
Arabidopsis RALF4 Rapidly Halts Pollen Tube Growth by Increasing ROS and Decreasing Calcium Cytoplasmic Tip Levels

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Stable Fusion Protein Expression in Arabidopsis Pollen Tubes
2.3. Synthetic Peptides
2.4. In Vitro Pollen Germination
2.5. Measurement of Root Growth Inhibition by Synthetic RALFs
2.6. Ratiometric Imaging
2.6.1. HyPer1 Imaging
2.6.2. YC3.60 Imaging
3. Results
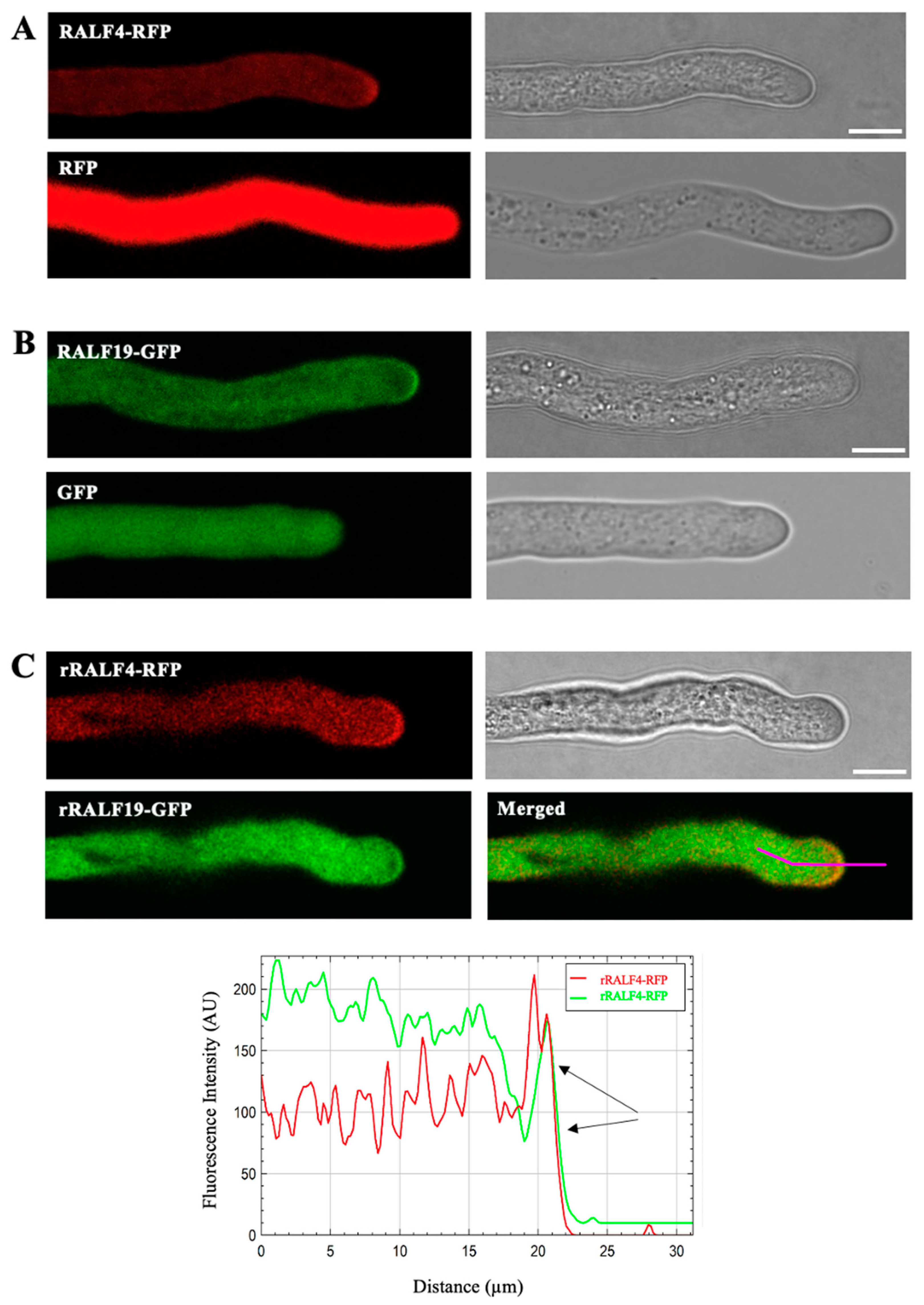
3.1. RALF4 and RALF19 Peptides Colocalize in the Tip Region of the Pollen Tube
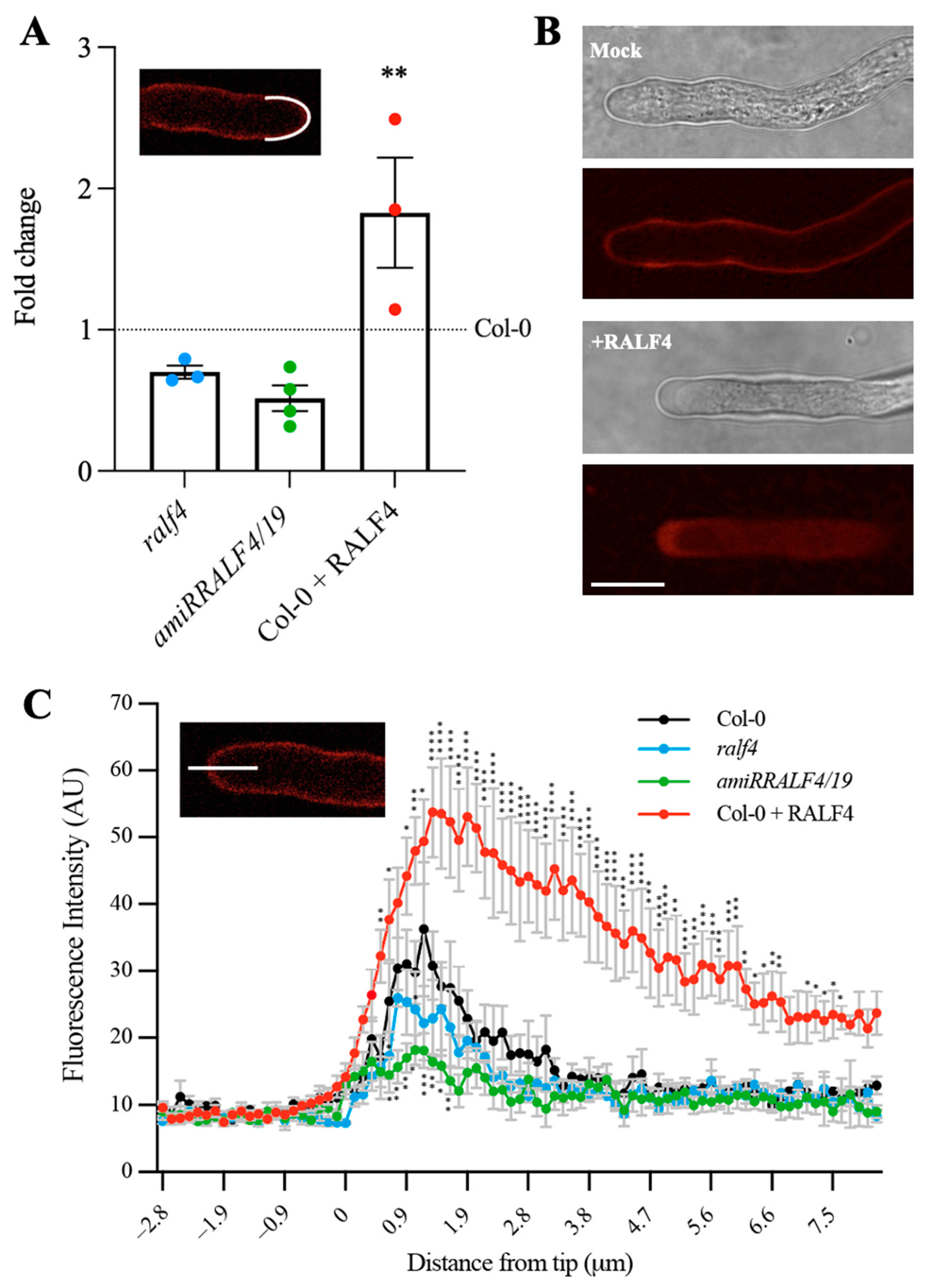
3.2. Synthetic RALF4 Peptide Inhibits Pollen Tube Growth
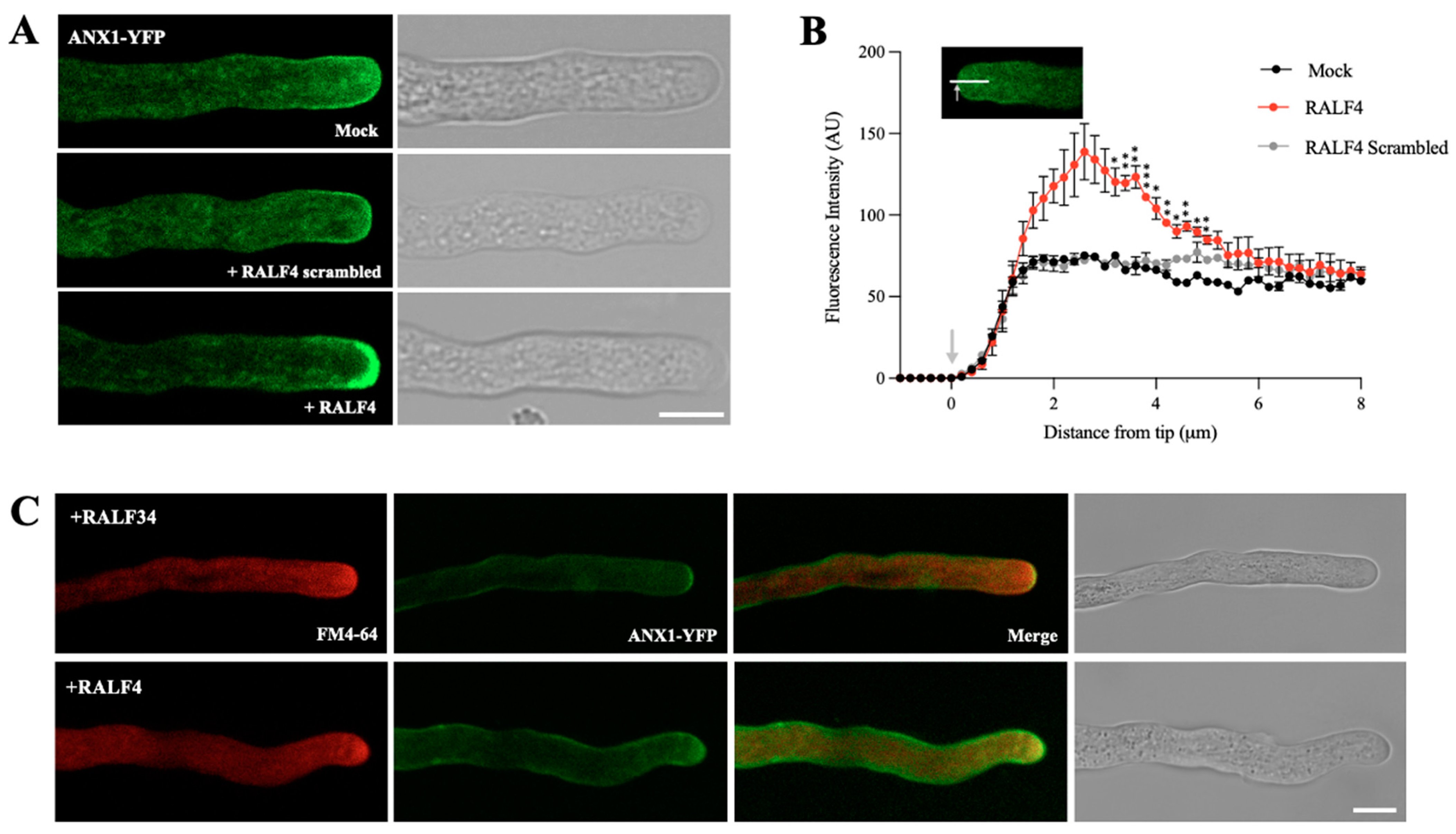
3.3. Synthetic RALF4 Triggers Cell Wall Accumulation and Plasma Membrane Retraction at the Tip of Pollen Tubes
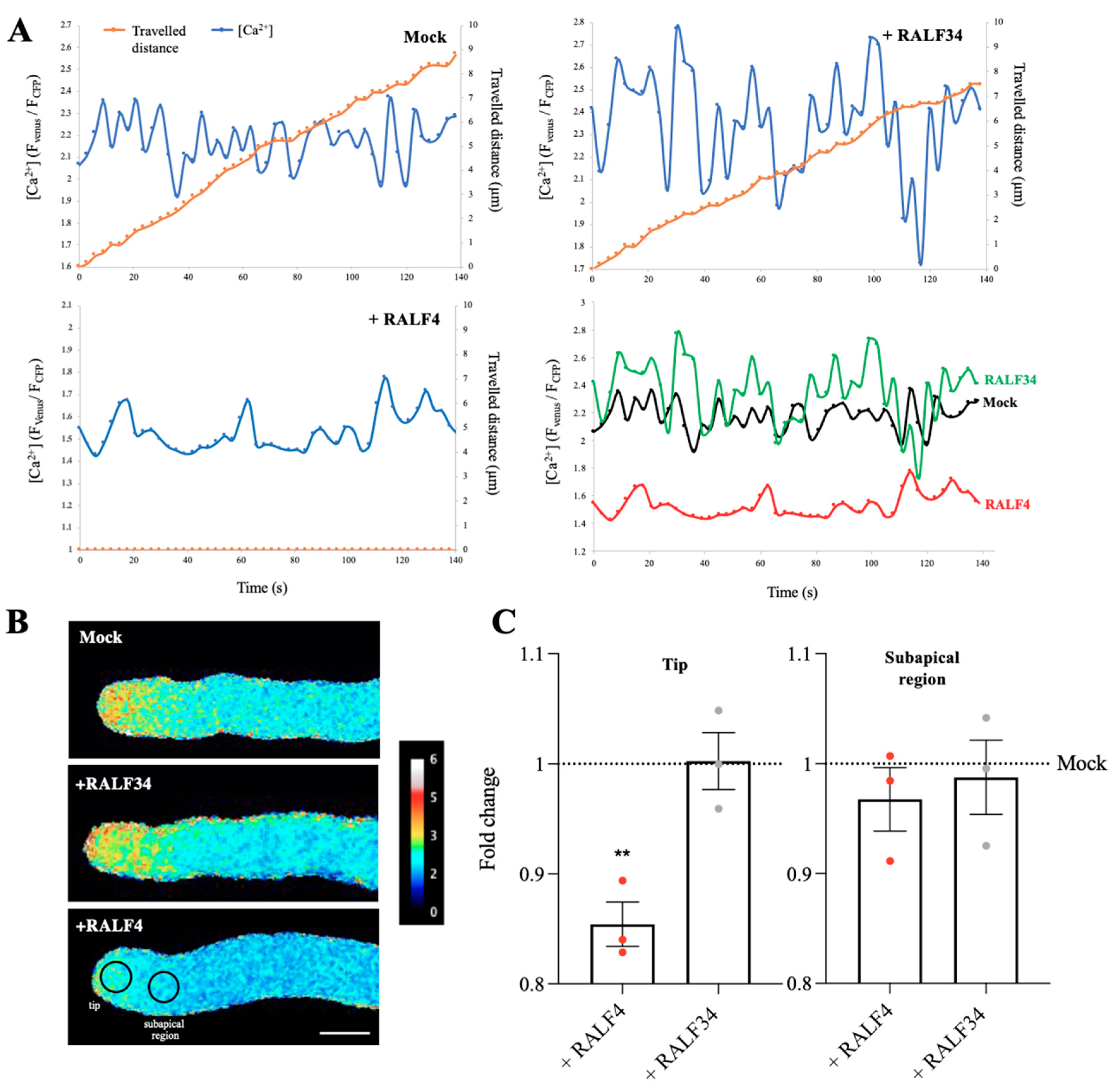
3.4. Synthetic RALF4 Increases Cytoplasmic ROS Levels and Decreases Cytoplasmic Calcium Levels in Pollen Tubes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, M.A.; Harper, J.F.; Palanivelu, R. A Fruitful Journey: Pollen Tube Navigation from Germination to Fertilization. Annu. Rev. Plant Biol. 2019, 70, 809–837. [Google Scholar] [CrossRef] [PubMed]
- Pearce, G.; Moura, D.S.; Stratmann, J.; Ryan, C.A. RALF, a 5-kDa Ubiquitous Polypeptide in Plants, Arrests Root Growth and Development. Proc. Natl. Acad. Sci. USA 2001, 98, 12843–12847. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.; Smet, I.D. Understanding the RALF Family: A Tale of Many Species. Trends Plant Sci. 2014, 19, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, M.R.; Haruta, M.; Moura, D.S. Twenty Years of Progress in Physiological and Biochemical Investigation of RALF Peptides. Plant Physiol. 2020, 182, 1657–1666. [Google Scholar] [CrossRef] [PubMed]
- Somoza, S.C.; Sede, A.R.; Boccardo, N.A.; Muschietti, J.P. Keeping up with the RALFs: How These Small Peptides Control Pollen–Pistil Interactions in Arabidopsis. New Phytol. 2021, 229, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.; Zhao, Y.; Liu, M.-C.; Zhou, L.-Z.; Wang, L.; Zhong, S.; Hou, S.; Jiang, J.; Liu, T.; Huang, Q.; et al. LLG2/3 Are Co-Receptors in BUPS/ANX-RALF Signaling to Regulate Arabidopsis Pollen Tube Integrity. Curr. Biol. 2019, 29, 3256–3265.e5. [Google Scholar] [CrossRef]
- Zhu, L.; Chu, L.-C.; Liang, Y.; Zhang, X.-Q.; Chen, L.-Q.; Ye, D. The Arabidopsis CrRLK1L Protein Kinases BUPS1 and BUPS2 Are Required for Normal Growth of Pollen Tubes in the Pistil. Plant J. Cell Mol. Biol. 2018, 95, 474–486. [Google Scholar] [CrossRef]
- Feng, H.; Liu, C.; Fu, R.; Zhang, M.; Li, H.; Shen, L.; Wei, Q.; Sun, X.; Xu, L.; Ni, B.; et al. LORELEI-LIKE GPI-ANCHORED PROTEINS 2/3 Regulate Pollen Tube Growth as Chaperones and Coreceptors for ANXUR/BUPS Receptor Kinases in Arabidopsis. Mol. Plant 2019, 12, 1612–1623. [Google Scholar] [CrossRef]
- Li, H.; Yang, Y.; Zhang, H.; Li, C.; Du, P.; Bi, M.; Chen, T.; Qian, D.; Niu, Y.; Ren, H.; et al. The Arabidopsis GPI-Anchored Protein COBL11 Is Necessary for Regulating Pollen Tube Integrity. Cell Rep. 2023, 42, 113353. [Google Scholar] [CrossRef]
- Boisson-Dernier, A.; Franck, C.M.; Lituiev, D.S.; Grossniklaus, U. Receptor-like Cytoplasmic Kinase MARIS Functions Downstream of CrRLK1L-Dependent Signaling during Tip Growth. Proc. Natl. Acad. Sci. USA 2015, 112, 12211–12216. [Google Scholar] [CrossRef]
- Boisson-Dernier, A.; Lituiev, D.S.; Nestorova, A.; Franck, C.M.; Thirugnanarajah, S.; Grossniklaus, U. ANXUR Receptor-like Kinases Coordinate Cell Wall Integrity with Growth at the Pollen Tube Tip via NADPH Oxidases. PLoS Biol. 2013, 11, e1001719. [Google Scholar] [CrossRef] [PubMed]
- Lassig, R.; Gutermuth, T.; Bey, T.D.; Konrad, K.R.; Romeis, T. Pollen Tube NAD(P)H Oxidases Act as a Speed Control to Dampen Growth Rate Oscillations during Polarized Cell Growth. Plant J. Cell Mol. Biol. 2014, 78, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Sede, A.R.; Borassi, C.; Wengier, D.L.; Mecchia, M.A.; Estevez, J.M.; Muschietti, J.P. Arabidopsis Pollen Extensins LRX Are Required for Cell Wall Integrity during Pollen Tube Growth. FEBS Lett. 2018, 592, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Moussu, S.; Lee, H.K.; Haas, K.T.; Broyart, C.; Rathgeb, U.; De Bellis, D.; Levasseur, T.; Schoenaers, S.; Fernandez, G.S.; Grossniklaus, U.; et al. Plant Cell Wall Patterning and Expansion Mediated by Protein-Peptide-Polysaccharide Interaction. Science 2023, 382, 719–725. [Google Scholar] [CrossRef]
- Mecchia, M.A.; Santos-Fernandez, G.; Duss, N.N.; Somoza, S.C.; Boisson-Dernier, A.; Gagliardini, V.; Martínez-Bernardini, A.; Fabrice, T.N.; Ringli, C.; Muschietti, J.P.; et al. RALF4/19 Peptides Interact with LRX Proteins to Control Pollen Tube Growth in Arabidopsis. Science 2017, 358, 1600–1603. [Google Scholar] [CrossRef]
- Fabrice, T.N.; Vogler, H.; Draeger, C.; Munglani, G.; Gupta, S.; Herger, A.G.; Knox, P.; Grossniklaus, U.; Ringli, C. LRX Proteins Play a Crucial Role in Pollen Grain and Pollen Tube Cell Wall Development. Plant Physiol. 2018, 176, 1981–1992. [Google Scholar] [CrossRef]
- Wang, X.; Wang, K.; Yin, G.; Liu, X.; Liu, M.; Cao, N.; Duan, Y.; Gao, H.; Wang, W.; Ge, W.; et al. Pollen-Expressed Leucine-Rich Repeat Extensins Are Essential for Pollen Germination and Growth. Plant Physiol. 2018, 176, 1993–2006. [Google Scholar] [CrossRef]
- Ge, Z.; Bergonci, T.; Zhao, Y.; Zou, Y.; Du, S.; Liu, M.-C.; Luo, X.; Ruan, H.; García-Valencia, L.E.; Zhong, S.; et al. Arabidopsis Pollen Tube Integrity and Sperm Release Are Regulated by RALF-Mediated Signaling. Science 2017, 358, 1596–1600. [Google Scholar] [CrossRef]
- Rounds, C.M.; Lubeck, E.; Hepler, P.K.; Winship, L.J. Propidium Iodide Competes with Ca2+ to Label Pectin in Pollen Tubes and Arabidopsis Root Hairs. Plant Physiol. 2011, 157, 175–187. [Google Scholar] [CrossRef]
- Nakagawa, T.; Kurose, T.; Hino, T.; Tanaka, K.; Kawamukai, M.; Niwa, Y.; Toyooka, K.; Matsuoka, K.; Jinbo, T.; Kimura, T. Development of Series of Gateway Binary Vectors, pGWBs, for Realizing Efficient Construction of Fusion Genes for Plant Transformation. J. Biosci. Bioeng. 2007, 104, 34–41. [Google Scholar] [CrossRef]
- Boavida, L.C.; McCormick, S. Temperature as a Determinant Factor for Increased and Reproducible in Vitro Pollen Germination in Arabidopsis Thaliana. Plant J. Cell Mol. Biol. 2007, 52, 570–582. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Barrera, A.; Velarde-Buendía, A.; Zepeda, I.; Sanchez, F.; Quinto, C.; Sánchez-Lopez, R.; Cheung, A.Y.; Wu, H.-M.; Cardenas, L. Hyper, a Hydrogen Peroxide Sensor, Indicates the Sensitivity of the Arabidopsis Root Elongation Zone to Aluminum Treatment. Sensors 2015, 15, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Nagai, T.; Yamada, S.; Tominaga, T.; Ichikawa, M.; Miyawaki, A. Expanded Dynamic Range of Fluorescent Indicators for Ca(2+) by Circularly Permuted Yellow Fluorescent Proteins. Proc. Natl. Acad. Sci. USA 2004, 101, 10554–10559. [Google Scholar] [CrossRef] [PubMed]
- Morato do Canto, A.; Ceciliato, P.H.O.; Ribeiro, B.; Ortiz Morea, F.A.; Franco Garcia, A.A.; Silva-Filho, M.C.; Moura, D.S. Biological Activity of Nine Recombinant AtRALF Peptides: Implications for Their Perception and Function in Arabidopsis. Plant Physiol. Biochem. 2014, 75, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Abarca, A.; Franck, C.M.; Zipfel, C. Family-Wide Evaluation of RAPID ALKALINIZATION FACTOR Peptides. Plant Physiol. 2021, 187, 996–1010. [Google Scholar] [CrossRef]
- Zhou, X.; Lu, J.; Zhang, Y.; Guo, J.; Lin, W.; Van Norman, J.M.; Qin, Y.; Zhu, X.; Yang, Z. Membrane Receptor-Mediated Mechano-Transduction Maintains Cell Integrity during Pollen Tube Growth within the Pistil. Dev. Cell 2021, 56, 1030–1042.e6. [Google Scholar] [CrossRef]
- Gonneau, M.; Desprez, T.; Martin, M.; Doblas, V.G.; Bacete, L.; Miart, F.; Sormani, R.; Hématy, K.; Renou, J.; Landrein, B.; et al. Receptor Kinase THESEUS1 Is a Rapid Alkalinization Factor 34 Receptor in Arabidopsis. Curr. Biol. 2018, 28, 2452–2458.e4. [Google Scholar] [CrossRef]
- Rößling, A.-K.; Dünser, K.; Liu, C.; Lauw, S.; Rodriguez-Franco, M.; Kalmbach, L.; Barbez, E.; Kleine-Vehn, J. Pectin Methylesterase Activity Is Required for RALF1 Peptide Signalling Output. eLife 2024, 13, RP96943. [Google Scholar] [CrossRef]
- Liu, M.-C.J.; Yeh, F.-L.J.; Yvon, R.; Simpson, K.; Jordan, S.; Chambers, J.; Wu, H.-M.; Cheung, A.Y. Extracellular Pectin-RALF Phase Separation Mediates FERONIA Global Signaling Function. Cell 2024, 187, 312–330.e22. [Google Scholar] [CrossRef]
- Gao, Q.; Wang, C.; Xi, Y.; Shao, Q.; Hou, C.; Li, L.; Luan, S. RALF Signaling Pathway Activates MLO Calcium Channels to Maintain Pollen Tube Integrity. Cell Res. 2023, 33, 71–79. [Google Scholar] [CrossRef]
- Barberini, M.L.; Sigaut, L.; Huang, W.; Mangano, S.; Juarez, S.P.D.; Marzol, E.; Estevez, J.; Obertello, M.; Pietrasanta, L.; Tang, W.; et al. Calcium Dynamics in Tomato Pollen Tubes Using the Yellow Cameleon 3.6 Sensor. Plant Reprod. 2018, 31, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.-G.; Swanson, S.J.; Gilroy, S. High-Resolution Imaging of Ca2+, Redox Status, ROS and pH Using GFP Biosensors. Plant J. 2012, 70, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.E.; Postiglione, A.E.; Muday, G.K. Reactive Oxygen Species Function as Signaling Molecules in Controlling Plant Development and Hormonal Responses. Curr. Opin. Plant Biol. 2022, 69, 102293. [Google Scholar] [CrossRef] [PubMed]
- Michard, E.; Dias, P.; Feijó, J. Tobacco Pollen Tubes as Cellular Models for Ion Dynamics: Improved Spatial and Temporal Resolution of Extracellular Flux and Free Cytosolic Concentration of Calcium and Protons Using pHluorin and YC3.1 CaMeleon. Sex. Plant Reprod. 2008, 21, 169–181. [Google Scholar] [CrossRef]
- Grenzi, M.; Resentini, F.; Vanneste, S.; Zottini, M.; Bassi, A.; Costa, A. Illuminating the Hidden World of Calcium Ions in Plants with a Universe of Indicators. Plant Physiol. 2021, 187, 550–571. [Google Scholar] [CrossRef]
- Gao, Q.; Wang, C.; Xi, Y.; Shao, Q.; Li, L.; Luan, S. A Receptor-Channel Trio Conducts Ca2+ Signalling for Pollen Tube Reception. Nature 2022, 607, 534–539. [Google Scholar] [CrossRef]
- Zhong, S.; Li, L.; Wang, Z.; Zengxiang, G.; Li, Q.; Bleckmann, A.; Wang, J.; Song, Z.; Shi, Y.; Liu, T.; et al. RALF Peptide Signaling Controls the Polytubey Block in Arabidopsis. Science 2022, 375, 290–296. [Google Scholar] [CrossRef]
- Loraine, A.E.; McCormick, S.; Estrada, A.; Patel, K.; Qin, P. RNA-Seq of Arabidopsis Pollen Uncovers Novel Transcription and Alternative Splicing. Plant Physiol. 2013, 162, 1092–1109. [Google Scholar] [CrossRef]
- Meng, J.-G.; Liang, L.; Jia, P.-F.; Wang, Y.-C.; Li, H.-J.; Yang, W.-C. Integration of Ovular Signals and Exocytosis of a Ca2+ Channel by MLOs in Pollen Tube Guidance. Nat. Plants 2020, 6, 143–153. [Google Scholar] [CrossRef]
- Pei, S.; Tao, Q.; Li, W.; Qi, G.; Wang, B.; Wang, Y.; Dai, S.; Shen, Q.; Wang, X.; Wu, X.; et al. Osmosensor-Mediated Control of Ca2+ Spiking in Pollen Germination. Nature 2024, 629, 1118–1125. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Somoza, S.C.; Boccardo, N.A.; Santin, F.; Sede, A.R.; Wengier, D.L.; Boisson-Dernier, A.; Muschietti, J.P. Arabidopsis RALF4 Rapidly Halts Pollen Tube Growth by Increasing ROS and Decreasing Calcium Cytoplasmic Tip Levels. Biomolecules 2024, 14, 1375. https://doi.org/10.3390/biom14111375
Somoza SC, Boccardo NA, Santin F, Sede AR, Wengier DL, Boisson-Dernier A, Muschietti JP. Arabidopsis RALF4 Rapidly Halts Pollen Tube Growth by Increasing ROS and Decreasing Calcium Cytoplasmic Tip Levels. Biomolecules. 2024; 14(11):1375. https://doi.org/10.3390/biom14111375
Chicago/Turabian StyleSomoza, Sofía C., Noelia A. Boccardo, Franco Santin, Ana R. Sede, Diego L. Wengier, Aurélien Boisson-Dernier, and Jorge P. Muschietti. 2024. "Arabidopsis RALF4 Rapidly Halts Pollen Tube Growth by Increasing ROS and Decreasing Calcium Cytoplasmic Tip Levels" Biomolecules 14, no. 11: 1375. https://doi.org/10.3390/biom14111375
APA StyleSomoza, S. C., Boccardo, N. A., Santin, F., Sede, A. R., Wengier, D. L., Boisson-Dernier, A., & Muschietti, J. P. (2024). Arabidopsis RALF4 Rapidly Halts Pollen Tube Growth by Increasing ROS and Decreasing Calcium Cytoplasmic Tip Levels. Biomolecules, 14(11), 1375. https://doi.org/10.3390/biom14111375