-Related Proteins | Apoptosis-related proteins like Fas/CD95, caspases, Bcl-2 family proteins, DAPK, TNFR1, PrP, and FLIP were identified as associating with lipid rafts or being influenced by the organization of these membrane microdomains [42,51,52]. |
3. Lipid Rafts in the Brain
4. Methodology behind Lipid Rafts: Biochemical Methods
4.1. Cholesterol and Oxysterols
4.2. Raft Proteins and Bulk Membrane Protein Markers
4.3. Phospholipids
4.4. Sphingolipids
5. Visualizing Lipid Rafts
5.1. Antibodies, Toxins, and Fluorescent Probes
5.2. Direct Visualization
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Danielli, J.F.; Davson, H. A Contribution to the Theory of Permeability of Thin Films. J. Cell. Comp. Physiol. 1935, 5, 495–508. [Google Scholar] [CrossRef]
- Singer, S.J.; Nicolson, G.L. The Fluid Mosaic Model of the Structure of Cell Membranes. Science 1972, 175, 720–731. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Van Meer, G. Lipid Sorting in Epithelial Cells. Biochemistry 1988, 27, 6197–6202. [Google Scholar] [CrossRef] [PubMed]
- Goodsaid-Zalduondo, F.; Rintoul, D.A.; Carlson, J.C.; Hansel, W. Luteolysis-Induced Changes in Phase Composition and Fluidity of Bovine Luteal Cell Membranes. Proc. Natl. Acad. Sci. USA 1982, 79, 4332–4336. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Ikonen, E. Functional Rafts in Cell Membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Pike, L.J. Rafts Defined: A Report on the Keystone Symposium on Lipid Rafts and Cell Function. J. Lipid Res. 2006, 47, 1597–1598. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.E. Chapter 53—Lipid-Mediated Localization of Signaling Proteins. In Handbook of Cell Signaling, 2nd ed.; Bradshaw, R.A., Dennis, E.A., Eds.; Academic Press: San Diego, CA, USA, 2010; pp. 365–371. ISBN 978-0-12-374145-5. [Google Scholar]
- Head, B.P.; Patel, H.H.; Insel, P.A. Interaction of Membrane/Lipid Rafts with the Cytoskeleton: Impact on Signaling and Function. Biochim. Biophys. Acta 2014, 1838, 532–545. [Google Scholar] [CrossRef] [PubMed]
- Pike, L.J. Lipid Rafts: Bringing Order to Chaos. J. Lipid Res. 2003, 44, 655–667. [Google Scholar] [CrossRef]
- Simons, K.; Sampaio, J.L. Membrane Organization and Lipid Rafts. Cold Spring Harb. Perspect. Biol. 2011, 3, a004697. [Google Scholar] [CrossRef]
- Sapoń, K.; Mańka, R.; Janas, T.; Janas, T. The Role of Lipid Rafts in Vesicle Formation. J. Cell Sci. 2023, 136, jcs260887. [Google Scholar] [CrossRef]
- Chamberlain, L.H.; Gould, G.W. The Vesicle-and Target-SNARE Proteins That Mediate Glut4 Vesicle Fusion Are Localized in Detergent-Insoluble Lipid Rafts Present on Distinct Intracellular Membranes. J. Biol. Chem. 2002, 277, 49750–49754. [Google Scholar] [CrossRef]
- Fiedler, K.; Kobayashi, T.; Kurzchalia, T.V.; Simons, K. Glycosphingolipid-Enriched, Detergent-Insoluble Complexes in Protein Sorting in Epithelial Cells. Biochemistry 1993, 32, 6365–6373. [Google Scholar] [CrossRef]
- Ouweneel, A.B.; Thomas, M.J.; Sorci-Thomas, M.G. The Ins and Outs of Lipid Rafts: Functions in Intracellular Cholesterol Homeostasis, Microparticles, and Cell Membranes: Thematic Review Series: Biology of Lipid Rafts. J. Lipid Res. 2020, 61, 676–686. [Google Scholar] [CrossRef]
- Garofalo, T.; Giammarioli, A.M.; Misasi, R.; Tinari, A.; Manganelli, V.; Gambardella, L.; Pavan, A.; Malorni, W.; Sorice, M. Lipid Microdomains Contribute to Apoptosis-Associated Modifications of Mitochondria in T Cells. Cell Death Differ. 2005, 12, 1378–1389. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Su, T.-P. σ-1 Receptors (Σ1 Binding Sites) Form Raft-Like Microdomains and Target Lipid Droplets on the Endoplasmic Reticulum: Roles in Endoplasmic Reticulum Lipid Compartmentalization and Export. J. Pharmacol. Exp. Ther. 2003, 306, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, I.; Sano, R.; d’Azzo, A. Mitochondria-Associated ER Membranes (MAMs) and Lysosomal Storage Diseases. Cell Death Dis. 2018, 9, 328. [Google Scholar] [CrossRef] [PubMed]
- Resende, R.; Fernandes, T.; Pereira, A.C.; De Pascale, J.; Marques, A.P.; Oliveira, P.; Morais, S.; Santos, V.; Madeira, N.; Pereira, C.F.; et al. Mitochondria, Endoplasmic Reticulum and Innate Immune Dysfunction in Mood Disorders: Do Mitochondria-Associated Membranes (MAMs) Play a Role? Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165752. [Google Scholar] [CrossRef]
- Manganelli, V.; Matarrese, P.; Antonioli, M.; Gambardella, L.; Vescovo, T.; Gretzmeier, C.; Longo, A.; Capozzi, A.; Recalchi, S.; Riitano, G.; et al. Raft-like Lipid Microdomains Drive Autophagy Initiation via AMBRA1-ERLIN1 Molecular Association within MAMs. Autophagy 2021, 17, 2528–2548. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R.R.; Montesinos, J.; Larrea, D.; Area-Gomez, E.; Pera, M. The Silence of the Fats: A MAM’s Story about Alzheimer. Neurobiol. Dis. 2020, 145, 105062. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Ehehalt, R. Cholesterol, Lipid Rafts, and Disease. J. Clin. Investig. 2002, 110, 597–603. [Google Scholar] [CrossRef]
- Crane, J.M.; Tamm, L.K. Role of Cholesterol in the Formation and Nature of Lipid Rafts in Planar and Spherical Model Membranes. Biophys. J. 2004, 86, 2965–2979. [Google Scholar] [CrossRef]
- Lee, A.G. Lipid–Protein Interactions in Biological Membranes: A Structural Perspective. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2003, 1612, 1–40. [Google Scholar] [CrossRef]
- Zhukovsky, M.A.; Filograna, A.; Luini, A.; Corda, D.; Valente, C. Phosphatidic Acid in Membrane Rearrangements. FEBS Lett. 2019, 593, 2428–2451. [Google Scholar] [CrossRef]
- Srivatsav, A.T.; Mishra, M.; Kapoor, S. Small-Molecule Modulation of Lipid-Dependent Cellular Processes against Cancer: Fats on the Gunpoint. BioMed Res. Int. 2018, 2018, e6437371. [Google Scholar] [CrossRef]
- Strzyz, P. Tying Lipid Rafts to Oncogenic Signalling. Nat. Rev. Mol. Cell Biol. 2019, 20, 513. [Google Scholar] [CrossRef]
- Nguyen, T.T.M.; Chintamsetti, V.R.; Chennuru, S. The Stability of Lipid Rafts-Like Micro-Domains Is Dependent on the Available Amount of Cholesterol. J. Biophys. Chem. 2016, 7, 74. [Google Scholar] [CrossRef]
- Grzybek, M.; Kubiak, J.; Łach, A.; Przybyło, M.; Sikorski, A.F. A Raft-Associated Species of Phosphatidylethanolamine Interacts with Cholesterol Comparably to Sphingomyelin. A Langmuir-Blodgett Monolayer Study. PLoS ONE 2009, 4, e5053. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Malcor, J.-D.M.; Harper, M.T. Lipid Rafts Are Essential for Release of Phosphatidylserine-Exposing Extracellular Vesicles from Platelets. Sci. Rep. 2018, 8, 9987. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Huang, F.; Xia, T.; Shi, Y.; Yue, T. Phosphatidylinositol 4,5-Bisphosphate Sensing Lipid Raft via Inter-Leaflet Coupling Regulated by Acyl Chain Length of Sphingomyelin. Langmuir 2023, 39, 5995–6005. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, D.; Maier, O.; van der Wouden, J.M.; Slimane, T.A.; van IJzendoorn, S.C.D. Membrane Dynamics and Cell Polarity: The Role of Sphingolipids. J. Lipid Res. 2003, 44, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Codini, M.; Garcia-Gil, M.; Albi, E. Cholesterol and Sphingolipid Enriched Lipid Rafts as Therapeutic Targets in Cancer. Int. J. Mol. Sci. 2021, 22, 726. [Google Scholar] [CrossRef]
- Vona, R.; Iessi, E.; Matarrese, P. Role of Cholesterol and Lipid Rafts in Cancer Signaling: A Promising Therapeutic Opportunity? Front. Cell Dev. Biol. 2021, 9, 622908. [Google Scholar] [CrossRef]
- Silvius, J.R. Role of Cholesterol in Lipid Raft Formation: Lessons from Lipid Model Systems. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2003, 1610, 174–183. [Google Scholar] [CrossRef]
- Korade, Z.; Kenworthy, A.K. Lipid Rafts, Cholesterol, and the Brain. Neuropharmacology 2008, 55, 1265–1273. [Google Scholar] [CrossRef]
- Filomenko, R.; Fourgeux, C.; Bretillon, L.; Gambert-Nicot, S. Oxysterols: Influence on Plasma Membrane Rafts Microdomains and Development of Ocular Diseases. Steroids 2015, 99, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Wnętrzak, A.; Chachaj–Brekiesz, A.; Stępniak, A.; Kobierski, J.; Dynarowicz–Latka, P. Different Effects of Oxysterols on a Model Lipid Raft—Langmuir Monolayer Study Complemented with Theoretical Calculations. Chem. Phys. Lipids 2022, 244, 105182. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Murakami, Y.; Yasui, T.; Wakana, S.; Kikutani, H.; Kinoshita, T.; Maeda, Y. Significance of Glycosylphosphatidylinositol-Anchored Protein Enrichment in Lipid Rafts for the Control of Autoimmunity. J. Biol. Chem. 2013, 288, 25490–25499. [Google Scholar] [CrossRef] [PubMed]
- Sonnino, S.; Mauri, L.; Chigorno, V.; Prinetti, A. Gangliosides as Components of Lipid Membrane Domains. Glycobiology 2007, 17, 1R–13R. [Google Scholar] [CrossRef]
- Fujinaga, Y.; Wolf, A.A.; Rodighiero, C.; Wheeler, H.; Tsai, B.; Allen, L.; Jobling, M.G.; Rapoport, T.; Holmes, R.K.; Lencer, W.I. Gangliosides That Associate with Lipid Rafts Mediate Transport of Cholera and Related Toxins from the Plasma Membrane to Endoplasmic Reticulm. Mol. Biol. Cell 2003, 14, 4783–4793. [Google Scholar] [CrossRef]
- Puig, B.; Altmeppen, H.; Glatzel, M. The GPI-Anchoring of PrP. Prion 2014, 8, 11–18. [Google Scholar] [CrossRef]
- Sorice, M.; Mattei, V.; Tasciotti, V.; Manganelli, V.; Garofalo, T.; Misasi, R. Trafficking of PrPc to Mitochondrial Raft-like Microdomains during Cell Apoptosis. Prion 2012, 6, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Martellucci, S.; Santacroce, C.; Santilli, F.; Manganelli, V.; Sorice, M.; Mattei, V. Prion Protein in Stem Cells: A Lipid Raft Component Involved in the Cellular Differentiation Process. Int. J. Mol. Sci. 2020, 21, 4168. [Google Scholar] [CrossRef]
- Pike, L.J. Growth Factor Receptors, Lipid Rafts and Caveolae: An Evolving Story. Biochim. Et Biophys. Acta (BBA)—Mol. Cell Res. 2005, 1746, 260–273. [Google Scholar] [CrossRef] [PubMed]
- Villar, V.A.M.; Cuevas, S.; Zheng, X.; Jose, P.A. Localization and Signaling of GPCRs in Lipid Rafts. Methods Cell Biol. 2016, 132, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Arcaro, A.; Aubert, M.; Espinosa del Hierro, M.E.; Khanzada, U.K.; Angelidou, S.; Tetley, T.D.; Bittermann, A.G.; Frame, M.C.; Seckl, M.J. Critical Role for Lipid Raft-Associated Src Kinases in Activation of PI3K-Akt Signalling. Cell. Signal. 2007, 19, 1081–1092. [Google Scholar] [CrossRef]
- Helms, J.B.; Zurzolo, C. Lipids as Targeting Signals: Lipid Rafts and Intracellular Trafficking—PubMed. Traffic 2004, 5, 247–254. Available online: https://pubmed.ncbi.nlm.nih.gov/15030566/ (accessed on 30 December 2023). [CrossRef]
- Varshney, P.; Yadav, V.; Saini, N. Lipid Rafts in Immune Signalling: Current Progress and Future Perspective. Immunology 2016, 149, 13–24. [Google Scholar] [CrossRef]
- Li, X.; He, S.; Ma, B. Autophagy and Autophagy-Related Proteins in Cancer. Mol. Cancer 2020, 19, 12. [Google Scholar] [CrossRef]
- Mercer, T.J.; Gubas, A.; Tooze, S.A. A Molecular Perspective of Mammalian Autophagosome Biogenesis. J. Biol. Chem. 2018, 293, 5386–5395. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Mollinedo, F.; Gajate, C. Mitochondrial Targeting Involving Cholesterol-Rich Lipid Rafts in the Mechanism of Action of the Antitumor Ether Lipid and Alkylphospholipid Analog Edelfosine. Pharmaceutics 2021, 13, 763. [Google Scholar] [CrossRef]
- Grassi, S.; Giussani, P.; Mauri, L.; Prioni, S.; Sonnino, S.; Prinetti, A. Lipid Rafts and Neurodegeneration: Structural and Functional Roles in Physiologic Aging and Neurodegenerative Diseases. J. Lipid Res. 2020, 61, 636–654. Available online: https://www.jlr.org/article/S0022-2275(20)43614-4/fulltext (accessed on 30 December 2023). [CrossRef]
- Díaz, M.; Marin, R.; Díaz, M.; Marin, R. Lipid Rafts and Development of Alzheimer’s Disease. In Cerebral and Cerebellar Cortex—Interaction and Dynamics in Health and Disease; IntechOpen: London, UK, 2021; ISBN 978-1-83968-094-6. [Google Scholar]
- Grassmé, H.; Riethmüller, J.; Gulbins, E. Biological Aspects of Ceramide-Enriched Membrane Domains. Prog. Lipid Res. 2007, 46, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Ilic, K.; Lin, X.; Malci, A.; Stojanović, M.; Puljko, B.; Rožman, M.; Vukelić, Ž.; Heffer, M.; Montag, D.; Schnaar, R.L.; et al. Plasma Membrane Calcium ATPase-Neuroplastin Complexes Are Selectively Stabilized in GM1-Containing Lipid Rafts. Int. J. Mol. Sci. 2021, 22, 13590. [Google Scholar] [CrossRef]
- Chamberlain, L.H. Detergents as Tools for the Purification and Classification of Lipid Rafts. FEBS Lett. 2004, 559, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Sezgin, E.; Levental, I.; Mayor, S.; Eggeling, C. The Mystery of Membrane Organization: Composition, Regulation and Roles of Lipid Rafts. Nat. Rev. Mol. Cell Biol. 2017, 18, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Grassi, S.; Giussani, P.; Mauri, L.; Prioni, S.; Prinetti, A. Isolation and Analysis of Lipid Rafts from Neural Cells and Tissues | Springer Nature Experiments. Lipid Rafts Methods Protoc. 2021, pp. 1–25. Available online: https://experiments.springernature.com/articles/10.1007/978-1-0716-0814-2_1 (accessed on 30 December 2023).
- Wang, X.-Q.; Paller, A.S. Lipid Rafts: Membrane Triage Centers. J. Investig. Dermatol. 2006, 126, 951–953. [Google Scholar] [CrossRef] [PubMed]
- Aureli, M.; Grassi, S.; Sonnino, S.; Prinetti, A. Isolation and Analysis of Detergent-Resistant Membrane Fractions. In Lipid Signaling Protocols; Waugh, M.G., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2016; pp. 107–131. ISBN 978-1-4939-3170-5. [Google Scholar]
- Komura, N.; Suzuki, K.G.N.; Ando, H.; Konishi, M.; Koikeda, M.; Imamura, A.; Chadda, R.; Fujiwara, T.K.; Tsuboi, H.; Sheng, R.; et al. Raft-Based Interactions of Gangliosides with a GPI-Anchored Receptor. Nat. Chem. Biol. 2016, 12, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Seong, J. Lipid Rafts Signaling Detected by FRET-Based Molecular Biosensors. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 629–636. [Google Scholar] [CrossRef]
- Mlinac-Jerkovic, K.; Ilic, K.; Zjalić, M.; Mandić, D.; Debeljak, Ž.; Balog, M.; Damjanović, V.; Maček Hrvat, N.; Habek, N.; Kalanj-Bognar, S.; et al. Who’s in, Who’s out? Re-Evaluation of Lipid Raft Residents. J. Neurochem. 2021, 158, 657–672. [Google Scholar] [CrossRef]
- Schuck, S.; Honsho, M.; Ekroos, K.; Shevchenko, A.; Simons, K. Resistance of Cell Membranes to Different Detergents. Proc. Natl. Acad. Sci. USA 2003, 100, 5795–5800. Available online: https://www.pnas.org/doi/10.1073/pnas.0631579100 (accessed on 30 December 2023).
- Persaud-Sawin, D.-A.; Lightcap, S.; Harry, G.J. Isolation of Rafts from Mouse Brain Tissue by a Detergent-Free Method. J. Lipid Res. 2009, 50, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Nothdurfter, C.; Tanasic, S.; Di Benedetto, B.; Uhr, M.; Wagner, E.-M.; Gilling, K.E.; Parsons, C.G.; Rein, T.; Holsboer, F.; Rupprecht, R.; et al. Lipid Raft Integrity Affects GABAA Receptor, but Not NMDA Receptor Modulation by Psychopharmacological Compounds. Int. J. Neuropsychopharmacol. 2013, 16, 1361–1371. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.A.; Rose, J.K. Sorting of GPI-Anchored Proteins to Glycolipid-Enriched Membrane Subdomains during Transport to the Apical Cell Surface. Cell 1992, 68, 533–544. [Google Scholar] [CrossRef]
- Scandroglio, F.; Venkata, J.K.; Loberto, N.; Prioni, S.; Schuchman, E.H.; Chigorno, V.; Prinetti, A.; Sonnino, S. Lipid Content of Brain, Brain Membrane Lipid Domains, and Neurons from Acid Sphingomyelinase Deficient Mice. J. Neurochem. 2008, 107, 329–338. [Google Scholar] [CrossRef]
- Drevot, P.; Langlet, C.; Guo, X.-J.; Bernard, A.-M.; Colard, O.; Chauvin, J.-P.; Lasserre, R.; He, H.-T. TCR Signal Initiation Machinery Is Pre-Assembled and Activated in a Subset of Membrane Rafts. EMBO J. 2002, 21, 1899–1908. [Google Scholar] [CrossRef]
- Kosicek, M.; Malnar, M.; Goate, A.; Hecimovic, S. Cholesterol Accumulation in Niemann Pick Type C (NPC) Model Cells Causes a Shift in APP Localization to Lipid Rafts. Biochem. Biophys. Res. Commun. 2010, 393, 404–409. [Google Scholar] [CrossRef]
- Won, J.-S.; Im, Y.-B.; Khan, M.; Contreras, M.; Singh, A.K.; Singh, I. Lovastatin Inhibits Amyloid Precursor Protein (APP) Beta-Cleavage through Reduction of APP Distribution in Lubrol WX Extractable Low Density Lipid Rafts. J. Neurochem. 2008, 105, 1536–1549. [Google Scholar] [CrossRef]
- Miersch, S.; Espey, M.G.; Chaube, R.; Akarca, A.; Tweten, R.; Ananvoranich, S.; Mutus, B. Plasma Membrane Cholesterol Content Affects Nitric Oxide Diffusion Dynamics and Signaling. J. Biol. Chem. 2008, 283, 18513–18521. [Google Scholar] [CrossRef]
- Palacios-Rápalo, S.N.; De Jesús-González, L.A.; Cordero-Rivera, C.D.; Farfan-Morales, C.N.; Osuna-Ramos, J.F.; Martínez-Mier, G.; Quistián-Galván, J.; Muñoz-Pérez, A.; Bernal-Dolores, V.; Del Ángel, R.M.; et al. Cholesterol-Rich Lipid Rafts as Platforms for SARS-CoV-2 Entry. Front. Immunol. 2021, 12, 796855. [Google Scholar] [CrossRef]
- Griffiths, W.J.; Crick, P.J.; Wang, Y. Methods for Oxysterol Analysis: Past, Present and Future. Biochem. Pharmacol. 2013, 86, 3–14. [Google Scholar] [CrossRef]
- Borah, K.; Rickman, O.J.; Voutsina, N.; Ampong, I.; Gao, D.; Baple, E.L.; Dias, I.H.K.; Crosby, A.H.; Griffiths, H.R. A Quantitative LC-MS/MS Method for Analysis of Mitochondrial -Specific Oxysterol Metabolism. Redox Biol. 2020, 36, 101595. [Google Scholar] [CrossRef]
- Murata, M.; Peränen, J.; Schreiner, R.; Wieland, F.; Kurzchalia, T.V.; Simons, K. VIP21/Caveolin Is a Cholesterol-Binding Protein. Proc. Natl. Acad. Sci. USA 1995, 92, 10339–10343. [Google Scholar] [CrossRef]
- Melkonian, K.A.; Ostermeyer, A.G.; Chen, J.Z.; Roth, M.G.; Brown, D.A. Role of Lipid Modifications in Targeting Proteins to Detergent-Resistant Membrane Rafts. Many Raft Proteins Are Acylated, While Few Are Prenylated. J. Biol. Chem. 1999, 274, 3910–3917. [Google Scholar] [CrossRef] [PubMed]
- Yurtsever, D.; Lorent, J.H. Structural Modifications Controlling Membrane Raft Partitioning and Curvature in Human and Viral Proteins. J. Phys. Chem. B 2020, 124, 7574–7585. Available online: https://pubs.acs.org/doi/10.1021/acs.jpcb.0c03435 (accessed on 30 December 2023).
- Heider, S.; Dangerfield, J.A.; Metzner, C. Biomedical Applications of Glycosylphosphatidylinositol-Anchored Proteins. J. Lipid Res. 2016, 57, 1778–1788. [Google Scholar] [CrossRef] [PubMed]
- Ilic, K.; Auer, B.; Mlinac-Jerkovic, K.; Herrera-Molina, R. Neuronal Signaling by Thy-1 in Nanodomains With Specific Ganglioside Composition: Shall We Open the Door to a New Complexity? Front. Cell Dev. Biol. 2019, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Puljko, B.; Stojanović, M.; Ilic, K.; Maček Hrvat, N.; Zovko, A.; Damjanović, V.; Mlinac-Jerkovic, K.; Kalanj-Bognar, S. Redistribution of Gangliosides Accompanies Thermally Induced Na+, K+-ATPase Activity Alternation and Submembrane Localisation in Mouse Brain. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183475. [Google Scholar] [CrossRef] [PubMed]
- Rajamanickam, G.D.; Kastelic, J.P.; Thundathil, J.C. Na/K-ATPase Regulates Bovine Sperm Capacitation through Raft- and Non-Raft-Mediated Signaling Mechanisms. Mol. Reprod. Dev. 2017, 84, 1168–1182. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Tian, J.; Liu, L.; Pierre, S.; Liu, J.; Shapiro, J.; Xie, Z.-J. Identification of a Pool of Non-Pumping Na/K-ATPase. J. Biol. Chem. 2007, 282, 10585–10593. [Google Scholar] [CrossRef] [PubMed]
- Pike, L.J. The Challenge of Lipid Rafts. J. Lipid Res. 2009, 50, S323–S328. [Google Scholar] [CrossRef]
- Fridriksson, E.K.; Shipkova, P.A.; Sheets, E.D.; Holowka, D.; Baird, B.; McLafferty, F.W. Quantitative Analysis of Phospholipids in Functionally Important Membrane Domains from RBL-2H3 Mast Cells Using Tandem High-Resolution Mass Spectrometry. Biochemistry 1999, 38, 8056–8063. [Google Scholar] [CrossRef]
- Prinetti, A.; Chigorno, V.; Tettamanti, G.; Sonnino, S. Sphingolipid-Enriched Membrane Domains from Rat Cerebellar Granule Cells Differentiated in Culture. A Compositional Study. J. Biol. Chem. 2000, 275, 11658–11665. [Google Scholar] [CrossRef]
- D’Aprile, C.; Prioni, S.; Mauri, L.; Prinetti, A.; Grassi, S. Lipid Rafts as Platforms for Sphingosine 1-Phosphate Metabolism and Signalling. Cell. Signal. 2021, 80, 109929. [Google Scholar] [CrossRef] [PubMed]
- Sipione, S.; Monyror, J.; Galleguillos, D.; Steinberg, N.; Kadam, V. Gangliosides in the Brain: Physiology, Pathophysiology and Therapeutic Applications. Front. Neurosci. 2020, 14, 572965. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, N.; Hatsuta, H.; Murayama, S.; Suzuki, A.; Yanagisawa, K. Influence of APOE Genotype and the Presence of Alzheimer’s Pathology on Synaptic Membrane Lipids of Human Brains. J. Neurosci. Res. 2014, 92, 641–650. [Google Scholar] [CrossRef]
- Gupta, N.; DeFranco, A.L. Visualizing Lipid Raft Dynamics and Early Signaling Events during Antigen Receptor-Mediated B-Lymphocyte Activation. Mol. Biol. Cell 2003, 4, 432–444. Available online: https://www.molbiolcell.org/doi/10.1091/mbc.02-05-0078 (accessed on 30 December 2023).
- Mikhalyov, I.; Samsonov, A. Lipid Raft Detecting in Membranes of Live Erythrocytes. Biochim. Biophys. Acta 2011, 1808, 1930–1939. [Google Scholar] [CrossRef] [PubMed]
- Schnaar, R.L. Isolation of Glycosphingolipids. Methods Enzym. 1994, 230, 348–370. [Google Scholar] [CrossRef]
- Heffer-Lauc, M.; Viljetiæ, B.; Vajn, K.; Schnaar, R.L.; Lauc, G. Effects of Detergents on the Redistribution of Gangliosides and GPI-Anchored Proteins in Brain Tissue Sections. J. Histochem. Cytochem. 2007, 55, 805–812. [Google Scholar] [CrossRef]
- Davidsson, P.; Fredman, P.; Månsson, J.-E.; Svennerholm, L. Determination of Gangliosides and Sulfatide in Human Cerebrospinal Fluid with a Microimmunoaffinity Technique. Clin. Chim. Acta 1991, 197, 105–115. [Google Scholar] [CrossRef]
- Ivell, R.; Teerds, K.; Hoffman, G.E. Proper Application of Antibodies for Immunohistochemical Detection: Antibody Crimes and How to Prevent Them. Endocrinology 2014, 155, 676–687. [Google Scholar] [CrossRef]
- Du, Z.; Lin, J.-R.; Rashid, R.; Maliga, Z.; Wang, S.; Aster, J.C.; Izar, B.; Sorger, P.K.; Santagata, S. Qualifying Antibodies for Image-Based Immune Profiling and Multiplexed Tissue Imaging. Nat. Protoc. 2019, 14, 2900–2930. [Google Scholar] [CrossRef]
- Guthmann, M.; Venier, C.; Toledo, D.; Segatori, V.; Alonso, D.; Fainboim, L.; Vázquez, A.M.; Ostrowski, H. Anti-Ganglioside Antibodies Induced in Chickens by an Alum-Adsorbed Anti-Idiotype Antibody Targeting NeuGcGM3. Front. Immunol. 2013, 3, 422. [Google Scholar] [CrossRef]
- Kotani, M.; Ozawa, H.; Kawashima, I.; Ando, S.; Tai, T. Generation of One Set of Monoclonal Antibodies Specific for A-Pathway Ganglio-Series Gangliosides. Biochim. Biophys. Acta 1992, 1117, 97–103. [Google Scholar] [CrossRef]
- Lunn, M.P.; Johnson, L.A.A.; Fromholt, S.E.; Itonori, S.; Huang, J.; Vyas, A.A.; Hildreth, J.E.; Griffin, J.W.; Schnaar, R.L.; Sheikh, K.A. High-Affinity Anti-Ganglioside IgG Antibodies Raised in Complex Ganglioside Knockout Mice. J. Neurochem. 2000, 75, 404–412. [Google Scholar] [CrossRef]
- Bíró, A.; Cervenak, L.; Balogh, A.; Lőrincz, A.; Uray, K.; Horváth, A.; Romics, L.; Matkó, J.; Füst, G.; László, G. Novel Anti-Cholesterol Monoclonal Immunoglobulin G Antibodies as Probes and Potential Modulators of Membrane Raft-Dependent Immune Functions. J. Lipid Res. 2007, 48, 19–29. [Google Scholar] [CrossRef]
- Owen, D.M.; Magenau, A.; Williamson, D.; Gaus, K. The Lipid Raft Hypothesis Revisited—New Insights on Raft Composition and Function from Super-Resolution Fluorescence Microscopy. BioEssays 2012, 34, 739–747. [Google Scholar] [CrossRef]
- Sezgin, E. Super-Resolution Optical Microscopy for Studying Membrane Structure and Dynamics. J. Phys. Condens. Matter 2017, 29, 273001. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Garai, J.A.; Lorizate, M.; Contreras, F.-X. Shedding Light on Membrane Rafts Structure and Dynamics in Living Cells. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2022, 1864, 183813. [Google Scholar] [CrossRef]
- Heilemann, M. Fluorescence Microscopy beyond the Diffraction Limit. J. Biotechnol. 2010, 149, 243–251. [Google Scholar] [CrossRef]
- Renz, M. Fluorescence Microscopy—A Historical and Technical Perspective-Renz-2013-Cytometry Part A—Wiley Online Library. Cytom. Part A 2012, 83, 767–779. Available online: https://onlinelibrary.wiley.com/doi/full/10.1002/cyto.a.22295 (accessed on 30 December 2023).
- Simons, K.; Vaz, W.L.C. Model Systems, Lipid Rafts, and Cell Membranes. Annu. Rev. Biophys. Biomol. Struct. 2004, 33, 269–295. [Google Scholar] [CrossRef]
- Santos, G.; Díaz, M.; Torres, N.V. Lipid Raft Size and Lipid Mobility in Non-Raft Domains Increase during Aging and Are Exacerbated in APP/PS1 Mice Model of Alzheimer’s Disease. Predictions from an Agent-Based Mathematical Model. Front. Physiol. 2016, 7, 90. [Google Scholar] [CrossRef][Green Version]
- Leung, B.O.; Chou, K.C. Review of Super-Resolution Fluorescence Microscopy for Biology. Appl. Spectrosc. 2011, 65, 967–980. [Google Scholar] [CrossRef]
- Valli, J.; Garcia-Burgos, A.; Rooney, L.M.; Vale de Melo e Oliveira, B.; Duncan, R.R.; Rickman, C. Seeing beyond the Limit: A Guide to Choosing the Right Super-Resolution Microscopy Technique. J. Biol. Chem. 2021, 297, 100791. [Google Scholar] [CrossRef]
- Möckl, L.; Moerner, W.E. Super-Resolution Microscopy with Single Molecules in Biology and Beyond–Essentials, Current Trends, and Future Challenges. J. Am. Chem. Soc. 2020, 142, 17828–17844. [Google Scholar] [CrossRef] [PubMed]
- Colson, L.; Kwon, Y.; Nam, S.; Bhandari, A.; Maya, N.M.; Lu, Y.; Cho, Y. Trends in Single-Molecule Total Internal Reflection Fluorescence Imaging and Their Biological Applications with Lab-on-a-Chip Technology. Sensors 2023, 23, 7691. [Google Scholar] [CrossRef]
- Bourgeois, D. Single Molecule Imaging Simulations with Advanced Fluorophore Photophysics. Commun. Biol. 2023, 6, 53. Available online: https://www.nature.com/articles/s42003-023-04432-x (accessed on 30 December 2023). [CrossRef]
- Möbius, W.; Ohno-Iwashita, Y.; van Donselaar, E.G.; Oorschot, V.M.J.; Shimada, Y.; Fujimoto, T.; Heijnen, H.F.G.; Geuze, H.J.; Slot, J.W. Immunoelectron Microscopic Localization of Cholesterol Using Biotinylated and Non-Cytolytic Perfringolysin O. J. Histochem. Cytochem. 2002, 50, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.G.N.; Kusumi, A. Refinement of Singer-Nicolson Fluid-Mosaic Model by Microscopy Imaging: Lipid Rafts and Actin-Induced Membrane Compartmentalization. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2023, 1865, 184093. [Google Scholar] [CrossRef]
- Webster, J.D.; Miller, M.A.; DuSold, D.; Ramos-Vara, J. Effects of Prolonged Formalin Fixation on Diagnostic Immunohistochemistry in Domestic Animals. J. Histochem. Cytochem. 2009, 57, 753–761. [Google Scholar] [CrossRef]
- Shi, S.-R.; Shi, Y.; Taylor, C.R. Antigen Retrieval Immunohistochemistry: Review and Future Prospects in Research and Diagnosis over Two Decades. J. Histochem. Cytochem. 2011, 59, 13–32. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.A.K.; Suzuki, K.G.N.; Shirai, Y.M.; Shibutani, S.T.; Miyahara, M.S.H.; Tsuboi, H.; Yahara, M.; Yoshimura, A.; Mayor, S.; Fujiwara, T.K.; et al. Membrane Molecules Mobile Even after Chemical Fixation. Nat. Methods 2010, 7, 865–866. [Google Scholar] [CrossRef] [PubMed]
- Kamerbeek, C.B.; Borroni, V.; Pediconi, M.F.; Sato, S.B.; Kobayashi, T.; Barrantes, F.J. Antibody-Induced Acetylcholine Receptor Clusters Inhabit Liquid-Ordered and Liquid-Disordered Domains. Biophys. J. 2013, 105, 1601–1611. [Google Scholar] [CrossRef] [PubMed]
- Westphal, D.; Sytnyk, V.; Schachner, M.; Leshchyns’ka, I. Clustering of the Neural Cell Adhesion Molecule (NCAM) at the Neuronal Cell Surface Induces Caspase-8-and-3-Dependent Changes of the Spectrin Meshwork Required for NCAM-Mediated Neurite Outgrowth. J. Biol. Chem. 2010, 285, 42046–42057. [Google Scholar] [CrossRef] [PubMed]
- Wiatr, M.; Staubach, S.; Figueiredo, R.; Stump-Guthier, C.; Ishikawa, H.; Schwerk, C.; Schroten, H.; Hanisch, F.G.; Rudolph, H.; Tenenbaum, T. Echovirus-30 Infection Alters Host Proteins in Lipid Rafts at the Cerebrospinal Fluid Barrier In Vitro—PMC. Microorganisms 2020, 8, 1958. [Google Scholar] [CrossRef]
- Heffer-Lauc, M.; Lauc, G.; Nimrichter, L.; Fromholt, S.E.; Schnaar, R.L. Membrane Redistribution of Gangliosides and Glycosylphosphatidylinositol-Anchored Proteins in Brain Tissue Sections under Conditions of Lipid Raft Isolation. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2005, 1686, 200–208. [Google Scholar] [CrossRef]
- Blank, N.; Schiller, M.; Krienke, S.; Wabnitz, G.; Ho, A.D.; Lorenz, H.-M. Cholera Toxin Binds to Lipid Rafts but Has a Limited Specificity for Ganglioside GM1. Immunol. Cell Biol. 2007, 85, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Sethi, A.; Wands, A.M.; Mettlen, M.; Krishnamurthy, S.; Wu, H.; Kohler, J.J. Cell Type and Receptor Identity Regulate Cholera Toxin Subunit B (CTB) Internalization. Interface Focus 2019, 9, 20180076. [Google Scholar] [CrossRef]
- Ganser, A.L.; Kirschner, D.A.; Willinger, M. Ganglioside Localization on Myelinated Nerve Fibres by Cholera Toxin Binding. J. Neurocytol. 1983, 12, 921–938. [Google Scholar] [CrossRef]
- Vajn, K.; Viljetić, B.; Degmečić, I.V.; Schnaar, R.L.; Heffer, M. Differential Distribution of Major Brain Gangliosides in the Adult Mouse Central Nervous System. PLoS ONE 2013, 8, e75720. [Google Scholar] [CrossRef]
- Kotani, M.; Kawashima, I.; Ozawa, H.; Terashima, T.; Tai, T. Differential Distribution of Major Gangliosides in Rat Central Nervous System Detected by Specific Monoclonal Antibodies. Glycobiology 1993, 3, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Weishaupt, N.; Caughlin, S.; Yeung, K.K.-C.; Whitehead, S.N. Differential Anatomical Expression of Ganglioside GM1 Species Containing D18:1 or D20:1 Sphingosine Detected by MALDI Imaging Mass Spectrometry in Mature Rat Brain. Front. Neuroanat. 2015, 9, 155. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.X.; Whitehead, S.N. Imaging Mass Spectrometry Allows for Neuroanatomic-Specific Detection of Gangliosides in the Healthy and Diseased Brain—Analyst (RSC Publishing). Analyst 2020, 145, 2473–2481. Available online: https://pubs.rsc.org/en/content/articlelanding/2020/an/c9an02270h (accessed on 30 December 2023). [CrossRef]
- Rao, M.; Mayor, S. Use of Forster’s Resonance Energy Transfer Microscopy to Study Lipid Rafts. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2005, 1746, 221–233. [Google Scholar] [CrossRef]
- Kim, H.M.; Jeong, B.H.; Hyon, J.-Y.; An, M.J.; Seo, M.S.; Hong, J.H.; Lee, K.J.; Kim, C.H.; Joo, T.; Hong, S.-C.; et al. Two-Photon Fluorescent Turn-On Probe for Lipid Rafts in Live Cell and Tissue. J. Am. Chem. Soc. 2008, 130, 4246–4247. [Google Scholar] [CrossRef] [PubMed]
- Kenworthy, A.K.; Petranova, N.; Edidin, M. High-Resolution FRET Microscopy of Cholera Toxin B-Subunit and GPI-Anchored Proteins in Cell Plasma Membranes. Mol. Biol. Cell 2000, 11, 1645–1655. [Google Scholar] [CrossRef] [PubMed]
- Collot, M.; Ashokkumar, P.; Anton, H.; Boutant, E.; Faklaris, O.; Galli, T.; Mély, Y.; Danglot, L.; Klymchenko, A.S. MemBright: A Family of Fluorescent Membrane Probes for Advanced Cellular Imaging and Neuroscience—ScienceDirect. Cell Chem. Biol. 2019, 26, 600–614. Available online: https://www.sciencedirect.com/science/article/pii/S2451945619300315 (accessed on 30 December 2023). [CrossRef]
- Kotani, N.; Nakano, T.; Ida, Y.; Ito, R.; Hashizume, M.; Yamaguchi, A.; Seo, M.; Araki, T.; Hojo, Y.; Honke, K.; et al. Analysis of Lipid Raft Molecules in the Living Brain Slices. Neurochem. Int. 2018, 119, 140–150. [Google Scholar] [CrossRef]
- Klymchenko, A.S.; Kreder, R. Fluorescent Probes for Lipid Rafts: From Model Membranes to Living Cells. Chem. Biol. 2014, 21, 97–113. [Google Scholar] [CrossRef]
- Kenworthy, A.K.; Schmieder, S.S.; Raghunathan, K.; Tiwari, A.; Wang, T.; Kelly, C.V.; Lencer, W.I. Cholera Toxin as a Probe for Membrane Biology. Toxins 2021, 13, 543. [Google Scholar] [CrossRef]
- Day, C.A.; Kenworthy, A.K. Functions of Cholera Toxin B-Subunit as a Raft Cross-Linker. Essays Biochem. 2015, 57, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Lai, B.-Q.; Qiu, X.-C.; Zhang, K.; Zhang, R.-Y.; Jin, H.; Li, G.; Shen, H.-Y.; Wu, J.-L.; Ling, E.-A.; Zeng, Y.-S. Cholera Toxin B Subunit Shows Transneuronal Tracing after Injection in an Injured Sciatic Nerve. PLoS ONE 2015, 10, e0144030. [Google Scholar] [CrossRef] [PubMed]
- Fivaz, M.; Abrami, L.; van der Goot, F.G. Pathogens, Toxins, and Lipid Rafts. Protoplasma 2000, 212, 8–14. [Google Scholar] [CrossRef]
- Barrantes, F.J. Chapter Nine—Fluorescence Sensors for Imaging Membrane Lipid Domains and Cholesterol. In Current Topics in Membranes; Model, M.A., Levitan, I., Eds.; New Methods and Sensors for Membrane and Cell Volume Research; Academic Press: Cambridge, MA, USA, 2021; Volume 88, pp. 257–314. [Google Scholar]
- Kim, H.M.; Cho, B.R. Two-Photon Probes for Intracellular Free Metal Ions, Acidic Vesicles, and Lipid Rafts in Live Tissues. Acc. Chem. Res. 2009, 42, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Rohrbough, J.; Rushton, E.; Palanker, L.; Woodruff, E.; Matthies, H.J.G.; Acharya, U.; Acharya, J.K.; Broadie, K. Ceramidase Regulates Synaptic Vesicle Exocytosis and Trafficking. J. Neurosci. 2004, 24, 7789–7803. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.L.; McLeod, C.W.; Bunch, J. Imaging of Phospholipids in Formalin Fixed Rat Brain Sections by Matrix Assisted Laser Desorption/Ionization Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2011, 22, 1991–1998. [Google Scholar] [CrossRef] [PubMed]
- Strnad, Š.; Pražienková, V.; Holubová, M.; Sýkora, D.; Cvačka, J.; Maletínská, L.; Železná, B.; Kuneš, J.; Vrkoslav, V. Mass Spectrometry Imaging of Free-Floating Brain Sections Detects Pathological Lipid Distribution in a Mouse Model of Alzheimer’s-like Pathology. Analyst 2020, 145, 4595–4605. [Google Scholar] [CrossRef]
- Angelini, R.; Yutuc, E.; Wyatt, M.F.; Newton, J.; Yusuf, F.A.; Griffiths, L.; Cooze, B.J.; El Assad, D.; Frache, G.; Rao, W.; et al. Visualizing Cholesterol in the Brain by On-Tissue Derivatization and Quantitative Mass Spectrometry Imaging. Anal. Chem. 2021, 93, 4932–4943. [Google Scholar] [CrossRef]
- Heverin, M.; Bogdanovic, N.; Lütjohann, D.; Bayer, T.; Pikuleva, I.; Bretillon, L.; Diczfalusy, U.; Winblad, B.; Björkhem, I. Changes in the Levels of Cerebral and Extracerebral Sterols in the Brain of Patients with Alzheimer’s Disease. J. Lipid Res. 2004, 45, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Şişecioğlu, M.; Budak, H.; Geffers, L.; Çankaya, M.; Çiftci, M.; Thaller, C.; Eichele, G.; Küfrevioğlu, Ö.İ.; Özdemir, H. A Compendium of Expression Patterns of Cholesterol Biosynthetic Enzymes in the Mouse Embryo. J. Lipid Res. 2015, 56, 1551–1559. [Google Scholar] [CrossRef]
- Korade, Z.; Mi, Z.; Portugal, C.; Schor, N.F. Expression and P75 Neurotrophin Receptor Dependence of Cholesterol Synthetic Enzymes in Adult Mouse Brain. Neurobiol. Aging 2007, 28, 1522–1531. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Cheng, X.; Liu, R.; Li, X.; Hang, L.; Hang, W.; Xu, J.; Yan, X.; Li, J.; Tian, Z. Chemical and Topographical Single-Cell Imaging by Near-Field Desorption Mass Spectrometry. Angew. Chem. Int. Ed. Engl. 2019, 58, 4541–4546. [Google Scholar] [CrossRef]
- Niehaus, M.; Soltwisch, J.; Belov, M.E.; Dreisewerd, K. Transmission-Mode MALDI-2 Mass Spectrometry Imaging of Cells and Tissues at Subcellular Resolution. Nat. Methods 2019, 16, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.S.; Vickerman, J.C. Secondary Ion Mass Spectrometry: Characterizing Complex Samples in Two and Three Dimensions. Anal. Chem. 2013, 85, 610–639. [Google Scholar] [CrossRef]
- Winograd, N. Gas Cluster Ion Beams for Secondary Ion Mass Spectrometry. Annu. Rev. Anal. Chem. 2018, 11, 29–48. [Google Scholar] [CrossRef]
- Tian, H.; Sheraz née Rabbani, S.; Vickerman, J.C.; Winograd, N. Multiomics Imaging Using High-Energy Water Gas Cluster Ion Beam Secondary Ion Mass Spectrometry [(H2O)n-GCIB-SIMS] of Frozen-Hydrated Cells and Tissue. Anal. Chem. 2021, 93, 7808–7814. [Google Scholar] [CrossRef]
- Lim, H.; Lee, S.Y.; Park, Y.; Jin, H.; Seo, D.; Jang, Y.H.; Moon, D.W. Mass Spectrometry Imaging of Untreated Wet Cell Membranes in Solution Using Single-Layer Graphene. Nat. Methods 2021, 18, 316–320. [Google Scholar] [CrossRef]
- Yoon, J.H.; Seo, Y.; Jo, Y.S.; Lee, S.; Cho, E.; Cazenave-Gassiot, A.; Shin, Y.-S.; Moon, M.H.; An, H.J.; Wenk, M.R.; et al. Brain Lipidomics: From Functional Landscape to Clinical Significance. Sci. Adv. 2022, 8, eadc9317. [Google Scholar] [CrossRef]
- Samhan-Arias, A.K.; Poejo, J.; Marques-da-Silva, D.; Martínez-Costa, O.H.; Gutierrez-Merino, C. Are There Lipid Membrane-Domain Subtypes in Neurons with Different Roles in Calcium Signaling? Molecules 2023, 28, 7909. [Google Scholar] [CrossRef]


| Material | Homogenization Method | Solubilization Method | Density Gradient | Analyzed Membrane Constituents | Reference | ||
|---|---|---|---|---|---|---|---|
| Chol | Proteins | Lipids | |||||
| mouse brain | 23-gauge needle | no detergent | sucrose and OptiprepTM | + | Flot1, TfR, MBP, GRASP65 | − | [66] |
| mouse brain; HEK 293 cells | not specified; sonication | no detergent and Triton X-100 | sucrose | + | caveolin-1, Flot1, TfR, GABAA receptor subunits, NMDA receptor subunit NR1A | − | [67] |
| MDCK cells | Dounce homogenizer | Triton X-100 | sucrose | + | PLAP, GPI-anchored proteins | PLs, SM, cardiolipin, LacCer, cerebrosides, sulfatides, gangliosides | [68] original method reporting the use of Triton X-100 |
| mouse brain and cerebellar granule cells | Dounce homogenizer | Triton X-100 | sucrose | + | Fyn, Lyn, PrP, Akt | PLs, SM, gangliosides | [69] |
| 3A9 T cell hybridoma; mouse thymocytes; mouse T cells | sonication | Brij O20 | sucrose | + | Thy-1, Lck, Rab-5 | PLs, SM, gangliosides | [70] original method reporting the use of Brij O20 |
| mouse brain | Potter–Elvehjem glass homogenizer with a Teflon pestle | Brij O20 and Triton X-100 | sucrose | − | Flot1, TfR, GluA2, APP, Np65 | gangliosides | [64] |
| CHO cells | 23-gauge needle | Lubrol WX, Triton X-100 and CHAPSO | sucrose | + | Flot1, TfR, APP | − | [71] |
| primary rat hippocampal neurons | Dounce homogenizer | Lubrol WX and Triton X-100 | Nycodenz® | + | CD71, clathrin, Flot1, PrP, APP | − | [72] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mlinac-Jerkovic, K.; Kalanj-Bognar, S.; Heffer, M.; Blažetić, S. Methodological Pitfalls of Investigating Lipid Rafts in the Brain: What Are We Still Missing? Biomolecules 2024, 14, 156. https://doi.org/10.3390/biom14020156
Mlinac-Jerkovic K, Kalanj-Bognar S, Heffer M, Blažetić S. Methodological Pitfalls of Investigating Lipid Rafts in the Brain: What Are We Still Missing? Biomolecules. 2024; 14(2):156. https://doi.org/10.3390/biom14020156
Chicago/Turabian StyleMlinac-Jerkovic, Kristina, Svjetlana Kalanj-Bognar, Marija Heffer, and Senka Blažetić. 2024. "Methodological Pitfalls of Investigating Lipid Rafts in the Brain: What Are We Still Missing?" Biomolecules 14, no. 2: 156. https://doi.org/10.3390/biom14020156
APA StyleMlinac-Jerkovic, K., Kalanj-Bognar, S., Heffer, M., & Blažetić, S. (2024). Methodological Pitfalls of Investigating Lipid Rafts in the Brain: What Are We Still Missing? Biomolecules, 14(2), 156. https://doi.org/10.3390/biom14020156